
The Unusual Lipid A Structure and Immunoinhibitory Activity of LPS from Marine Bacteria Echinicola pacifica KMM 6172T and Echinicola vietnamensis KMM 6221T

 , ,
, ,  , ,
, ,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria Isolation and Growth
2.2. Isolation and Purification of the LPS from E. pacifica KMM 6172T and E. vietnamensis KMM 6221T
2.3. Fatty Acid Compositional Analysis of LPS from E. pacifica KMM 6172T and E. vietnamensis KMM 6221T
2.4. Isolation and Analysis of the Lipid A Fractions
2.5. MALDI-TOF Mass Spectrometry Analysis
2.6. HEK Cell Cultures
2.7. HEK-BlueTM Null2, hTLR2 and hTLR4 Stimulation and Competition Assays
3. Results
3.1. Isolation and Fatty Acid Compositional Analysis of the LPS from E. pacifica KMM 6172T and E. vietnamensis KMM 6221T
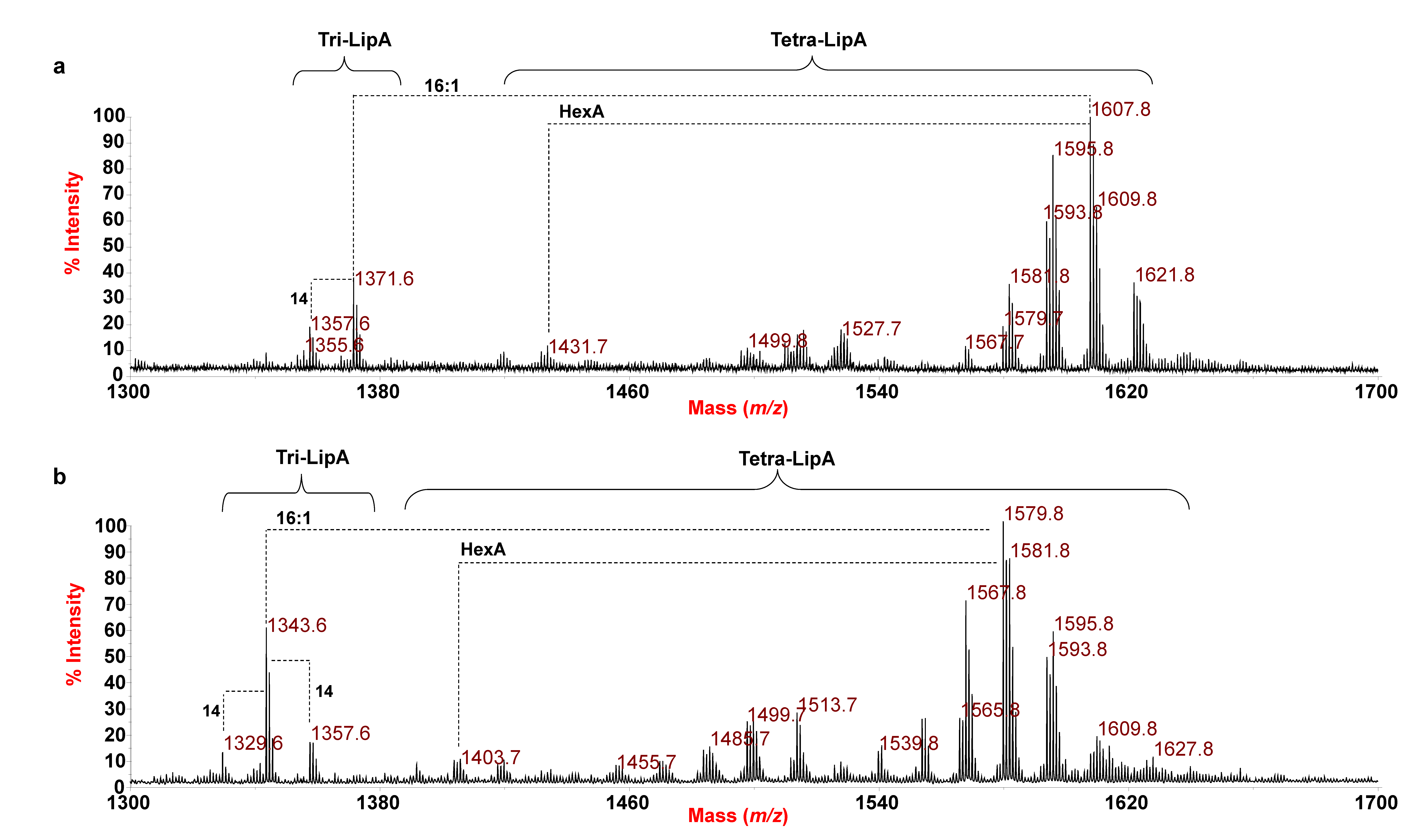
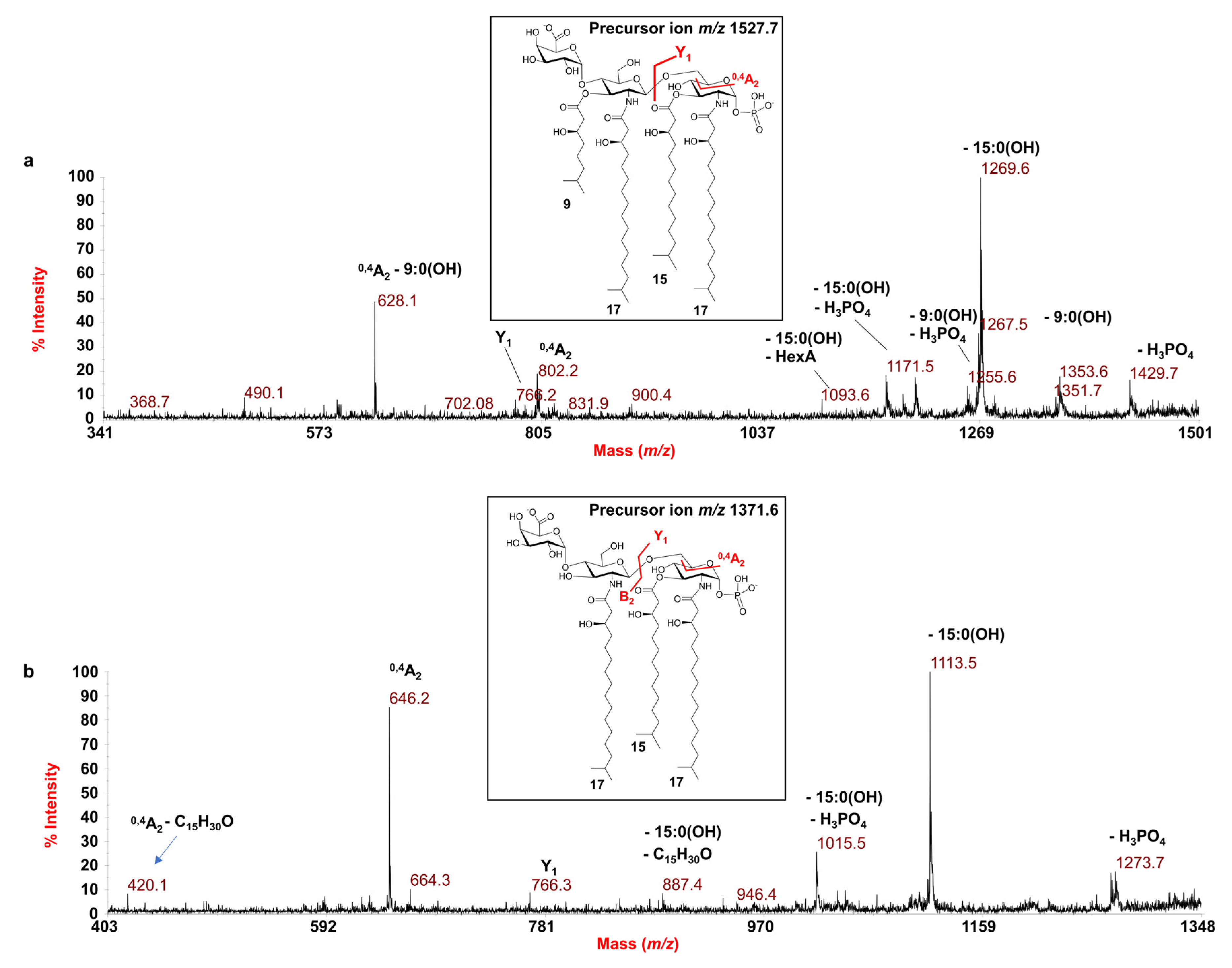
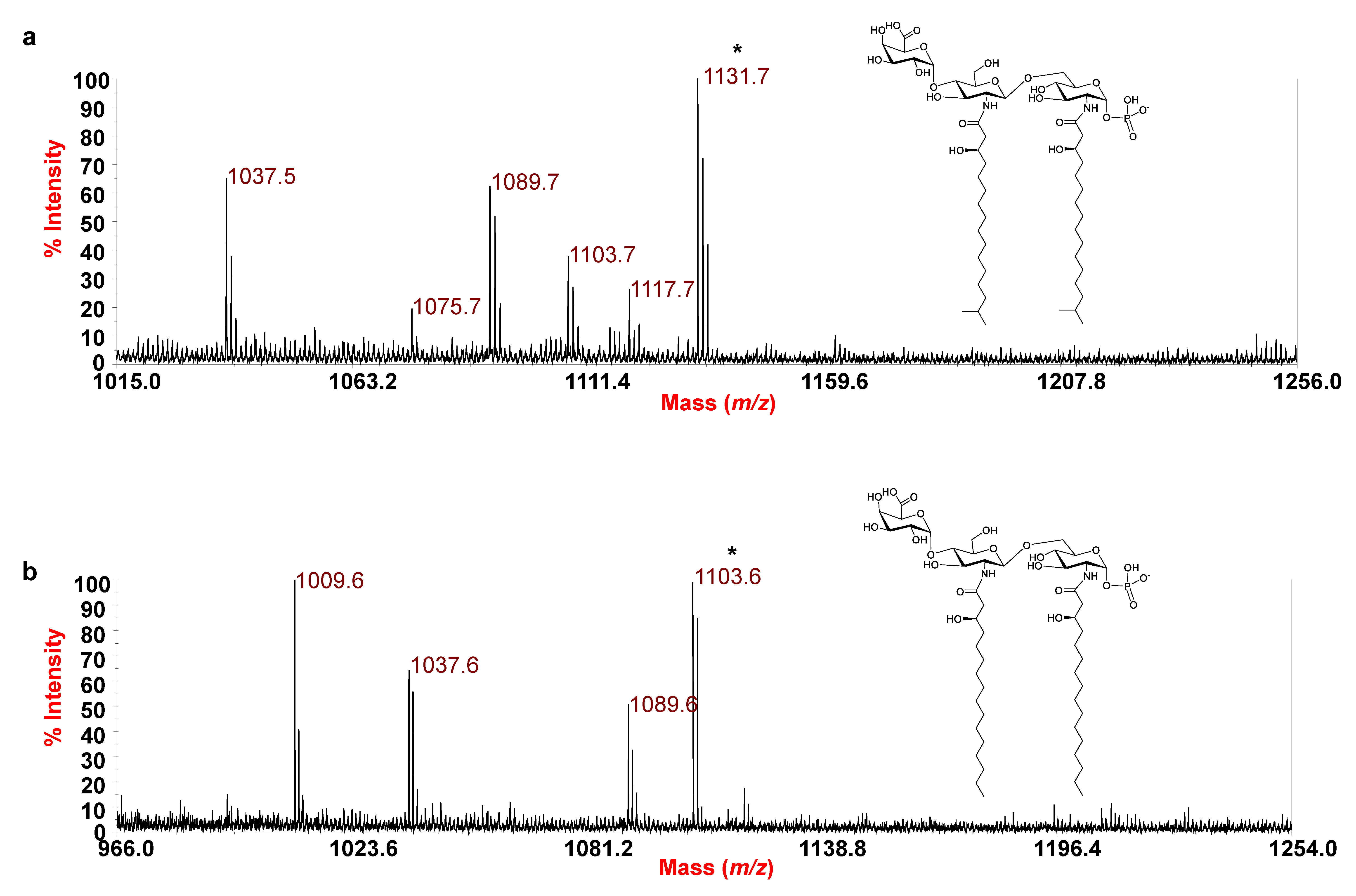
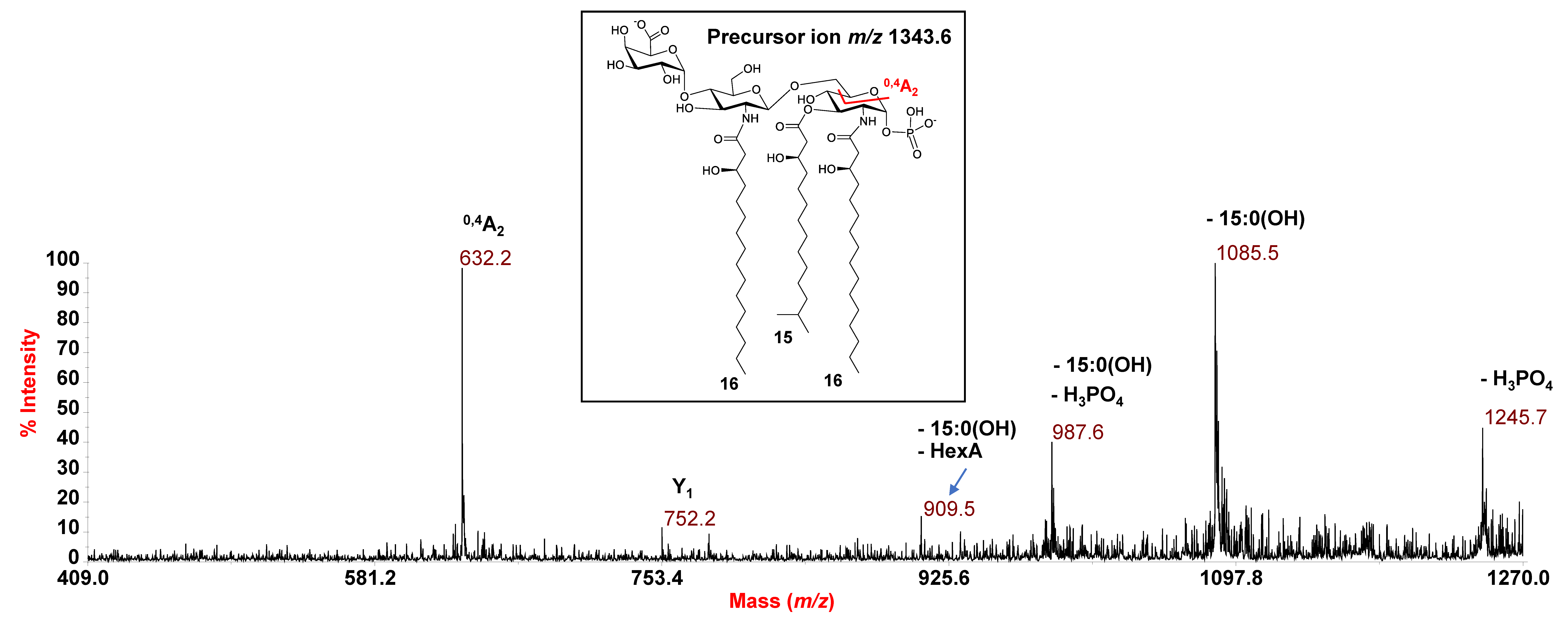
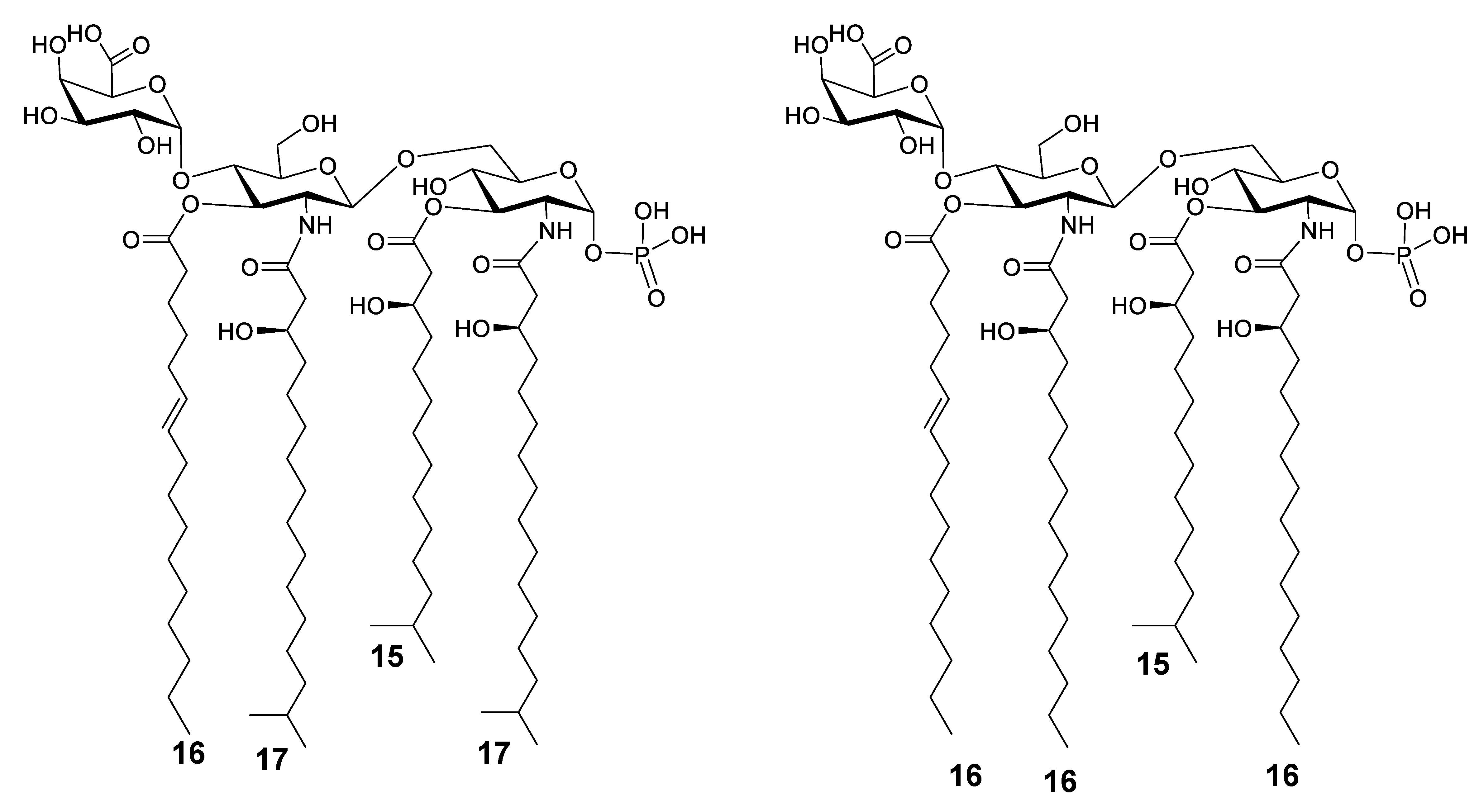
3.2. MALDI-TOF MS and MS/MS Analysis on the Isolated Lipid A from E. pacifica KMM 6172T
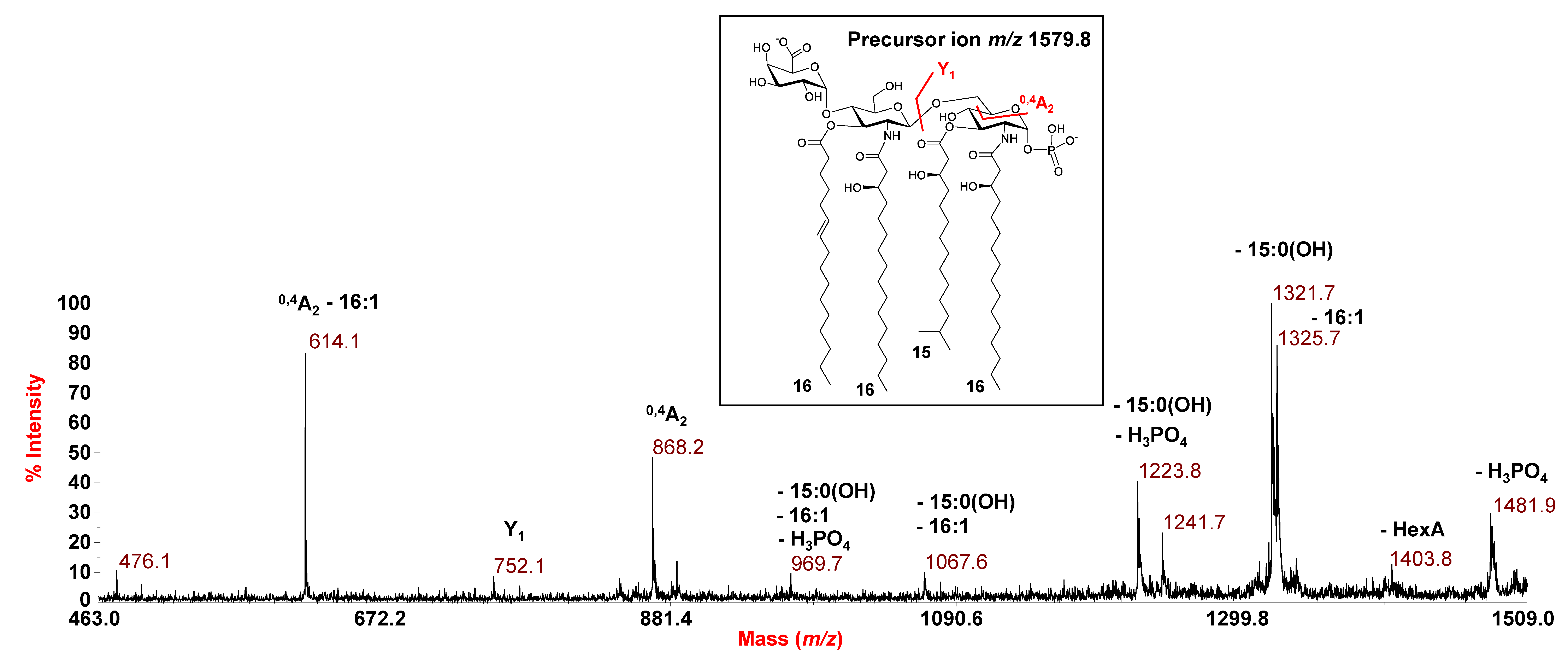
3.3. MALDI MS and MS/MS Analysis on the Isolated Lipid A from E. vietnamensis KMM 6221T
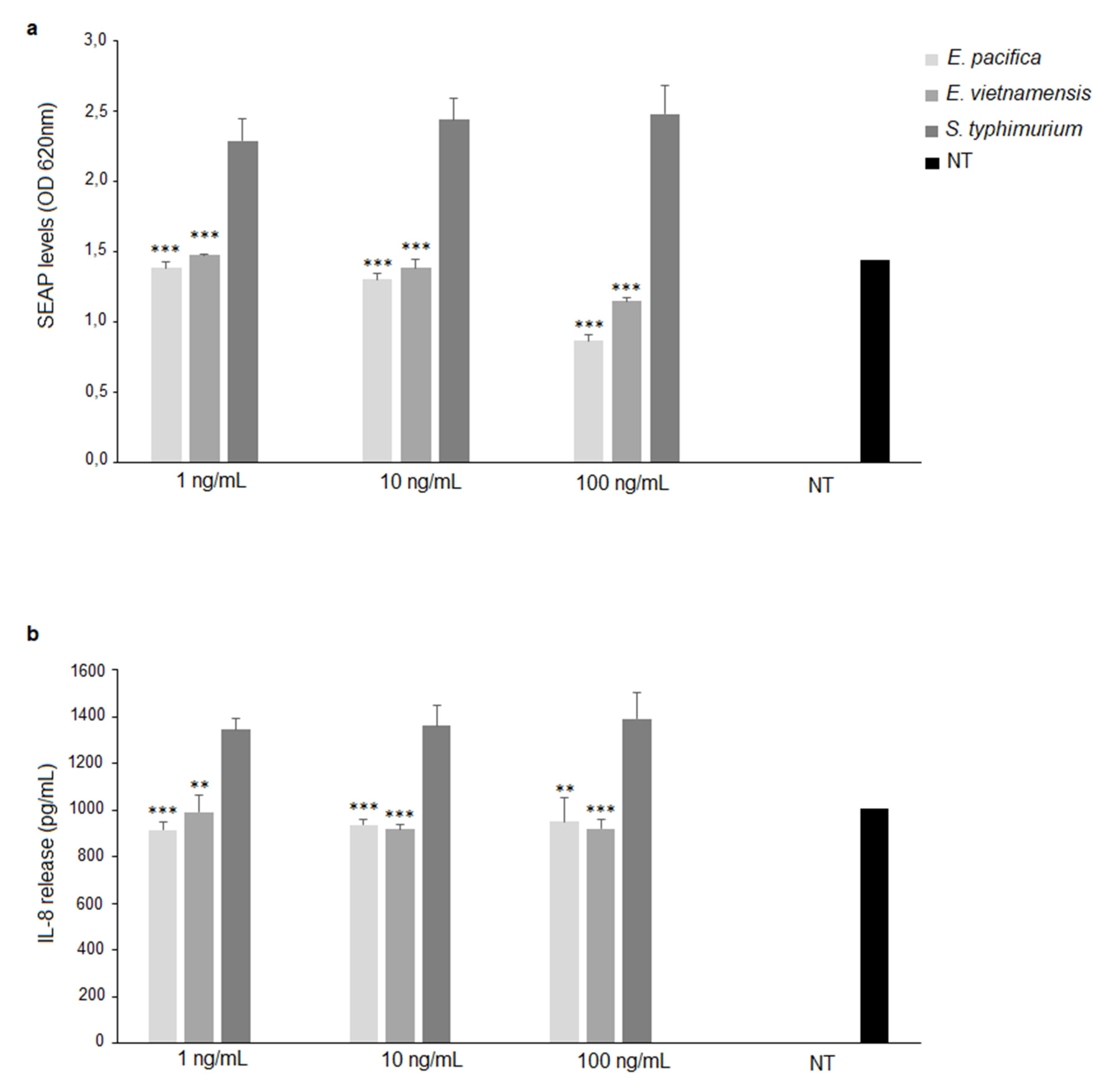
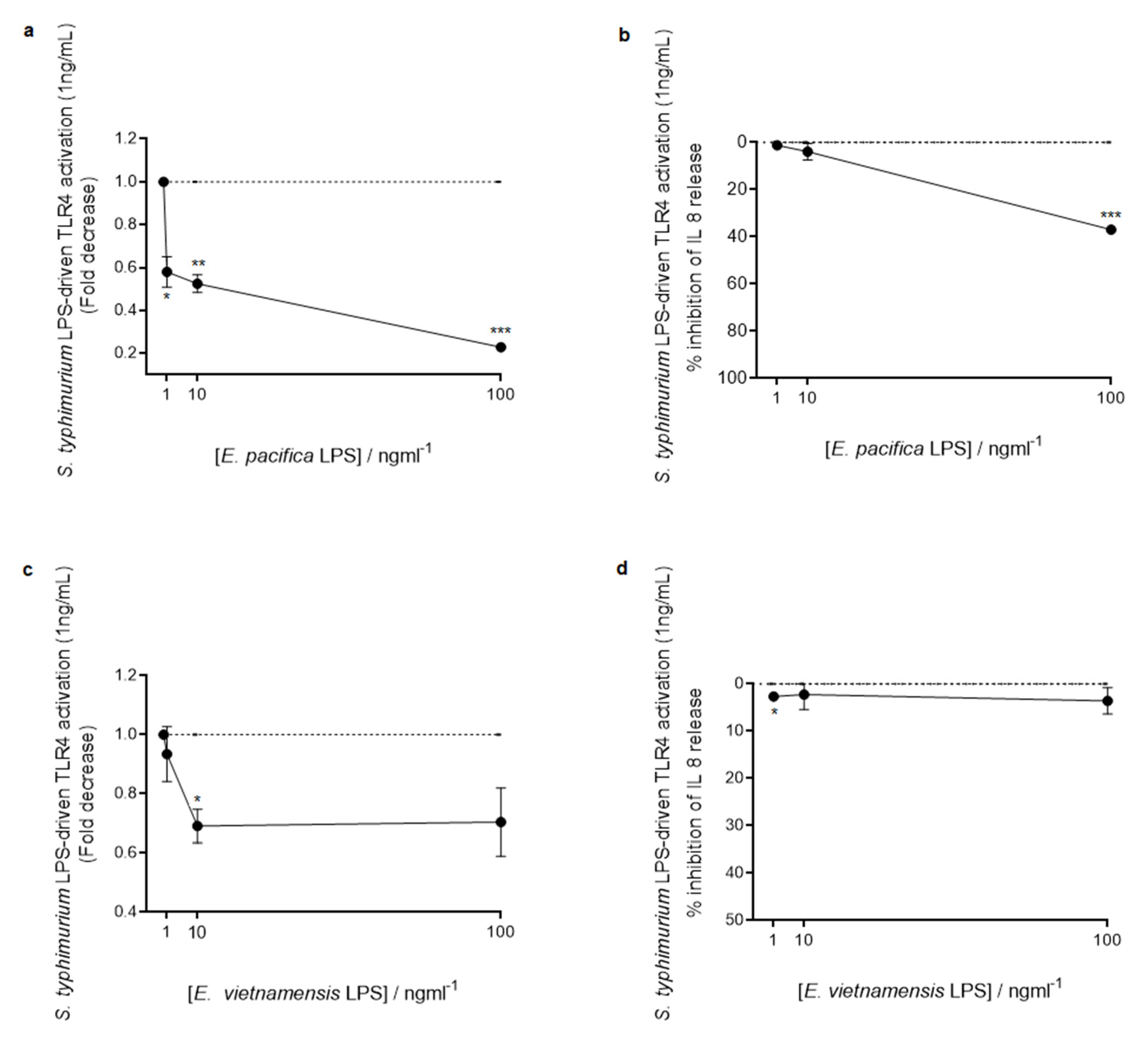
3.4. Immunological Properties of LPS from E. pacifica KMM 6172T and E. vietnamensis KMM 6221T
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Rothschild, L.J.; Mancinelli, R.L. Life in extreme environments. Nature 2001, 409, 1092–1100. [Google Scholar] [CrossRef] [PubMed]
- Debbab, A.; Aly, A.H.; Lin, W.H.; Proksch, P. Bioactive compounds from marine bacteria and fungi. Microb. Biotechn. 2010, 3, 544–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lorenzo, F.; Billod, J.-M.; Martín-Santamaría, S.; Silipo, A.; Molinaro, A. Gram-negative extremophile lipopolysaccharides: Promising source of inspiration for a new generation of endotoxin antagonists. Eur. J. Org. Chem. 2017, 2017, 4055–4073. [Google Scholar] [CrossRef] [Green Version]
- Molinaro, A.; Holst, O.; Di Lorenzo, F.; Callaghan, M.; Nurisso, A.; D’Errico, G.; Zamyatina, A.; Peri, F.; Berisio, R.; Jerala, R.; et al. Chemistry of lipid A: At the heart of innate immunity. Chemistry 2015, 21, 500–519. [Google Scholar] [CrossRef]
- Di Lorenzo, F.; Duda, K.A.; Lanzetta, R.; Silipo, A.; De Castro, C.; Molinaro, A. A journey from structure to function of bacterial lipopolysaccharides. Chem. Rev. 2021. [Google Scholar] [CrossRef]
- Di Lorenzo, F.; De Castro, C.; Lanzetta, R.; Parrilli, M.; Silipo, A.; Molinaro, A. Lipopolysaccharides as Microbe-associated Molecular Patterns: A structural perspective. In Carbohydrates in Drug Design and Discovery; Jimenez-Barbero, J., Javier-Canada, F., Martín-Santamaría, S., Eds.; RSC: Cambridge, UK, 2015; pp. 38–63. [Google Scholar]
- Kieser, K.J.; Kagan, J.C. Multi-receptor detection of individual bacterial products by the innate immune system. Nat. Rev. Immunol. 2017, 6, 376–390. [Google Scholar] [CrossRef]
- Mata-Haro, V.; Cekic, C.; Martin, M.; Chilton, P.M.; Casella, C.R.; Mitchell, T.C. The vaccine adjuvant monophosphoryl Lipid A as a TRIF-biased agonist of TLR4. Science 2007, 316, 1628–1632. [Google Scholar] [CrossRef]
- Cochet, F.; Peri, F. The role of carbohydrates in the lipopolysaccharide (LPS)/Toll-Like Receptor 4 (TLR4) signalling. Int. J. Mol. Sci. 2017, 18, 2318. [Google Scholar] [CrossRef] [Green Version]
- Di Lorenzo, F.; Palmigiano, A.; Albitar-Nehme, S.; Pallach, M.; Kokoulin, M.; Komandrova, N.; Romanenko, L.; Bernardini, M.-L.; Garozzo, D.; Molinaro, A.; et al. Lipid A structure and immunoinhibitory effect of the marine bacterium Cobetia pacifica KMM 3879T. Eur. J. Org. Chem. 2018, 20–21, 2707–2716. [Google Scholar] [CrossRef]
- Di Lorenzo, F.; Palmigiano, A.; Paciello, I.; Pallach, M.; Garozzo, D.; Bernardini, M.L.; La Cono, V.; Yakimov, M.M.; Molinaro, A.; Silipo, A. The deep-sea polyextremophile Halobacteroides lacunaris TB21 rough-type LPS: Structure and inhibitory activity towards toxic LPS. Mar. Drugs 2017, 15, 201. [Google Scholar] [CrossRef]
- Ialenti, A.; Di Meglio, P.; Grassia, G.; Maffia, P.; Di Rosa, M.; Lanzetta, R.; Molinaro, A.; Silipo, A.; Grant, W.D.; Ianaro, A. A novel lipid A from Halomonas magadiensis inhibits enteric LPS-induced human monocyte activation. Eur. J. Immunol. 2006, 36, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Nedashkovskaya, O.I.; Kim, S.B. Echinicola. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.B., Rainey, F., Kämpfer, P., Trujillo, M., Chun, J., DeVos, P., Hedlund, B., Dedysh, S., Eds.; Wiley: Hoboken, NJ, USA, 2015. [Google Scholar]
- Tomshich, S.V.; Kokoulin, M.S.; Kalinovsky, A.I.; Nedashkovskaya, O.I.; Komandrova, N.A. Structure of the O-specific polysaccharide from a marine bacterium Echinicola pacifica KMM 6172(T) containing 2,3-diacetamido-2,3-dideoxy-D-glucuronic acid. Carbohydr. Res. 2016, 425, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Tomshich, S.V.; Kokoulin, M.S.; Kalinovsky, A.I.; Komandrova, N.A.; Nedashkovskaya, O.I. Structure of the O-specific polysaccharide from a marine bacterium Echinicola vietnamensis KMM 6221T. Carbohydr. Res. 2015, 402, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Westphal, O.; Jann, K. Bacterial lipopolysaccharides. Extraction with phenol water and further applications of the procedure. In Methods in Carbohydrate Chemistry; Whistler, R.L., Ed.; Academic: New York, NY, USA, 1965; pp. 83–91. [Google Scholar]
- Kittelberger, R.; Hilbink, F. Sensitive silver-staining detection of bacterial lipopolysaccharides in polyacrylamide gels. J. Biochem. Biophys. Methods 1993, 26, 81–86. [Google Scholar] [CrossRef]
- Rietschel, E.T. Absolute configuration of 3-hydroxy fatty acids present in lipopolysaccharides from various bacterial groups. Eur. J. Biochem. 1976, 64, 423–428. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Pither, M.D.; McClean, S.; Silipo, A.; Molinaro, A.; Di Lorenzo, F. A chronic strain of the cystic fibrosis pathogen Pandoraea pulmonicola expresses a heterogenous hypo-acylated lipid A. Glycoconj. J. 2021, 38, 135–144. [Google Scholar] [CrossRef]
- Leontein, K.; Lindberg, B.; Lōnngren, J. Assignment of absolute configuration of sugars by glc of their acetylated glycosides formed from chiral alcohols. Methods Carbohydr. Chem. 1978, 62, 359–362. [Google Scholar]
- Silipo, A.; Lanzetta, R.; Amoresano, A.; Parrilli, M.; Molinaro, A. Ammonium hydroxide hydrolysis: A valuable support in the MALDI-TOF mass spectrometry analysis of Lipid A fatty acid distribution. J. Lipid Res. 2002, 43, 2188–2195. [Google Scholar] [CrossRef] [Green Version]
- Di Lorenzo, F. The lipopolysaccharide lipid A structure from the marine sponge-associated bacterium Pseudoalteromonas sp. 2A. Antonie Van Leeuwenhoek 2017, 110, 1401–1412. [Google Scholar] [CrossRef]
- Larrouy-Maumus, G.; Clements, A.; Filloux, A.; McCarthy, R.R.; Mostowy, S. Direct detection of lipid A on intact Gram-negative bacteria by MALDI-TOF mass spectrometry. J. Microbiol. Methods. 2016, 120, 68–71. [Google Scholar] [CrossRef] [Green Version]
- Domon, B.; Costello, C.E. A systematic nomenclature for carbohydrate fragmentations in FAB-MS/MS spectra of glycoconjugates. Glycoconj. J. 1988, 5, 397–409. [Google Scholar] [CrossRef]
- Di Lorenzo, F.; Palmigiano, A.; Bitar-Nehme, A.; Sturiale, L.; Duda, K.A.; Gully, L.; Lanzetta, R.; Giraud, E.; Garozzo, D.; Bernardini, M.L.; et al. The Lipid A from Rhodopseudomonas palustris strain BisA53 LPS possesses a unique structure and low immunostimulant properties. Chemistry 2017, 23, 3637–3647. [Google Scholar] [CrossRef] [PubMed]
- Darveau, R.P.; Pham, T.T.; Lemley, K.; Reife, R.A.; Bainbridge, B.W.; Coats, S.R.; Howald, W.N.; Way, S.S.; Hajjar, A.M. Porphyromonas gingivalis lipopolysaccharide contains multiple lipid A species that functionally interact with both toll-like receptors 2 and 4. Infect. Immun. 2004, 72, 5041–5051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lorenzo, F.; Pither, M.D.; Martufi, M.; Scarinci, I.; Guzmán-Caldentey, J.; Łakomiec, E.; Jachymek, W.; Bruijns, S.C.M.; Santamaría, S.M.; Frick, J.S.; et al. Pairing Bacteroides vulgatus LPS structure with its immunomodulatory effects on human cellular models. ACS Cent. Sci. 2020, 6, 1602–1616. [Google Scholar] [CrossRef] [PubMed]
- Leone, S.; Silipo, A.; Nazarenko, E.; Lanzetta, R.; Parrilli, M.; Molinaro, A. Molecular structure of endotoxins from Gram-negative marine bacteria: An update. Mar. Drugs 2007, 5, 85–112. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, F.; Silipo, A.; Matier, T.; Hanuszkiewicz, A.; Elborn, J.S.; Lanzetta, R.; Sturiale, L.; Scamporrino, A.; Garozzo, D.; Valvano, M.A.; et al. Prevotella denticola lipopolysaccharide from a cystic fibrosis isolate possesses a unique chemical structure. Eur. J. Org. Chem. 2016, 2016, 1732–1738. [Google Scholar] [CrossRef]
- Vatanen, T.; Kostic, A.D.; d’Hennezel, E.; Siljander, H.; Franzosa, E.A.; Yassour, M.; Kolde, R.; Vlamakis, H.; Arthur, T.D.; Hämäläinen, A.M.; et al. Variation in microbiome lps immunogenicity contributes to autoimmunity in humans. Cell 2016, 165, 1551. [Google Scholar] [CrossRef]
- Vorob’eva, E.V.; Krasikova, I.N.; Dmitrenok, A.S.; Dmitrenok, P.S.; Isakov, V.V.; Nedashkovskaia, O.I.; Solov’eva, T.F. An unusual lipid A from a marine bacterium Chryseobacterium scophtalmum CIP 104199(T). Bioorg. Khimiia 2006, 32, 538–545. [Google Scholar]
- Ray, B.L.; Painter, G.; Raetz, C.R. The biosynthesis of gram-negative endotoxin. Formation of lipid A disaccharides from monosaccharide precursors in extracts of Escherichia coli. J. Biol. Chem. 1984, 259, 4852–4859. [Google Scholar] [CrossRef]
- Reynolds, C.M.; Ribeiro, A.A.; McGrath, S.C.; Cotter, R.J.; Raetz, C.R.H.; Trent, M.S. An outer membrane enzyme encoded by Salmonella typhimurium lpxR that removes the 3′-acyloxyacyl moiety of lipid A. J. Biol. Chem. 2006, 281, 21974–21987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lorenzo, F.; Paciello, I.; Fazio, L.L.; Albuquerque, L.; Sturiale, L.; da Costa, M.S.; Lanzetta, R.; Parrilli, M.; Garozzo, D.; Bernardini, M.L.; et al. Thermophiles as potential source of novel endotoxin antagonists: The full structure and bioactivity of the lipo-oligosaccharide from Thermomonas hydrothermalis. ChemBioChem 2014, 15, 2146–2155. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid Component |
|---|
| 9:0 iso (3-OH) [i9:0(3-OH)] |
| 10:0(3-OH) |
| 14:0 15:1 iso (i15:1) 15:0 iso (i15:0) 15:0 anteiso (a15:0) 16:1 16:0 15:0 iso (3-OH) [i15:0(3-OH)] 15:0(3-OH) 17:0 iso (i17:0) 17:1 iso (i17:1) 16:0(3-OH) 17:0 iso 3-OH [i17:0(3-OH)] |
| Echinicola pacifica KMM 6172T | |||
| Observed Ion Peaks (m/z) | Predicted Mass (Da) | Acyl Substitution | Proposed Composition |
| 1607.8 | 1608.0 | Tetra-acyl | HexN2, P, HexA, [17:0(3-OH)]2 [15:0(3-OH)] (16:1) |
| 1621.8 | 1622.0 | Tetra-acyl | HexN2, P, HexA, [17:0(3-OH)]2 [15:0(3-OH)] (17:1) |
| 1595.8 | 1596.0 | Tetra-acyl | HexN2, P, HexA, [17:0(3-OH)]2 [15:0(3-OH)] (15:0) |
| 1527.7 | 1527.9 | Tetra-acyl | HexN2, P, HexA, [17:0(3-OH)]2 [15:0(3-OH)] [9:0(3-OH)] |
| 1431.7 | 1432.0 | Tetra-acyl | HexN2, P, [17:0(3-OH)]2 [15:0(3-OH)] (16:1) |
| 1371.6 | 1371.8 | Tri-acyl | HexN2, P, HexA, [17:0(3-OH)]2 [15:0(3-OH)] |
| Echinicola vietnamensis KMM 6221T | |||
| 1579.8 | 1580.0 | Tetra-acyl | HexN2, P, HexA, [16:0(3-OH)]2 [15:0(3-OH)] (16:1) |
| 1513.7 | 1513.9 | Tetra-acyl | HexN2, P, HexA, [16:0(3-OH)]2 [15:0(3-OH)] [10:0(3-OH)] |
| 1403.8 | 1403.9 | Tetra-acyl | HexN2, P, [16:0(3-OH)]2 [15:0(3-OH)] (16:1) |
| 1343.6 | 1343.8 | Tri-acyl | HexN2, P, HexA, [16:0(3-OH)]2 [15:0(3-OH)] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pither, M.D.; Mantova, G.; Scaglione, E.; Pagliuca, C.; Colicchio, R.; Vitiello, M.; Chernikov, O.V.; Hua, K.-F.; Kokoulin, M.S.; Silipo, A.; et al. The Unusual Lipid A Structure and Immunoinhibitory Activity of LPS from Marine Bacteria Echinicola pacifica KMM 6172T and Echinicola vietnamensis KMM 6221T. Microorganisms 2021, 9, 2552. https://doi.org/10.3390/microorganisms9122552
Pither MD, Mantova G, Scaglione E, Pagliuca C, Colicchio R, Vitiello M, Chernikov OV, Hua K-F, Kokoulin MS, Silipo A, et al. The Unusual Lipid A Structure and Immunoinhibitory Activity of LPS from Marine Bacteria Echinicola pacifica KMM 6172T and Echinicola vietnamensis KMM 6221T. Microorganisms. 2021; 9(12):2552. https://doi.org/10.3390/microorganisms9122552
Chicago/Turabian StylePither, Molly Dorothy, Giuseppe Mantova, Elena Scaglione, Chiara Pagliuca, Roberta Colicchio, Mariateresa Vitiello, Oleg V. Chernikov, Kuo-Feng Hua, Maxim S. Kokoulin, Alba Silipo, and et al. 2021. "The Unusual Lipid A Structure and Immunoinhibitory Activity of LPS from Marine Bacteria Echinicola pacifica KMM 6172T and Echinicola vietnamensis KMM 6221T" Microorganisms 9, no. 12: 2552. https://doi.org/10.3390/microorganisms9122552
APA StylePither, M. D., Mantova, G., Scaglione, E., Pagliuca, C., Colicchio, R., Vitiello, M., Chernikov, O. V., Hua, K.-F., Kokoulin, M. S., Silipo, A., Salvatore, P., Molinaro, A., & Di Lorenzo, F. (2021). The Unusual Lipid A Structure and Immunoinhibitory Activity of LPS from Marine Bacteria Echinicola pacifica KMM 6172T and Echinicola vietnamensis KMM 6221T. Microorganisms, 9(12), 2552. https://doi.org/10.3390/microorganisms9122552