
Lowering pO2 Interacts with Photoperiod to Alter Physiological Performance of the Coastal Diatom Thalassiosira pseudonana



Abstract
:1. Introduction
2. Materials and Methods
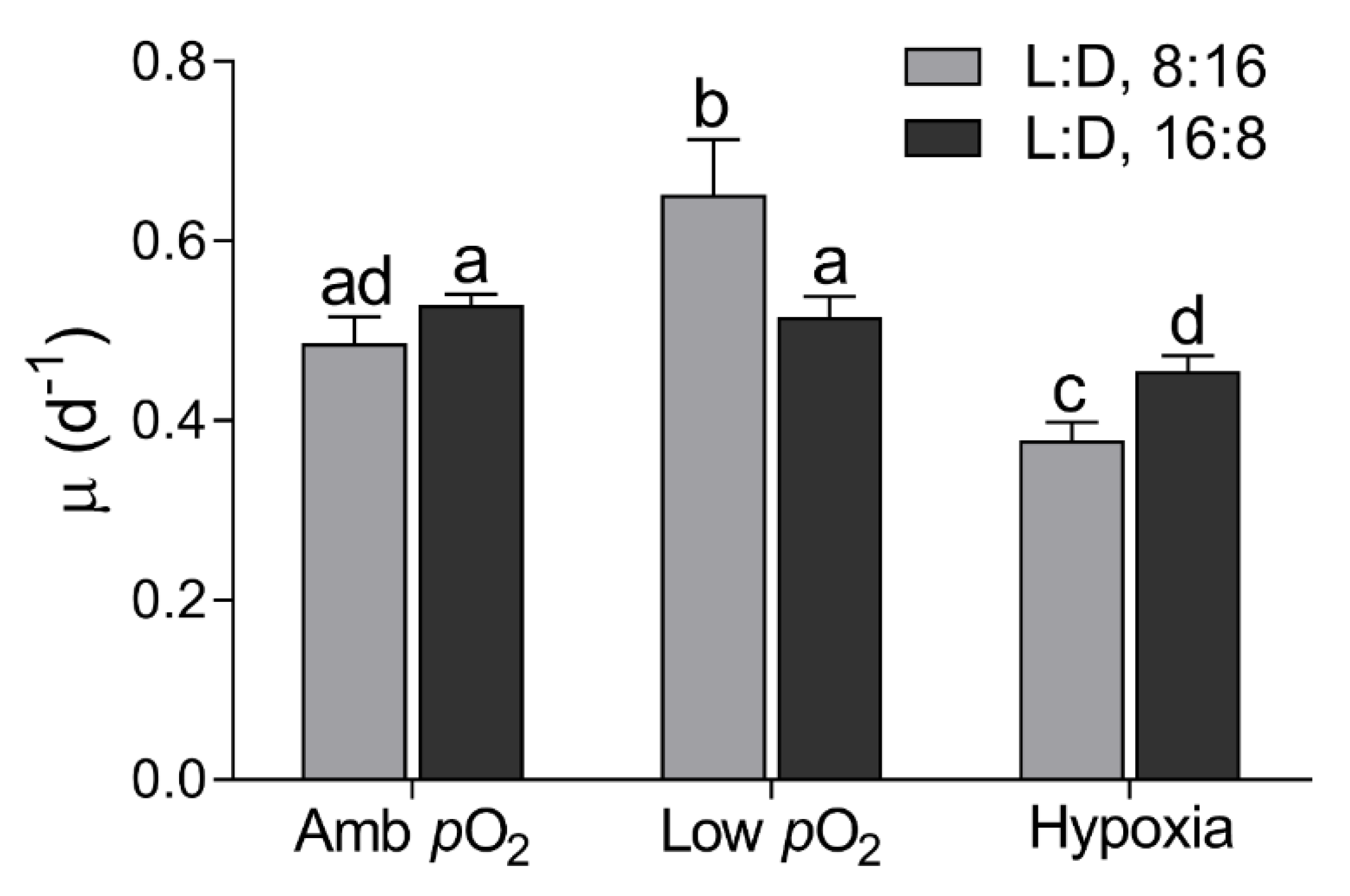
2.1. Culture Protocol and Growth Rate
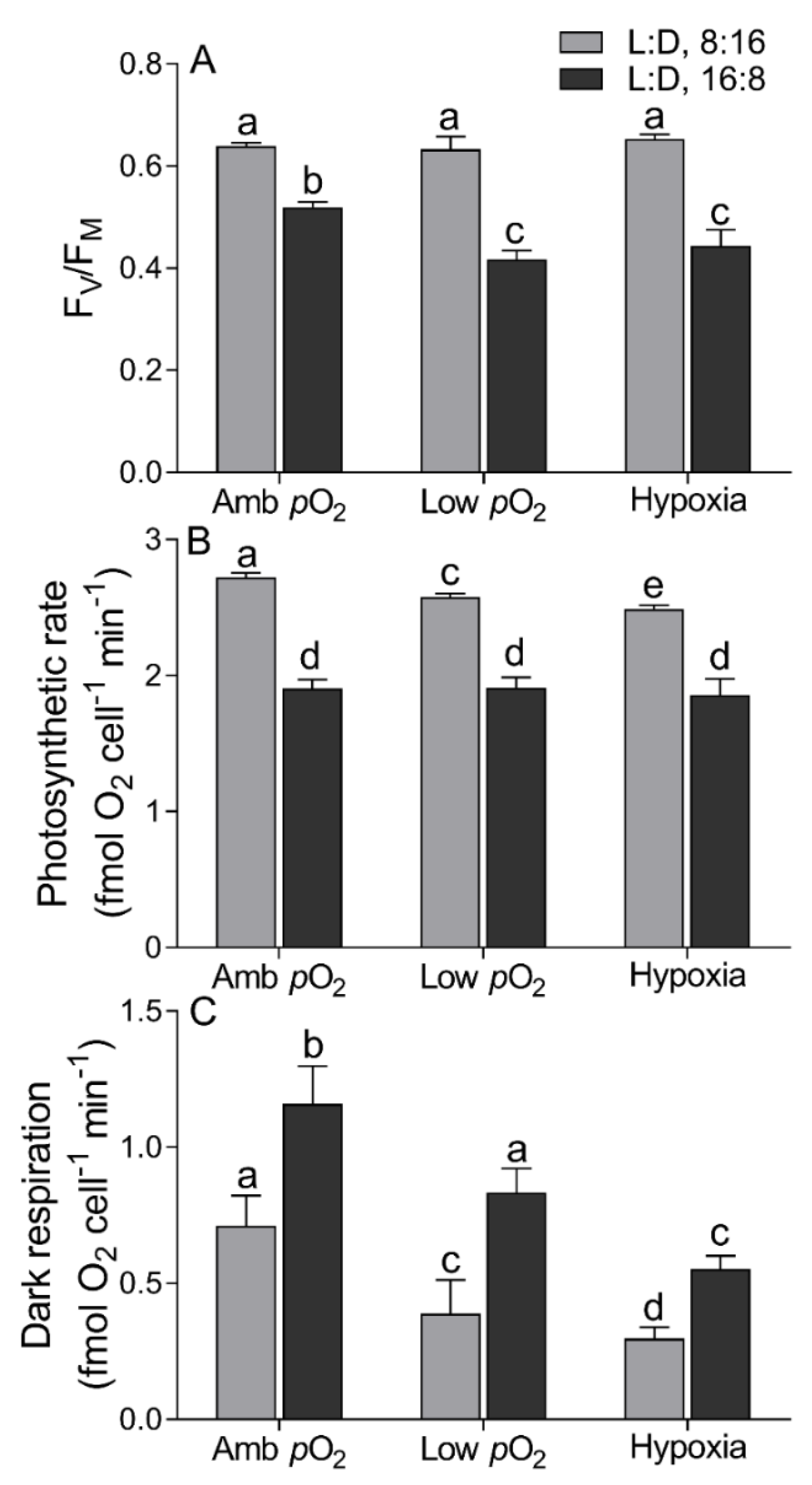
2.2. Chlorophyll Fluorescence
2.3. Photosynthetic and Respiratory Rate
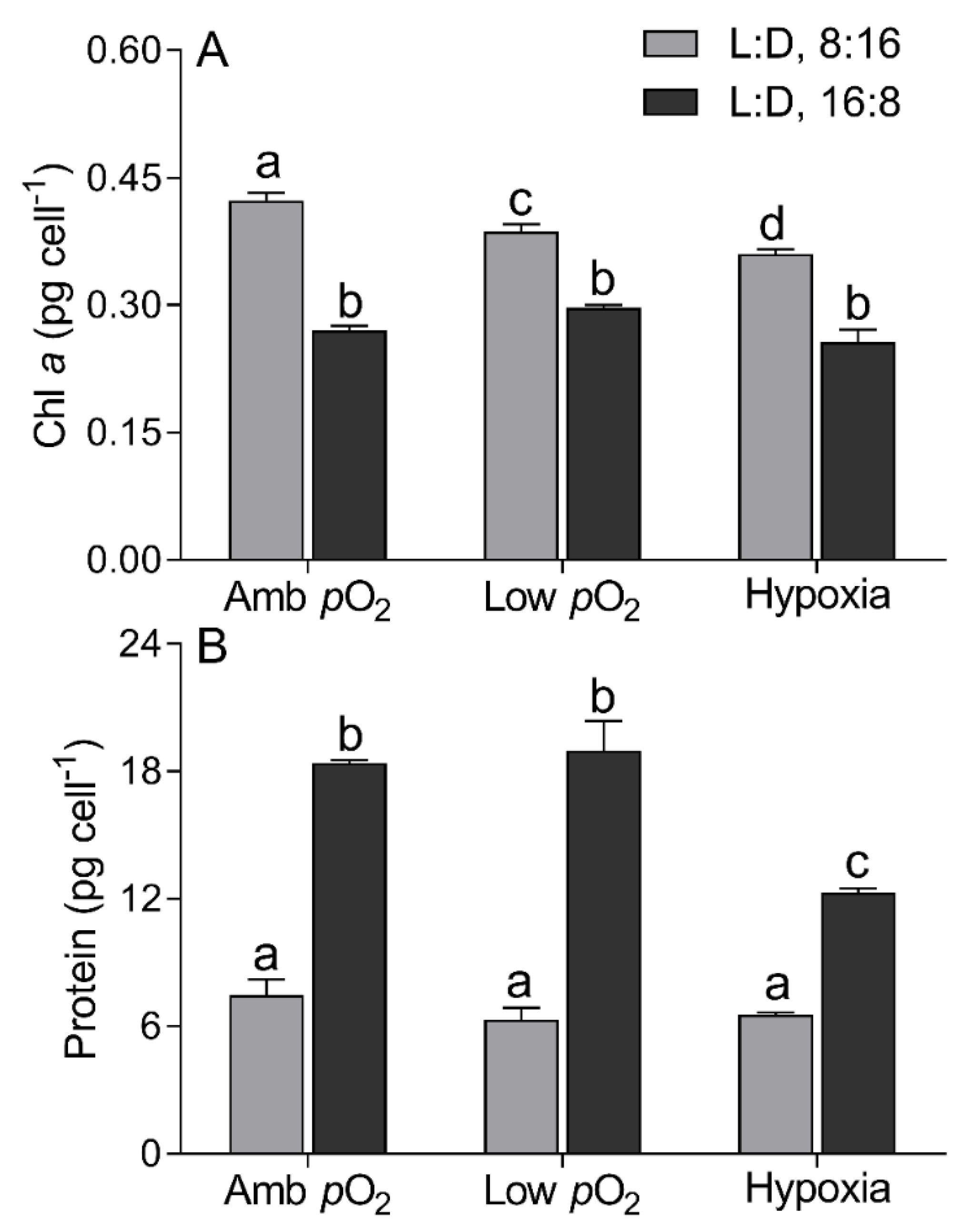
2.4. Cell Compositions
2.5. Transcriptome Sequencing and Analysis
2.6. Data Analysis
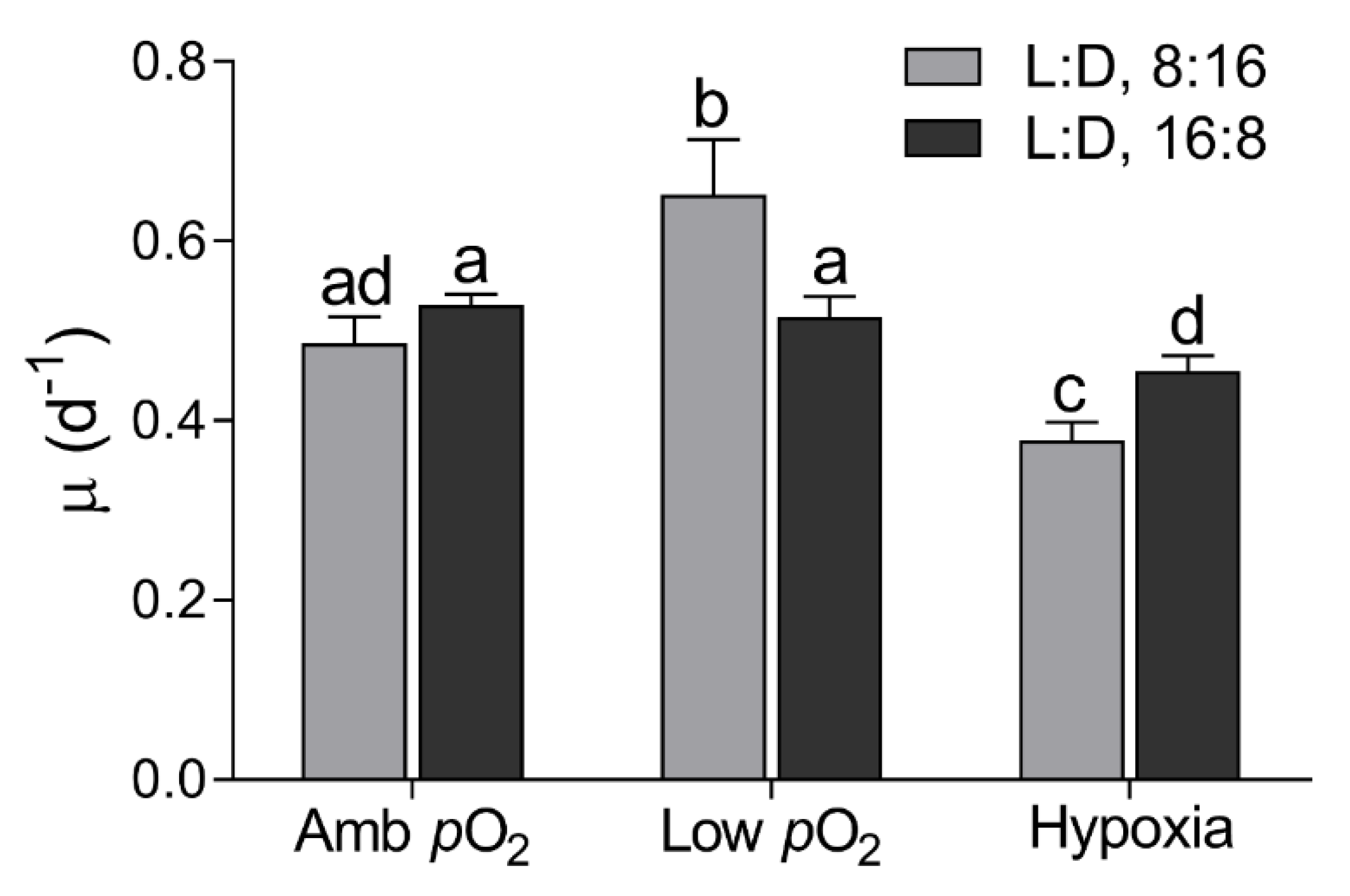
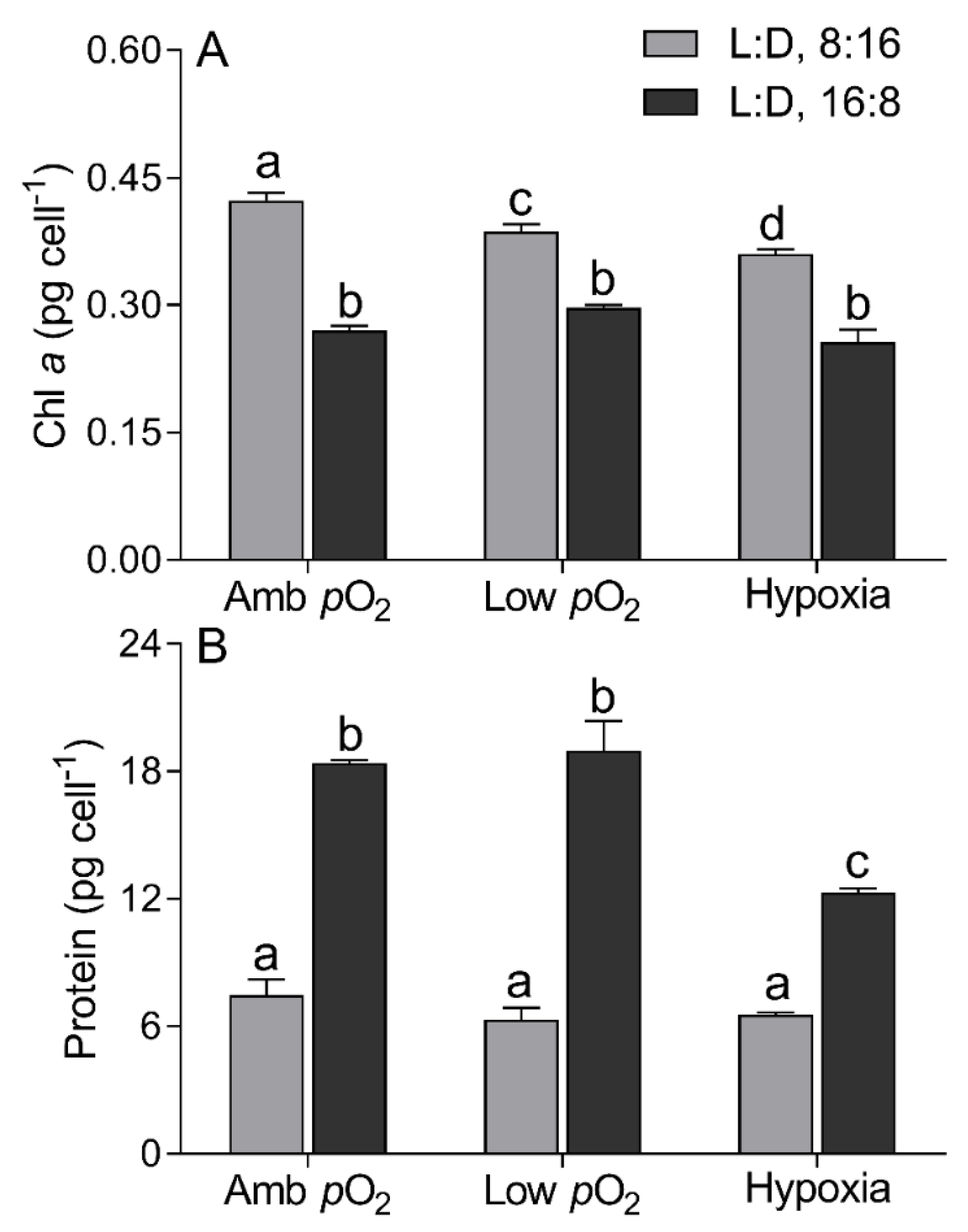
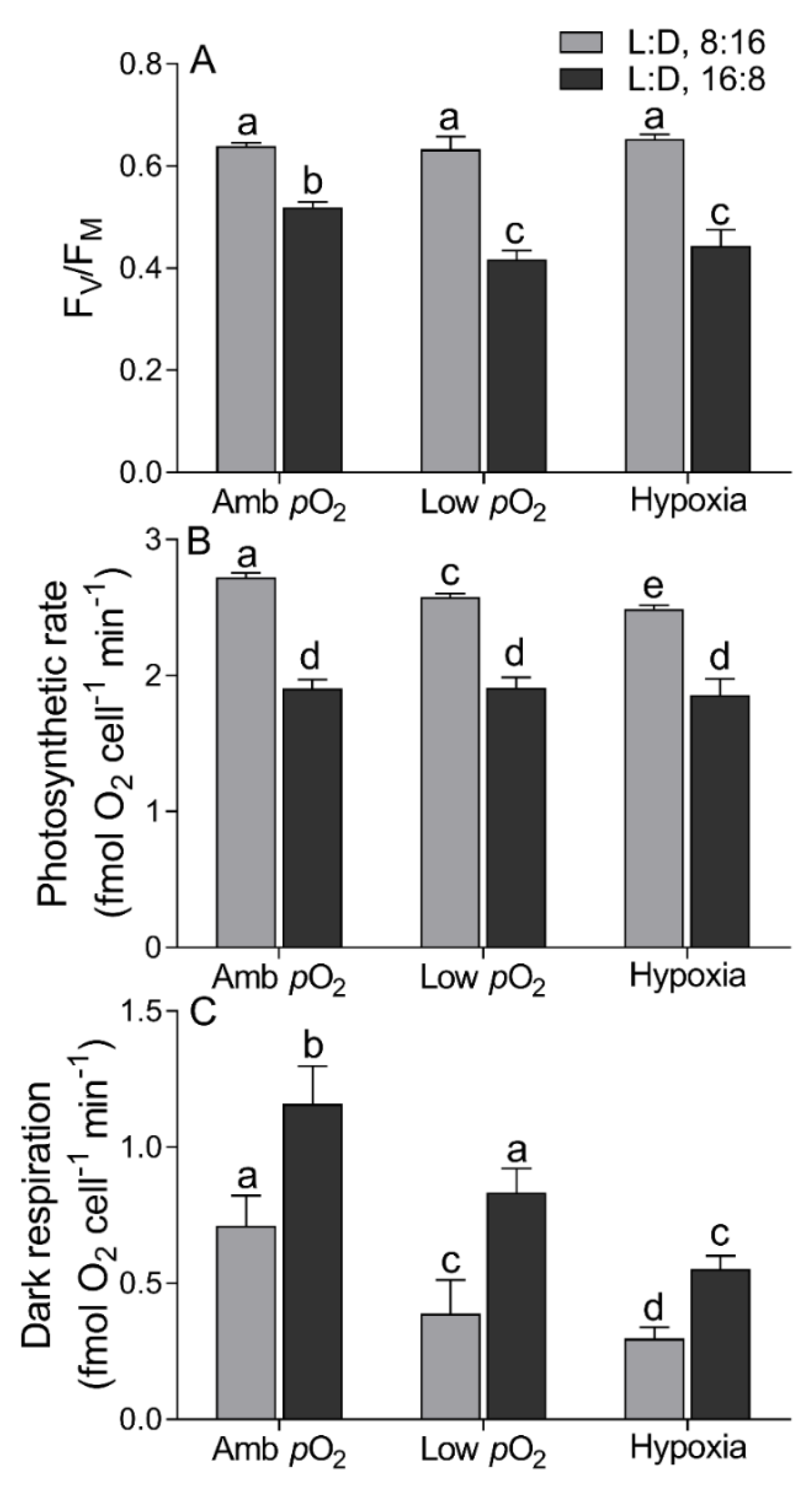
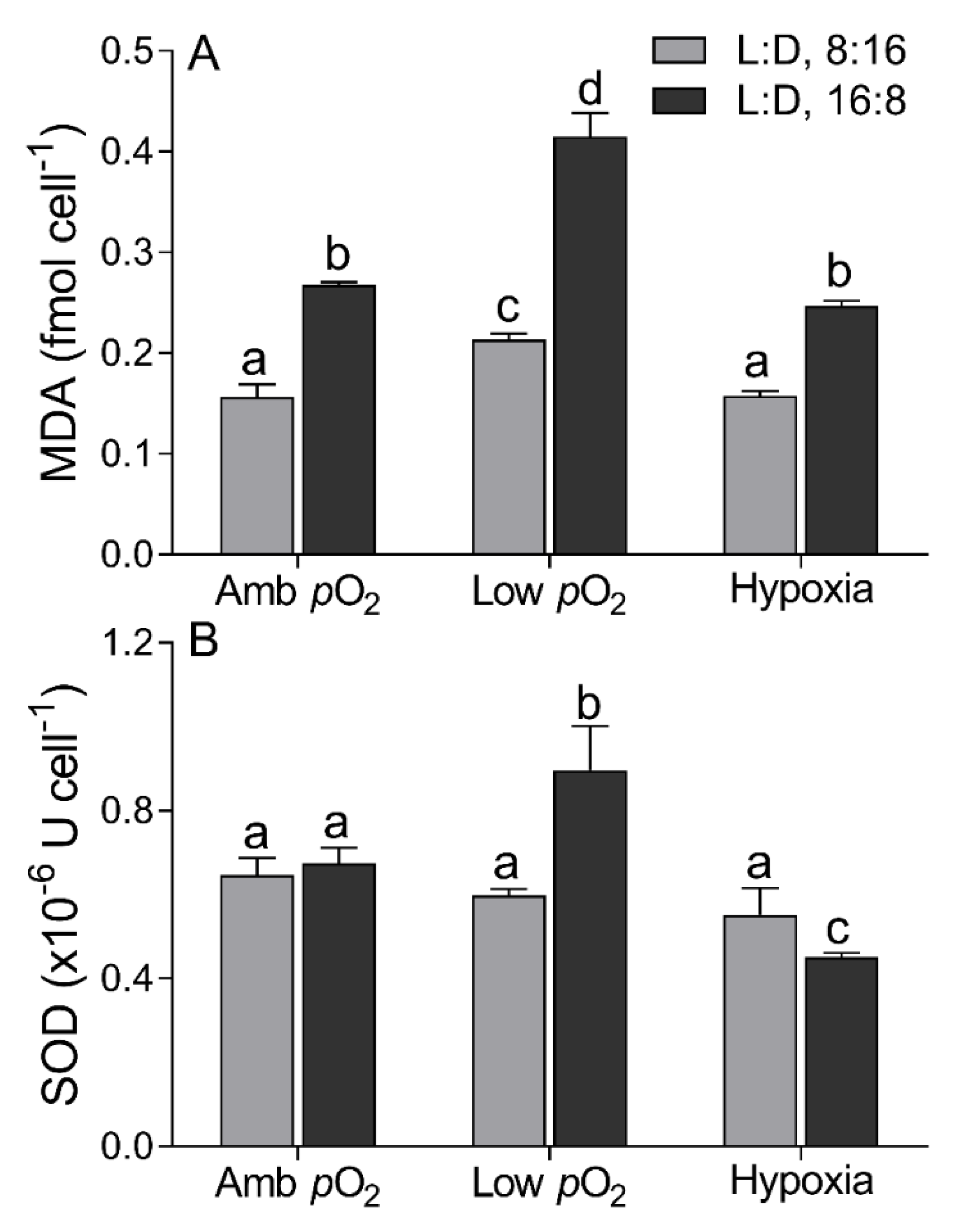
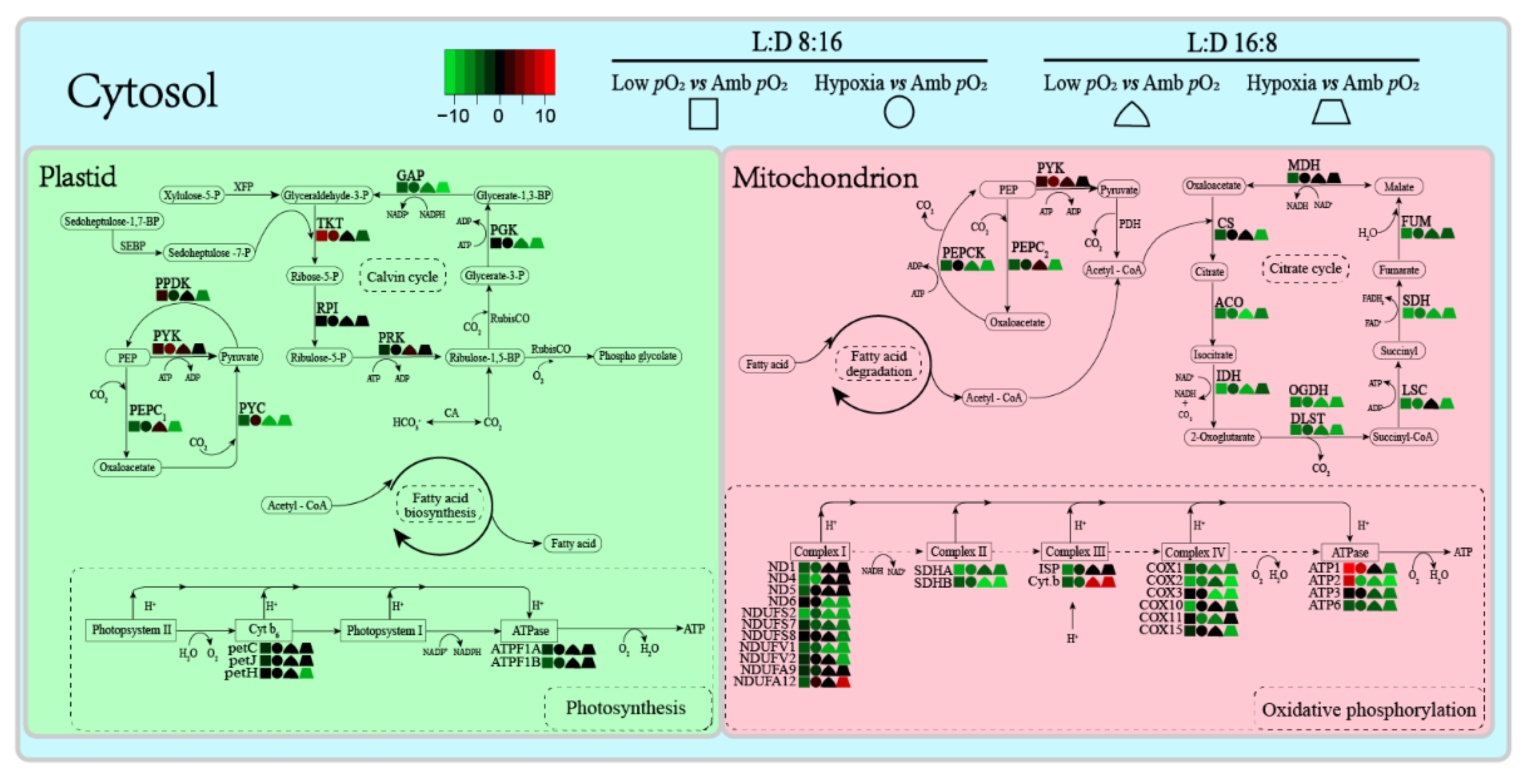
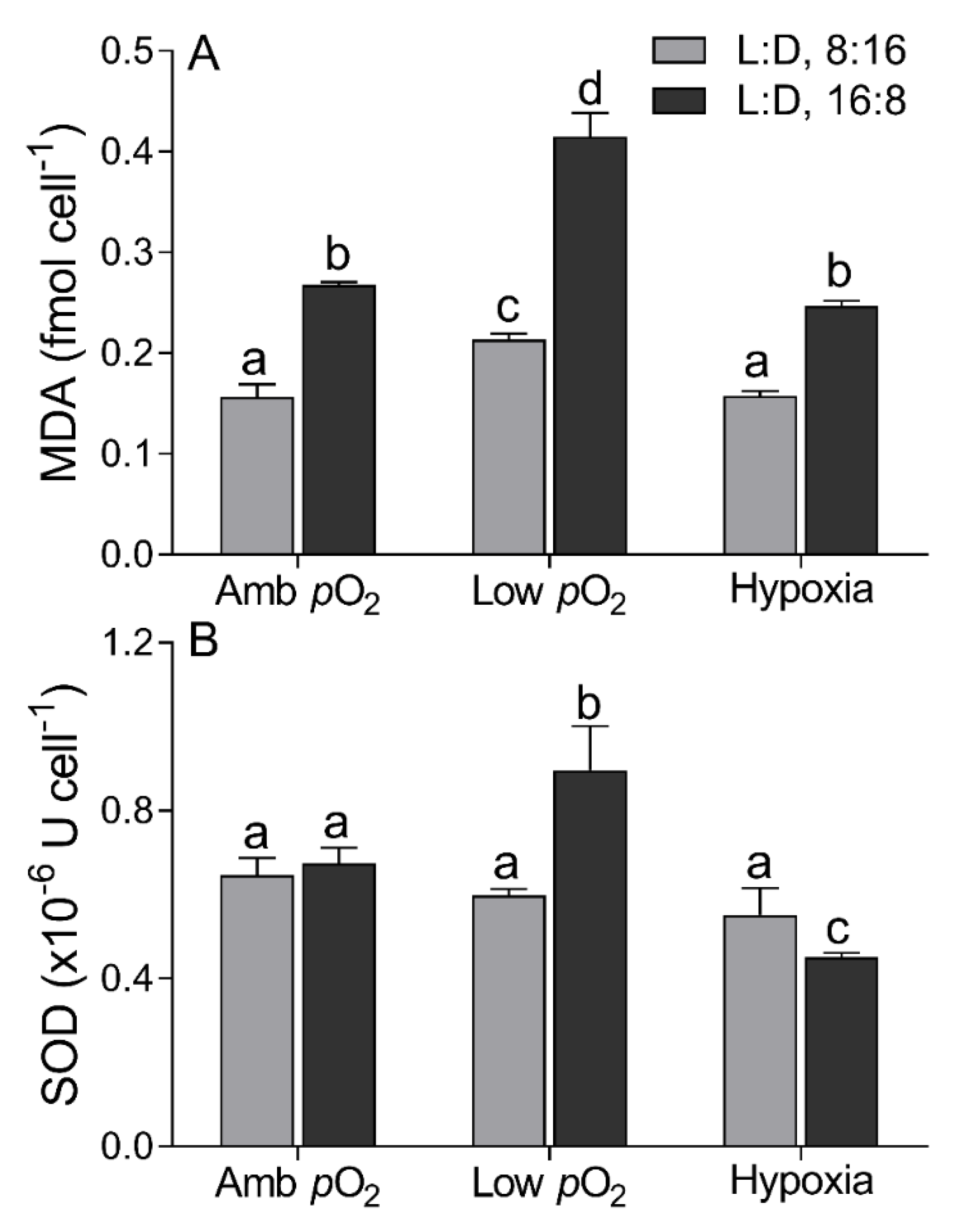
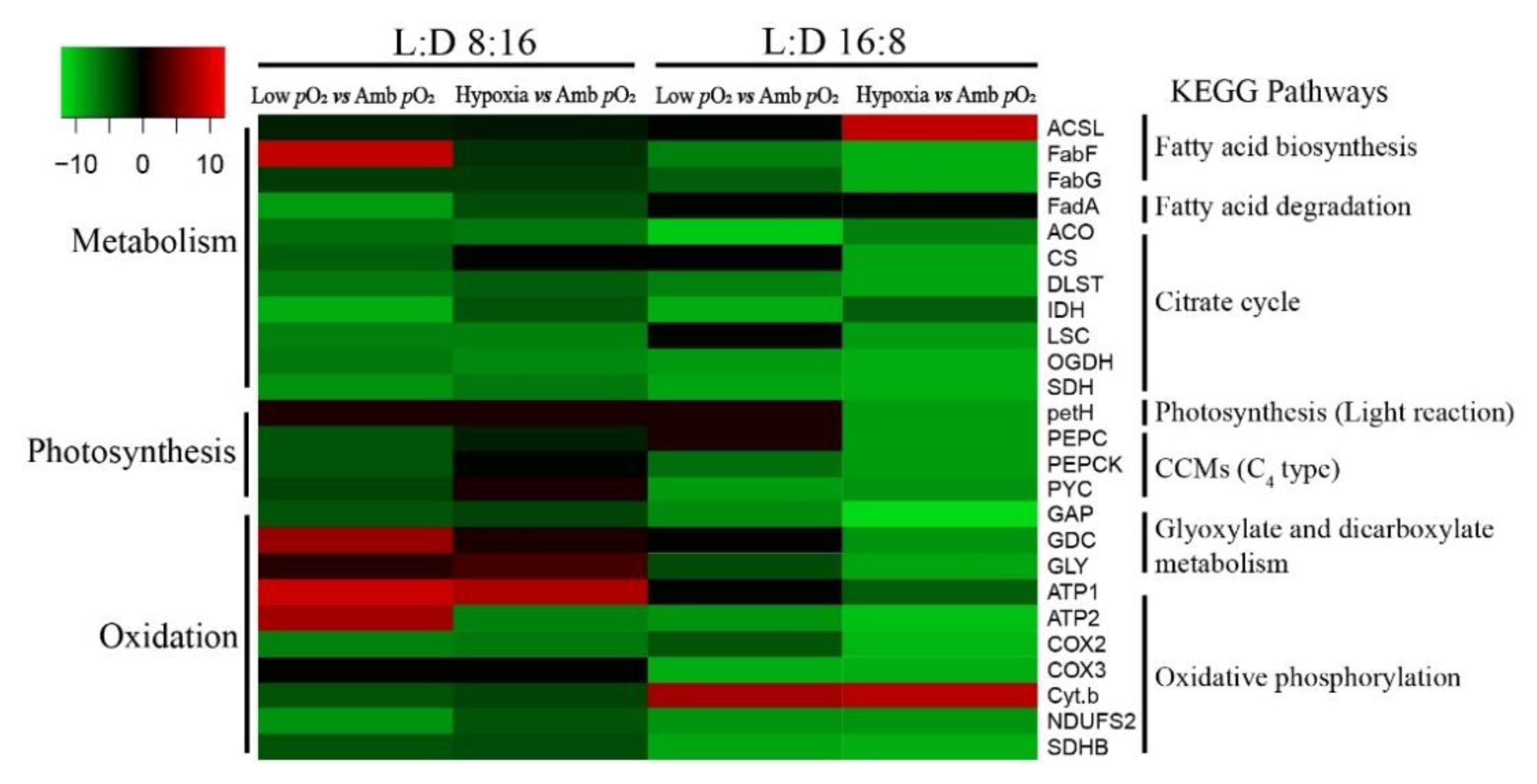
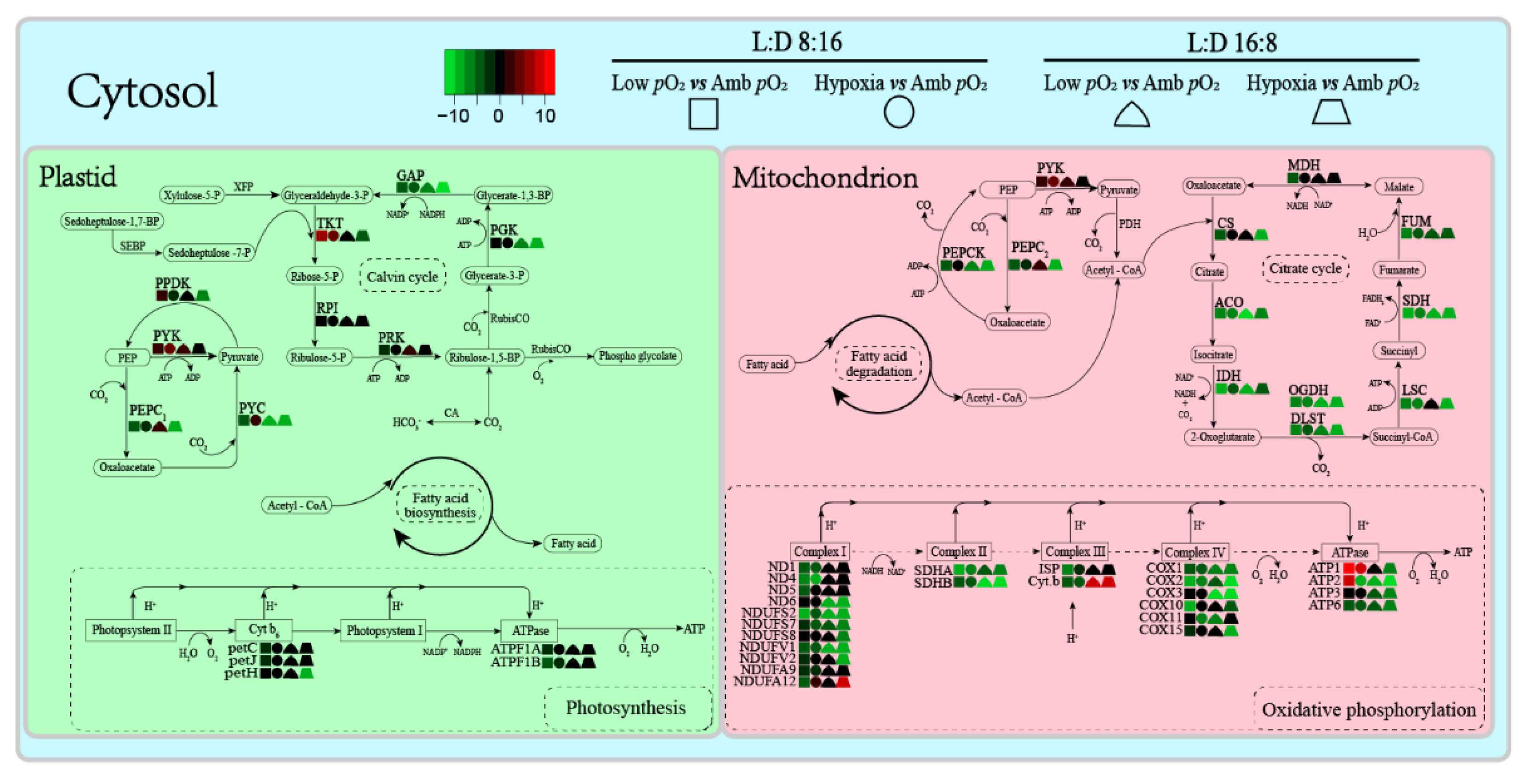
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilbert, D.; Rabalais, N.N.; Díaz, R.J.; Zhang, J. Evidence for greater oxygen decline rates in the coastal ocean than in the open ocean. Biogeosciences 2010, 7, 2283–2296. [Google Scholar] [CrossRef] [Green Version]
- Diaz, R.J.; Rosenberg, R. Spreading dead zones and consequences for marine ecosystems. Science 2008, 321, 926–929. [Google Scholar] [CrossRef]
- Schmidtko, S.; Stramma, L.; Visbeck, M. Decline in global oceanic oxygen content during the past five decades. Nature 2017, 542, 335–339. [Google Scholar] [CrossRef]
- Breitburg, D.; Levin, L.A.; Oschlies, A.; Grégoire, M.; Chavez, F.P.; Conley, D.J.; Garçon, V.; Gilbert, D.; Gutiérrez, D.; Isensee, K.; et al. Declining oxygen in the global ocean and coastal waters. Science 2018, 359, eaam7240. [Google Scholar] [CrossRef] [Green Version]
- Thomas, P.; Rahman, M.S.; Picha, M.E.; Tan, W. Impaired gamete production and viability in Atlantic croaker collected throughout the 20,000 km2 hypoxic region in the northern Gulf of Mexico. Mar. Pollut. Bull. 2015, 101, 182–192. [Google Scholar] [CrossRef]
- Wu, R.; Wo, K.T.; Chiu, J. Effects of hypoxia on growth of the diatom Skeletonema costatum. J. Exp. Mar. Biol. Ecol. 2012, 420–421, 65–68. [Google Scholar] [CrossRef]
- Haas, A.F.; Smith, J.E.; Thompson, M.; Deheyn, D.D. Effects of reduced dissolved oxygen concentrations on physiology and fluorescence of hermatypic corals and benthic algae. PeerJ 2014, 2, e235. [Google Scholar] [CrossRef]
- Kim, M.; Brodersen, K.E.; Szabó, M.; Larkum, A.W.D.; Raven, J.A.; Ralph, P.J.; Pernice, M. Low oxygen affects photophysiology and the level of expression of two-carbon metabolism genes in the seagrass Zostera muelleri. Photosynth. Res. 2018, 136, 147–160. [Google Scholar] [CrossRef]
- Garcia-Robledo, E.; Padilla, C.C.; Aldunate, M.; Stewart, F.J.; Ulloa, O.; Paulmier, A.; Gregori, G.; Revsbech, N.P. Cryptic oxygen cycling in anoxic marine zones. Proc. Natl. Acad. Sci. USA 2017, 114, 8319–8324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, M.; Beardall, J.; Raven, J.A. CO2 concentrating mechanisms in algae: Mechanisms, environmental modulation, and evolution. Ann. Rev. Plant Biol. 2005, 56, 99–131. [Google Scholar] [CrossRef] [Green Version]
- Clement, R.; Dimnet, L.; Maberly, S.C.; Gontero, B. The nature of the CO2-concentrating mechanisms in a marine diatom, Thalassiosira pseudonana. New Phytol. 2016, 209, 1417–1427. [Google Scholar] [CrossRef] [Green Version]
- Clement, R.; Lignon, S.; Mansuelle, P.; Jensen, E.; Pophillat, M.; Lebrun, R.; Denis, Y.; Puppo, C.; Maberly, S.C.; Gontero, B. Responses of the marine diatom Thalassiosira pseudonana to changes in CO2 concentration: A proteomic approach. Sci. Rep. 2017, 7, 42333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, K.; Campbell, D.A. Photophysiological responses of marine diatom to elevated CO2 and decreased pH: A review. Funct. Plant Biol. 2014, 41, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Sarthou, G.; Timmermans, K.R.; Blain, S.; Tréguer, P. Growth physiology and fate of diatoms in the ocean: A review. J. Sea Res. 2005, 53, 25–42. [Google Scholar] [CrossRef]
- Xiang, C.; Tan, Y.; Zhang, H.; Liu, J.; Ke, Z.; Li, G. The key to dinoflagellate (Noctiluca scintillans) blooming and outcompeting diatoms in winter off Pakistan, northern Arabian Sea. Sci. Total Environ. 2019, 694, 133396. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Liu, J.; Diao, Z.; Jiang, X.; Li, J.; Ke, Z.; Shen, P.; Ren, L.; Huang, L.; Tan, Y. Subsurface low dissolved oxygen occurred at fresh- and saline-water intersection of the Pearl River estuary during the summer period. Mar. Pollut. Bull. 2018, 126, 585–591. [Google Scholar] [CrossRef]
- Li, X.; Bianchi, T.S.; Yang, Z.; Osterman, L.E.; Allison, M.A.; Dimarco, S.F.; Yang, G. Historical trends of hypoxia in Changjiang River estuary: Applications of chemical biomarkers and microfossils. J. Mar. Syst. 2011, 86, 57–68. [Google Scholar] [CrossRef]
- Pruder, G.D.; Bolton, E.T. Differences between cell division and carbon fixation rates associated with light intensity and oxygen concentration: Implications in the cultivation of an estuarine diatom. Mar. Biol. 1980, 59, 1–6. [Google Scholar] [CrossRef]
- Raven, J.A.; Larkum, A. Are there ecological implications for the proposed energetic restrictions on photosynthetic oxygen evolution at high oxygen concentrations? Photosynth. Res. 2007, 94, 31–42. [Google Scholar] [CrossRef]
- Kliphuis, A.M.; Martens, D.E.; Janssen, M.; Wijffels, R.H. Effect of O2:CO2 ratio on the primary metabolism of Chlamydomonas reinhardtii. Biotechnol. Bioeng. 2011, 108, 2390–2402. [Google Scholar] [CrossRef] [PubMed]
- Vance, P.; Spalding, M.H. Growth, photosynthesis, and gene expression in Chlamydomonas over a range of CO2 concentrations and CO2/O2 ratios: CO2 regulates multiple acclimation states. Can. J. Bot. 2005, 83, 796–809. [Google Scholar] [CrossRef]
- Kamp, A.; de Beer, D.; Nitsch, J.L.; Lavik, G.; Stief, P. Diatoms respire nitrate to survive dark and anoxic conditions. Proc. Natl. Acad. Sci. USA. 2011, 108, 5649–5654. [Google Scholar] [CrossRef] [Green Version]
- Zhou, F.; Chai, F.; Huang, D.; Wells, M.; Ma, X.; Meng, Q.; Xue, H.; Xuan, J.; Wang, P.; Ni, X.; et al. Coupling and decoupling of high biomass phytoplankton production and hypoxia in a highly dynamic coastal system: The Changjiang (Yangtze River) Estuary. Front. Mar. Sci. 2020, 7, 259. [Google Scholar] [CrossRef]
- Wang, B.; Chen, J.; Jin, H.; Li, H.; Huang, D.; Cai, W.-J. Diatom bloom-derived bottom water hypoxia off the Changjiang estuary, with and without typhoon influence. Limnol. Oceanogr. 2017, 62, 1552–1569. [Google Scholar] [CrossRef]
- Yamori, W. Photosynthesis and respiration. In Plant Factory, 2nd ed.; Takagaki, T.K.N., Ed.; Academic Press: San Diego, CA, USA, 2020; pp. 197–206. [Google Scholar]
- Brand, L.E.; Guillard, R.R.L. The effects of continuous light and light intensity on the reproduction rates of twenty-two species of marine phytoplankton. J. Exp. Mar. Biol. Ecol. 1981, 50, 119–132. [Google Scholar] [CrossRef]
- Tilzer, M.M.; Dubinsky, Z. Effects of temperature and day length on the mass balance of Antarctic phytoplankton. Polar Biol. 1987, 7, 35–42. [Google Scholar] [CrossRef]
- Li, G.; Woroch, A.; Donaher, N.; Cockshutt, A.M.; Campbell, D.A. A Hard Day’s Night: Diatoms continue recycling Photosystem II in the dark. Front. Mar. Sci. 2016, 3, 218. [Google Scholar] [CrossRef] [Green Version]
- Chauton, M.S.; Winge, P.; Brembu, T.; Olav Vadstein, O.; Bones, A.M. Gene regulation of carbon fixation, storage, and utilization in the diatom Phaeodactylum tricornutum acclimated to light/dark cycles. Plant Physiol. 2013, 161, 1034–1048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Talmy, D.; Campbell, D.A. Diatom growth responses to photoperiod and light are predictable from diel reductant generation. J. Phycol. 2017, 53, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Liu, J.; Chen, B.; Li, G. Photoperiod mediates the differential physiological responses of smaller Thalassiosira pseudonana and larger Thalassiosira punctigera to temperature changes. J. Appl. Phycol. 2020, 32, 2863–2874. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, B.; Song, X.; Jian, X.; Tang, C.; Campbell, D.A.; Li, G. High antioxidant capability interacts with respiration to mediate two Alexandrium species growth exploitation of photoperiods and light intensities. Harmful Algae 2019, 82, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Ma, Q.; Xu, S.; Liu, W.; Ma, Z.; Ni, G. Opposite growth responses of Alexandrium minutum and Alexandrium catenella to photoperiods and temperatures. Plants 2021, 10, 1056. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Li, J.; Pan, X.; Jiang, H.; Sun, L.; Fu, Z. Photoperiod and temperature influence cadmium’s effects on photosynthe-sis-related gene transcription in Chlorella vulgaris. Ecotoxicol. Environ. Saf. 2010, 73, 1202–1206. [Google Scholar] [CrossRef]
- Shatwell, T.; Nicklisch, A.; Köhler, J. Temperature and photoperiod effects on phytoplankton growing under simulated mixed layer light fluctuations. Limnol. Oceanogr. 2012, 57, 541–553. [Google Scholar] [CrossRef]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [Green Version]
- Hallegraeff, G.M. Species of the diatom genus Thalassiosira in Australian water. Bot. Mar. 1984, 27, 495–513. [Google Scholar] [CrossRef]
- Armbrust, E.V.; Berges, J.A.; Bowler, C.; Green, B.R.; Martinez, D.; Putnam, N.H.; Zhou, S.; Allen, A.E.; Apt, K.E.; Bechner, M.; et al. The genome of the diatom Thalassiosira pseudonana: Ecology, evolution, and metabolism. Science 2014, 306, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Kooten, O.; Snel, J.F.H. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. BBA-Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Eilers, P.H.C.; Peeters, J.C.H. A model for the relationship between light intensity and the rate of photosynthesis in phytoplankton. Ecol. Model 1988, 42, 199–215. [Google Scholar] [CrossRef]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Qin, Z.; Xia, X.; Mai, G.; Tan, Y.; Li, G. Differential physiological responses of small Thalassiosira pseudonana and large Thalassiosira punctigera to the shifted-high light and nitrogen. J. Mar. Sci. Eng. 2021, 9, 450. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotech. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Davidson, N.M.; Oshlack, A. Corset: Enabling differential gene expression analysis for de novo assembled transcriptomes. Genome Biol. 2014, 15, 410. [Google Scholar] [PubMed] [Green Version]
- Dewey, C.N.; Li, B. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Storey, J.D. The positive false discovery rate: A Bayesian interpretation and the q-value. Ann. Stat. 2003, 31, 2013–2035. [Google Scholar] [CrossRef]
- Mao, X.; Tao, C.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Campbell, D.A. Rising CO2 interacts with growth light and growth rate to alter Photosystem II photoinactivation of the coastal diatom Thalassiosira pseudonana. PLoS ONE 2013, 8, e55562. [Google Scholar] [CrossRef]
- Spreitzer, R.J.; Salvucci, M.E. Rubisco: Structure, regulatory interactions, and possibilities for a better enzyme. Ann. Rev. Plant Biol. 2002, 53, 449–475. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, R.N.; Bopp, S.K.; Lettieri, T. Transcriptomics responses in marine diatom Thalassiosira pseudonana exposed to the polycyclic aromatic hydrocarbon benzo[a]pyrene. PLoS ONE 2011, 6, e26985. [Google Scholar] [CrossRef] [Green Version]
- Pfannschmidt, T. Chloroplast redox signals: How photosynthesis controls its own genes. Trends Plant Sci. 2003, 8, 33–41. [Google Scholar] [CrossRef] [Green Version]
- Padfield, D.; Yvon-Durocher, G.; Buckling, A.; Jennings, S.; Yvon-Durocher, G. Rapid evolution of metabolic traits explains thermal adaptation in phytoplankton. Ecol. Lett. 2015, 19, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Thny-Meyer, L.; Beck, C.; Preisig, O.; Hennecke, H. The ccoNOQP gene cluster codes for a cb-type cytochrome oxidase that functions in aerobic respiration of Rhodobacter capsulatus. Mol. Microbiol. 2010, 14, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Horken, K.M.; Tabita, F.R. Closely related form I ribulose bisphosphate carboxylase/oxygenase molecules that possess different CO2/O2 substrate specificities. Arch. Biochem. Biophys. 1999, 361, 183–194. [Google Scholar] [CrossRef]
- Papandreou, I.; Cairns, R.A.; Fontana, L.; Lim, A.L.; Denko, N.C. HIF-1 mediates adaptation to hypoxia by actively downregulating mitochondrial oxygen consumption. Cell Metab. 2006, 3, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Photosynthetic Parameters | L:D 8:16 | L:D 8:16 | ||||
|---|---|---|---|---|---|---|
| Amb pO2 | Low pO2 | Hypoxia | Amb pO2 | Low pO2 | Hypoxia | |
| α | 0.32 ± 0.01 a | 0.31 ± 0.02 a | 0.31 ± 0.04 a | 0.25 ± 0.01 b | 0.19 ± 0.04 b | 0.25 ± 0.02 b |
| EK | 469 ± 52.5 a | 582 ± 63.6 b | 499 ± 46.1 a | 200 ± 52.3 c | 230 ± 43.8 c | 230 ± 42.0 c |
| rETRmax | 148 ± 14.2 a | 179 ± 39.4 a | 159 ± 46.4 a | 49.7 ± 15.3 b | 43.8 ± 11.8 b | 57.1 ± 14.3 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, B.; Liu, J.; Xu, G.; Li, G. Lowering pO2 Interacts with Photoperiod to Alter Physiological Performance of the Coastal Diatom Thalassiosira pseudonana. Microorganisms 2021, 9, 2541. https://doi.org/10.3390/microorganisms9122541
Chen B, Liu J, Xu G, Li G. Lowering pO2 Interacts with Photoperiod to Alter Physiological Performance of the Coastal Diatom Thalassiosira pseudonana. Microorganisms. 2021; 9(12):2541. https://doi.org/10.3390/microorganisms9122541
Chicago/Turabian StyleChen, Bokun, Jihua Liu, Ge Xu, and Gang Li. 2021. "Lowering pO2 Interacts with Photoperiod to Alter Physiological Performance of the Coastal Diatom Thalassiosira pseudonana" Microorganisms 9, no. 12: 2541. https://doi.org/10.3390/microorganisms9122541
APA StyleChen, B., Liu, J., Xu, G., & Li, G. (2021). Lowering pO2 Interacts with Photoperiod to Alter Physiological Performance of the Coastal Diatom Thalassiosira pseudonana. Microorganisms, 9(12), 2541. https://doi.org/10.3390/microorganisms9122541