
ABO Blood Groups Do Not Predict Schistosoma mansoni Infection Profiles in Highly Endemic Villages of Uganda

 ,
,  ,
, 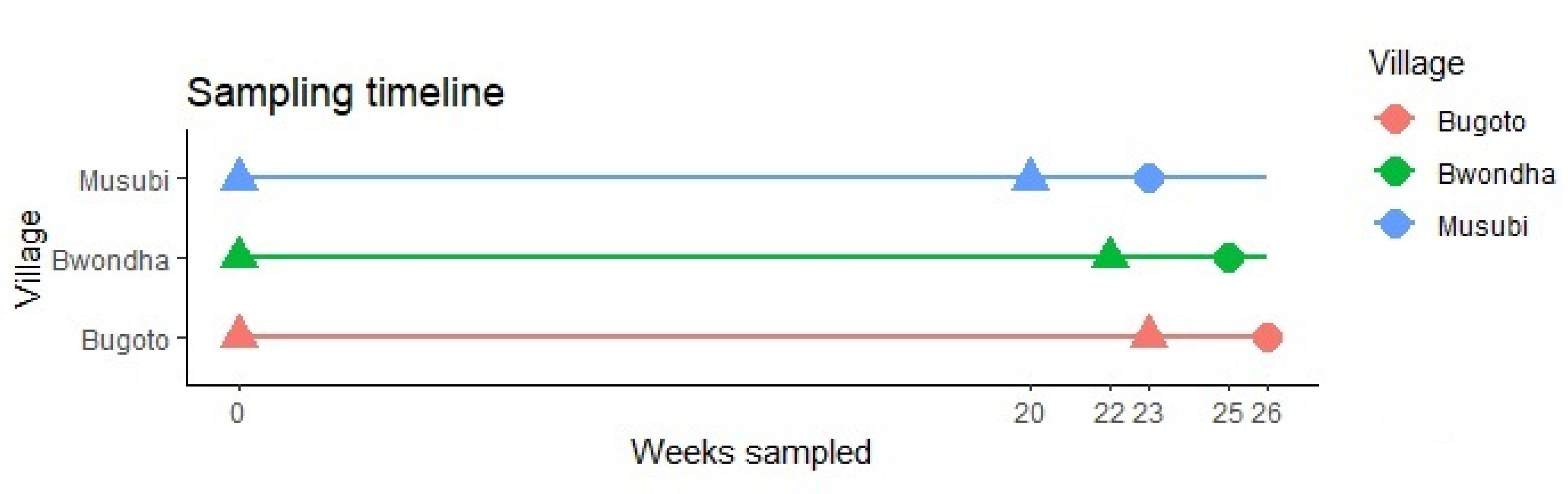{kind=link}
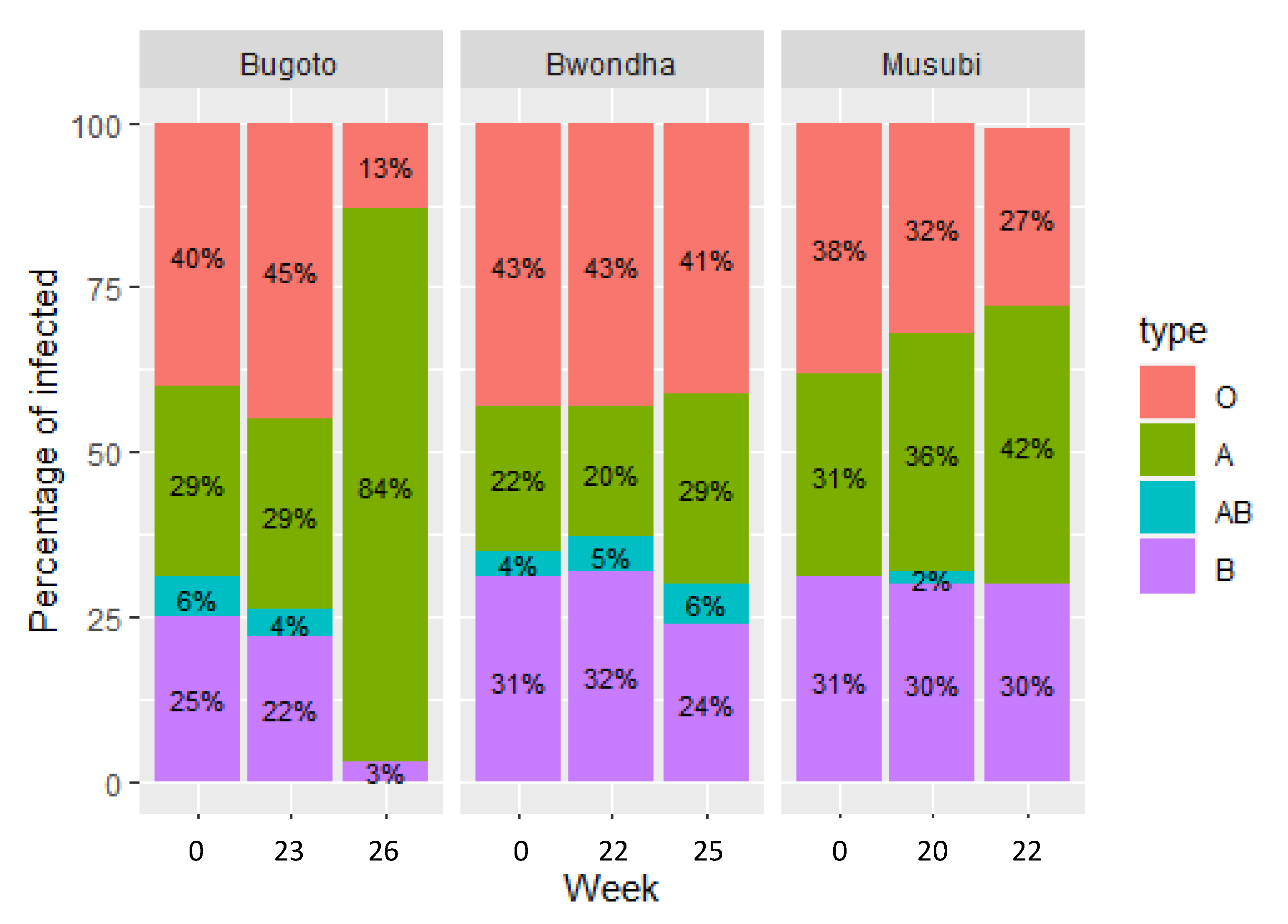{kind=link}
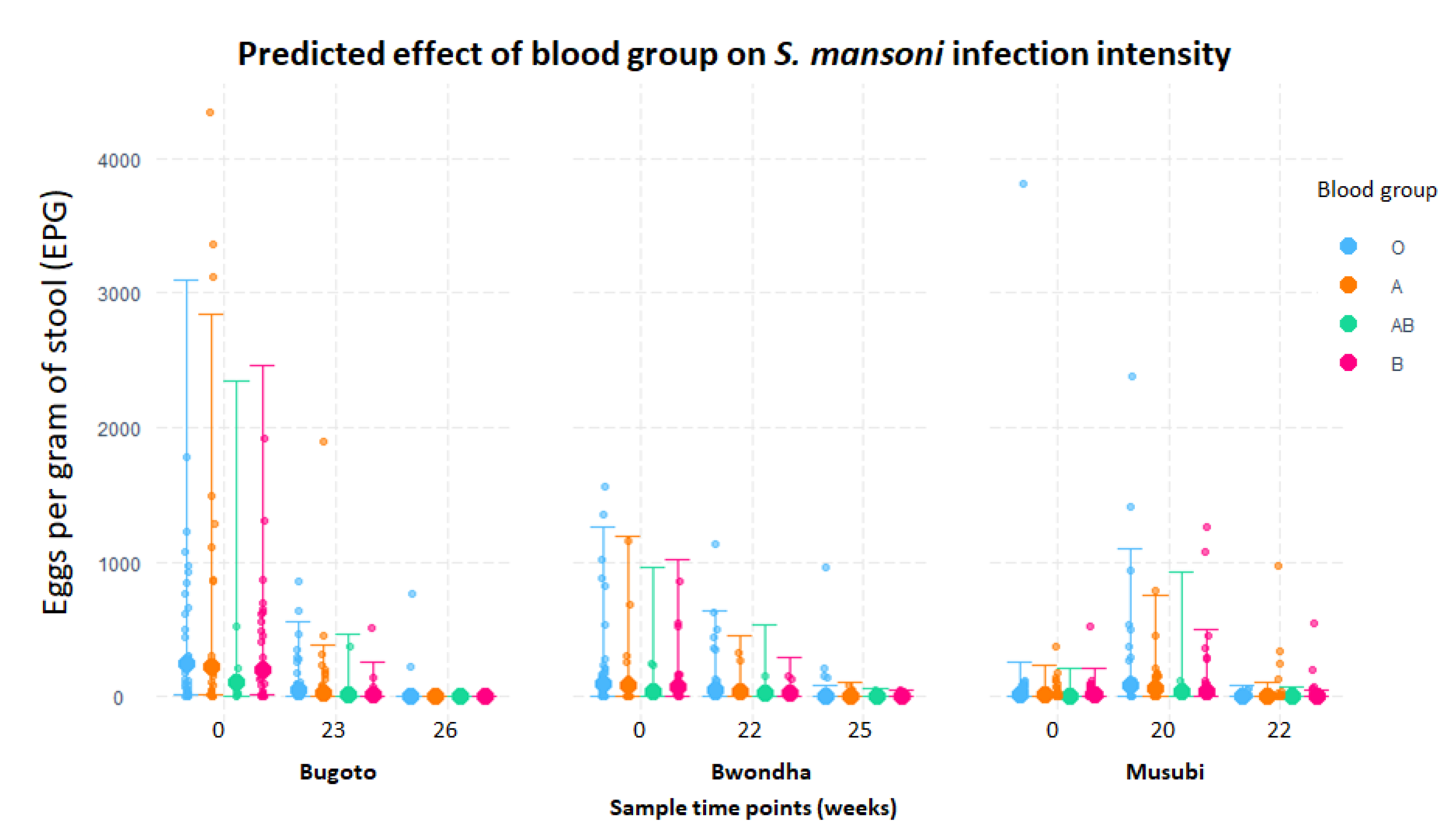{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, W.; Yang, K. The Global Status and Control of Human Schistosomiasis: An Overview. In Sino-African Cooperation for Schistosomiasis Control in Zanzibar; Springer: Berlin/Heidelberg, Germany, 2021; pp. 43–51. [Google Scholar]
- Trienekens, S.C.; Faust, C.L.; Meginnis, K.; Pickering, L.; Ericsson, O.; Nankasi, A.; Moses, A.; Tukahebwa, E.M.; Lamberton, P.H. Impacts of host gender on Schistosoma mansoni risk in rural Uganda—A mixed-methods approach. PLoS Negl. Trop. Dis. 2020, 14, e0008266. [Google Scholar] [CrossRef] [PubMed]
- Kittur, N.; Campbell, C.H., Jr.; Binder, S.; Shen, Y.; Wiegand, R.E.; Mwanga, J.R.; Kinung’hi, S.M.; Musuva, R.M.; Odiere, M.R.; Matendechero, S.H. Discovering, defining, and summarizing persistent hotspots in SCORE studies. Am. J. Trop. Med. Hyg. 2020, 103, 24–29. [Google Scholar] [CrossRef]
- Assaré, R.K.; N’Tamon, R.N.; Bellai, L.G.; Koffi, J.A.; Mathieu, T.-B.I.; Ouattara, M.; Hürlimann, E.; Coulibaly, J.T.; Diabaté, S.; N’Goran, E.K. Characteristics of persistent hotspots of Schistosoma mansoni in western Côte d’Ivoire. Parasites Vectors 2020, 13, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.W.; Kittur, N.; Binder, S.; Castleman, J.D.; Drake, J.M.; Campbell, C.H., Jr.; King, C.H.; Colley, D.G. Environmental predictors of schistosomiasis persistent hotspots following mass treatment with praziquantel. Am. J. Trop. Med. Hyg. 2020, 102, 328. [Google Scholar] [CrossRef] [PubMed]
- Mawa, P.A.; Kincaid-Smith, J.; Tukahebwa, E.M.; Webster, J.P.; Wilson, S. Schistosomiasis morbidity hotspots: Roles of the human host, the parasite and their interface in the development of severe morbidity. Front. Immunol. 2021, 12, 751. [Google Scholar] [CrossRef]
- Wiegand, R.E.; Mwinzi, P.N.; Montgomery, S.P.; Chan, Y.L.; Andiego, K.; Omedo, M.; Muchiri, G.; Ogutu, M.O.; Rawago, F.; Odiere, M.R. A persistent hotspot of Schistosoma mansoni infection in a five-year randomized trial of praziquantel preventative chemotherapy strategies. J. Infect. Dis. 2017, 216, 1425–1433. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.E.; Werkman, M.; Dunn, J.C.; Anderson, R.M. Current epidemiological evidence for predisposition to high or low intensity human helminth infection: A systematic review. Parasites Vectors 2018, 11, 65. [Google Scholar] [CrossRef] [PubMed]
- Tiongco, R.E.; Paragas, N.A.; Dominguez, M.J.; Lasta, S.L.; Pandac, J.K.; Pineda-Cortel, M.R. ABO blood group antigens may be associated with increased susceptibility to schistosomiasis: A systematic review and meta-analysis. J. Helminthol. 2018, 94, e21. [Google Scholar] [CrossRef]
- Jefferys, S.; Kenneth, C. Transfusion biology and therapy. In Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases, 6th ed.; Churchill Livingston: Philadephia, PA, USA, 2005; Volume 46, p. 708. [Google Scholar]
- Degarege, A.; Animut, A.; Medhin, G.; Legesse, M.; Erko, B. The association between multiple intestinal helminth infections and blood group, anaemia and nutritional status in human populations from Dore Bafeno, southern Ethiopia. J. Helminthol. 2014, 88, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Wokem, G.; Abah, A.; Iwuaba, P. Hookworm Infection and ABO Blood Group among Children in Aba Metropolis, Abia State, Nigeria. World Appl. Sci. J. 2016, 34, 1216–1221. [Google Scholar]
- AbdelMassih, A.F.; Mahrous, R.; Taha, A.; Saud, A.; Osman, A.; Kamel, B.; Yacoub, E.; Menshawey, E.; Ismail, H.-A.; Aita, L. The potential use of ABO blood group system for risk stratification of COVID-19. Med. Hypotheses 2020, 145, 110343. [Google Scholar] [CrossRef] [PubMed]
- Pendu, J.L.; Breiman, A.; Rocher, J.; Dion, M.; Ruvoën-Clouet, N. ABO blood types and COVID-19: Spurious, anecdotal, or truly important relationships? A reasoned review of available data. Viruses 2021, 13, 160. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Fan, H.; Lu, D.; Huang, F.; Meng, X.; Li, Z.; Tang, M.; Zhang, J.; Liu, N.; Liu, Z. Association between ABO blood groups and clinical outcome of coronavirus disease 2019: Evidence from two cohorts. medRxiv 2020. [Google Scholar] [CrossRef]
- Cabezas-Cruz, A.; Mateos-Hernández, L.; Alberdi, P.; Villar, M.; Riveau, G.; Hermann, E.; Schacht, A.-M.; Khalife, J.; Correia-Neves, M.; Gortazar, C. Effect of blood type on anti-α-Gal immunity and the incidence of infectious diseases. Exp. Mol. Med. 2017, 49, e301. [Google Scholar] [CrossRef]
- Daniels, G. Human Blood Groups; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Calafell, F.; Roubinet, F.; Ramírez-Soriano, A.; Saitou, N.; Bertranpetit, J.; Blancher, A. Evolutionary dynamics of the human ABO gene. Hum. Genet. 2008, 124, 123. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, M.; Cagliani, R.; Pozzoli, U.; Riva, S.; Comi, G.P.; Menozzi, G.; Bresolin, N.; Sironi, M. Widespread balancing selection and pathogen-driven selection at blood group antigen genes. Genome Res. 2009, 19, 199–212. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dürrbach, A.; Baple, E.; Preece, A.F.; Charpentier, B.; Gustafsson, K. Virus recognition by specific natural antibodies and complement results in MHC I cross-presentation. Eur. J. Immunol. 2007, 37, 1254–1265. [Google Scholar] [CrossRef] [PubMed]
- Esref, A.; Solmaz, I.; Akkoc, H.; Donmezdil, S.; Karahan, Z.; Safak, K.; Mertsoy, Y.; Yildirim, M.S.; Nazim, E.; Songul, A. Association between the Rh blood group and the Covid-19 susceptibility. Int. J. Hematol. Oncol. 2020, 31, 81–86. [Google Scholar]
- Hashan, M.R.; Ghozy, S.; El-Qushayri, A.E.; Pial, R.H.; Hossain, M.A.; Al Kibria, G.M. Association of dengue disease severity and blood group: A systematic review and meta-analysis. Rev. Med. Virol. 2021, 31, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ali, P.O.; Smithers, S.; Bickle, Q.; Phillips, S.; Harn, D.; Simpson, A. Analysis of the anti-Schistosoma mansoni surface antibody response during murine infection and its potential contribution to protective immunity. J. Immunol. 1988, 140, 3273–3279. [Google Scholar]
- Hokke, C.H.; Deelder, A.M. Schistosome glycoconjugates in host-parasite interplay. Glycoconj. J. 2001, 18, 573–587. [Google Scholar] [CrossRef] [PubMed]
- Goldring, O.; Clegg, J.; Smithers, S.; Terry, R. Acquisition of human blood group antigens by Schistosoma mansoni. Clin. Exp. Immunol. 1976, 26, 181. [Google Scholar]
- Mulvenna, J.; Moertel, L.; Jones, M.K.; Nawaratna, S.; Lovas, E.M.; Gobert, G.N.; Colgrave, M.; Jones, A.; Loukas, A.; McManus, D.P. Exposed proteins of the Schistosoma japonicum tegument. Int. J. Parasitol. 2010, 40, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Braschi, S.; Wilson, R.A. Proteins exposed at the adult schistosome surface revealed by biotinylation. Mol. Cell. Proteom. 2006, 5, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Dean, D.A. Schistosoma mansoni: Adsorption of human blood group A and B antigens by schistosomula. J. Parasitol. 1974, 60, 260–263. [Google Scholar] [CrossRef] [PubMed]
- Haseeb, M.; Thors, C.; Linder, E.; Eveland, L. Schistosoma mansoni: Chemoreception through N-acetyl-D-galactosamine-containing receptors in females offers insight into increased severity of schistosomiasis in individuals with blood group A. Exp. Parasitol. 2008, 119, 67–73. [Google Scholar] [CrossRef]
- Addisu, A.; Tekeste, Z.; Teshome, A.; Alemu, A.; Tesfa, H. The Association between Abo Blood Groups and Intestinal Schistosomiasis among Masero Primary School Children in Sanja, Northwest Ethiopia. J. Trop. Dis. 2016, 4, 2. [Google Scholar] [CrossRef]
- Camus, D.; Bina, J.; Carlier, Y.; Santoro, F. A, B, O blood groups and clinical forms of schistosomiasis mansoni. Trans. R. Soc. Trop. Med. Hyg. 1977, 71, 182. [Google Scholar] [CrossRef]
- Khattab, M.; El-Gengehy, M.; Sharaf, M. ABO blood groups in bilharzial hepatic fibrosis. J. Egypt. Med. Assoc. 1968, 51, 245–250. [Google Scholar] [PubMed]
- Wang, C.; Gong, W.C.; Qun, W.Q.; Wen, Z.Y.; Yu, H.P. ABO blood group and late schistosomiasis japonica. Chin. Med. J. 1983, 96, 370. [Google Scholar] [PubMed]
- Ndamba, J.; Gomo, E.; Nyazema, N.; Makaza, N.; Kaondera, K. Schistosomiasis infection in relation to the ABO blood groups among school children in Zimbabwe. Acta Trop. 1997, 65, 181–190. [Google Scholar] [CrossRef]
- Trangle, K.; Goluska, M.; O’Leary, M.; Douglas, S. Distribution of blood groups and secretor status in schistosomiasis. Parasite Immunol. 1979, 1, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Katz, N.; Tavares, J.; Aerantes, W. ABO and Rh. blood groups from patients with hepatosplenic schistosomiasis mansoni. J. Parasitol. 1967, 53, 99. [Google Scholar] [CrossRef]
- Kassim, O.; Ejezie, G. ABO blood groups in malaria and schistosomiasis haematobium. Acta Trop. 1982, 39, 179–184. [Google Scholar]
- Feldmeier, H.; Poggensee, G. Diagnostic techniques in schistosomiasis control. A review. Acta Trop. 1993, 52, 205–220. [Google Scholar] [CrossRef]
- Katz, N.; Chaves, A.; Pellegrino, J. A simple device for quantitative stool thick-smear technique in schistosomiasis mansoni. Rev. Inst. Med. Trop. Sao Paulo 1972, 14, 397–400. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Bolker, B.M. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; Version 3.5.2; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R Package Version 0.2. 2019. Available online: https://cran.r-project.org/web/packages/DHARMa/vignettes/DHARMa.html (accessed on 26 November 2021).
- Jahanpour, O.; Pyuza, J.J.; Ntiyakunze, E.O.; Mremi, A.; Shao, E.R. ABO and Rhesus blood group distribution and frequency among blood donors at Kilimanjaro Christian Medical Center, Moshi, Tanzania. BMC Res. Notes 2017, 10, 738. [Google Scholar] [CrossRef]
- Apecu, R.O.; Mulogo, E.M.; Bagenda, F.; Byamungu, A. ABO and Rhesus (D) blood group distribution among blood donors in rural south western Uganda: A retrospective study. BMC Res. Notes 2016, 9, 513. [Google Scholar] [CrossRef] [PubMed]
- De Onis, M. WHO Child Growth Standards; WHO: Geneva, Switzerland, 2006; pp. 1–336. [Google Scholar]
- Adriko, M.; Faust, C.L.; Carruthers, L.V.; Moses, A.; Tukahebwa, E.M.; Lamberton, P.H. Low praziquantel treatment coverage for Schistosoma mansoni in Mayuge District, Uganda, due to the absence of treatment opportunities, rather than systematic non-compliance. Trop. Med. Infect. Dis. 2018, 3, 111. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.; Arinaitwe, M.; Nankasi, A.; Faust, C.L.; Moses, A.; Ajambo, D.; Besigye, F.; Atuhaire, A.; Wamboko, A.; Carruthers, L.; et al. Reconciling egg-and antigen-based estimates of Schistosoma mansoni clearance and reinfection: A modelling study. Clin. Infect. Dis. 2021, ciab679. [Google Scholar] [CrossRef]
- Liu, C.; Lu, L.; Zhang, L.; Bai, Y.; Medina, A.; Rozelle, S.; Smith, D.S.; Zhou, C.; Zang, W. More poop, more precision: Improving epidemiologic surveillance of soil-transmitted helminths with multiple fecal sampling using the Kato–Katz technique. Am. J. Trop. Med. Hyg. 2017, 97, 870–875. [Google Scholar] [CrossRef] [PubMed]
- Abla, N.; Keiser, J.; Vargas, M.; Reimers, N.; Haas, H.; Spangenberg, T. Evaluation of the pharmacokinetic-pharmacodynamic relationship of praziquantel in the Schistosoma mansoni mouse model. PLoS Negl. Trop. Dis. 2017, 11, e0005942. [Google Scholar] [CrossRef] [PubMed]
- Enk, M.J.; Lima, A.C.L.; Drummond, S.C.; Schall, V.T.; Coelho, P.M.Z. The effect of the number of stool samples on the observed prevalence and the infection intensity with Schistosoma mansoni among a population in an area of low transmission. Acta Trop. 2008, 108, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Berhe, N.; Medhin, G.; Erko, B.; Smith, T.; Gedamu, S.; Bereded, D.; Moore, R.; Habte, E.; Redda, A.; Gebre-Michael, T. Variations in helminth faecal egg counts in Kato–Katz thick smears and their implications in assessing infection status with Schistosoma mansoni. Acta Trop. 2004, 92, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Milner, T.; Reilly, L.; Nausch, N.; Midzi, N.; Mduluza, T.; Maizels, R.; Mutapi, F. Circulating cytokine levels and antibody responses to human Schistosoma haematobium: IL-5 and IL-10 levels depend upon age and infection status. Parasite Immunol. 2010, 32, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Mutapi, F.; Burchmore, R.; Mduluza, T.; Midzi, N.; Turner, C.M.R.; Maizels, R.M. Age-related and infection intensity-related shifts in antibody recognition of defined protein antigens in a schistosome-exposed population. J. Infect. Dis. 2008, 198, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, M.K.; Halaly, S.; Awadalla, H.; Abdelrahman, A.; Balla, S. Prevalence, risk factors and effect of urinary schistosomiasis on academic performance of school children Age 6–15 years in Asalaya locality, White Nile State, Sudan 2017. J. Adv. Med. Med. Res. 2018, 1–7. [Google Scholar] [CrossRef]
- Gryseels, B.; Polman, K.; Clerinx, J.; Kestens, L. Human schistosomiasis. Lancet 2006, 368, 1106–1118. [Google Scholar] [CrossRef]
- Mitchell, K.M.; Mutapi, F.; Mduluza, T.; Midzi, N.; Savill, N.J.; Woolhouse, M.E.J. Predicted impact of mass drug administration on the development of protective immunity against Schistosoma haematobium. PLoS Negl. Trop. Dis. 2014, 8, e3059. [Google Scholar] [CrossRef]
- Bala, A.; Ladan, M.; Mainasara, M. Prevalence and intensity of urinary schistosomiasis in Abarma village, Gusau, Nigeria: A preliminary investigation. Sci. World J. 2012, 7, 1–4. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francoeur, R.; Atuhaire, A.; Arinaitwe, M.; Adriko, M.; Ajambo, D.; Nankasi, A.; Babayan, S.A.; Lamberton, P.H.L. ABO Blood Groups Do Not Predict Schistosoma mansoni Infection Profiles in Highly Endemic Villages of Uganda. Microorganisms 2021, 9, 2448. https://doi.org/10.3390/microorganisms9122448
Francoeur R, Atuhaire A, Arinaitwe M, Adriko M, Ajambo D, Nankasi A, Babayan SA, Lamberton PHL. ABO Blood Groups Do Not Predict Schistosoma mansoni Infection Profiles in Highly Endemic Villages of Uganda. Microorganisms. 2021; 9(12):2448. https://doi.org/10.3390/microorganisms9122448
Chicago/Turabian StyleFrancoeur, Rachel, Alon Atuhaire, Moses Arinaitwe, Moses Adriko, Diana Ajambo, Andrina Nankasi, Simon A. Babayan, and Poppy H. L. Lamberton. 2021. "ABO Blood Groups Do Not Predict Schistosoma mansoni Infection Profiles in Highly Endemic Villages of Uganda" Microorganisms 9, no. 12: 2448. https://doi.org/10.3390/microorganisms9122448
APA StyleFrancoeur, R., Atuhaire, A., Arinaitwe, M., Adriko, M., Ajambo, D., Nankasi, A., Babayan, S. A., & Lamberton, P. H. L. (2021). ABO Blood Groups Do Not Predict Schistosoma mansoni Infection Profiles in Highly Endemic Villages of Uganda. Microorganisms, 9(12), 2448. https://doi.org/10.3390/microorganisms9122448