
Effect of Carbon Sources on Pyrite-Arsenopyrite Concentrate Bio-oxidation and Growth of Microbial Population in Stirred Tank Reactors


Abstract
:1. Introduction
2. Materials and Methods
2.1. Concentrate
2.2. Experimental Setup and Biooxidaton
2.3. Sampling and Analysis
2.4. Microbial Population Analysis
2.5. Data Processing
3. Results
3.1. Biooxidation under Mesophilic Conditons
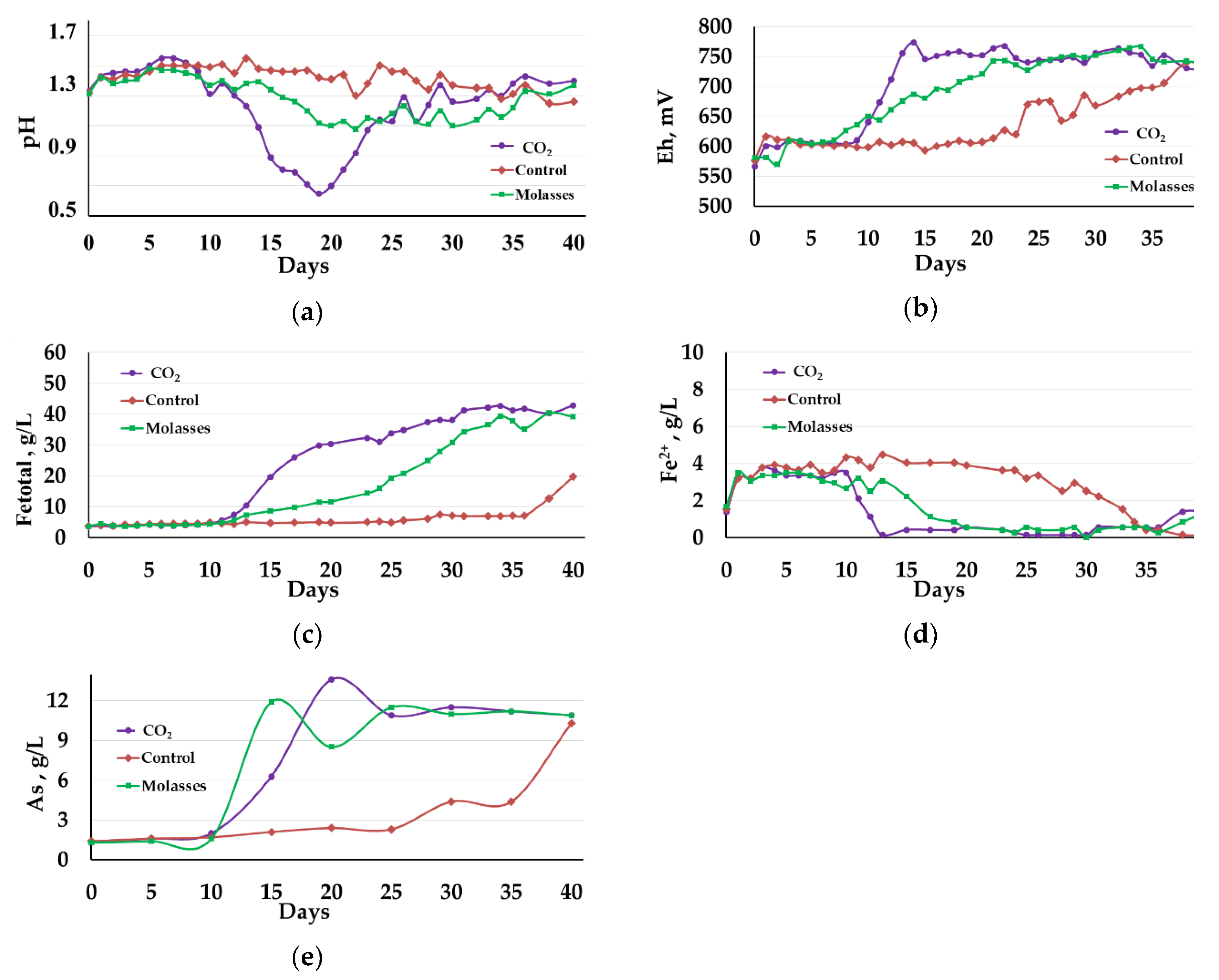
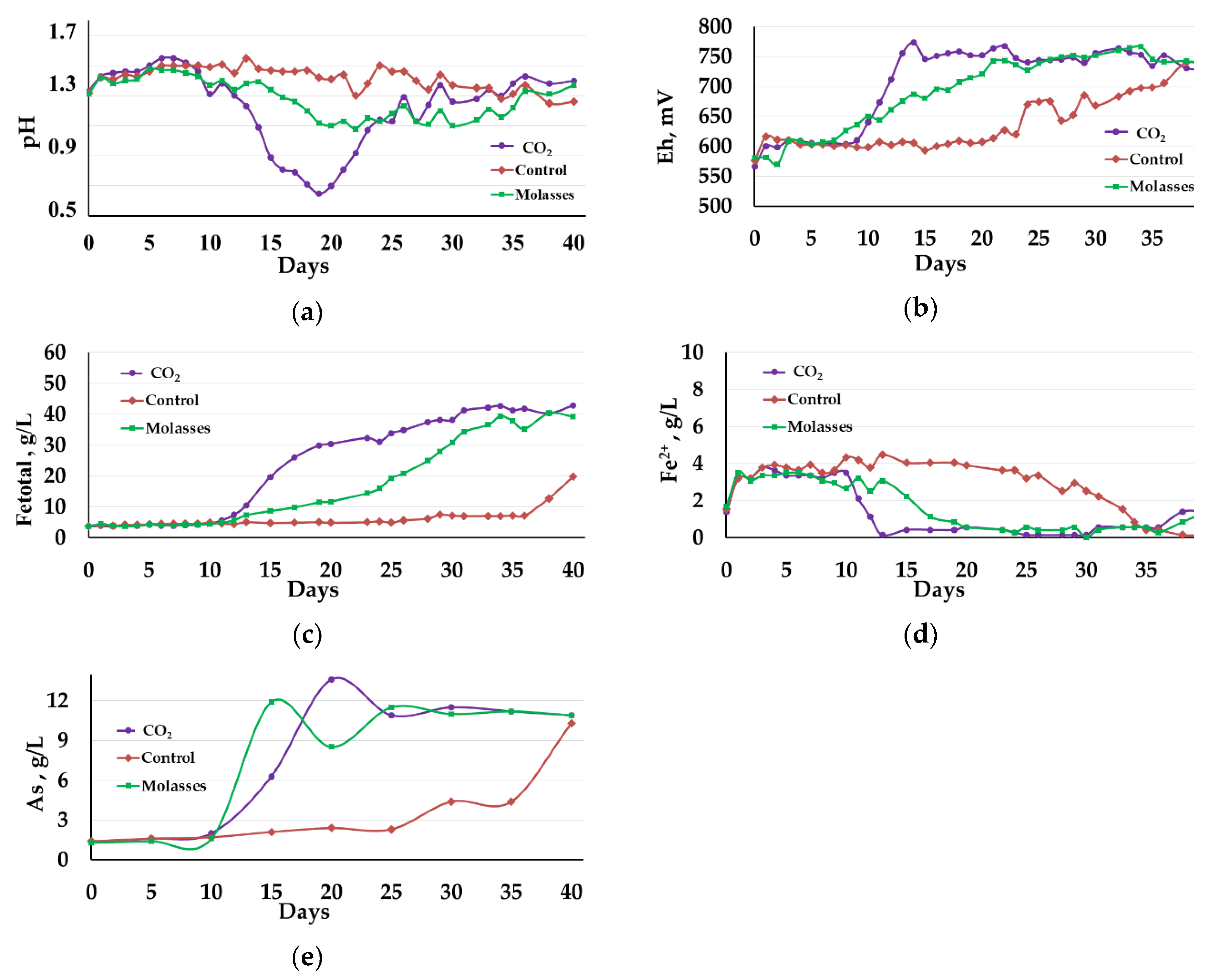
3.1.1. Liquid Phase Analysis
3.1.2. Solid Residue Analysis
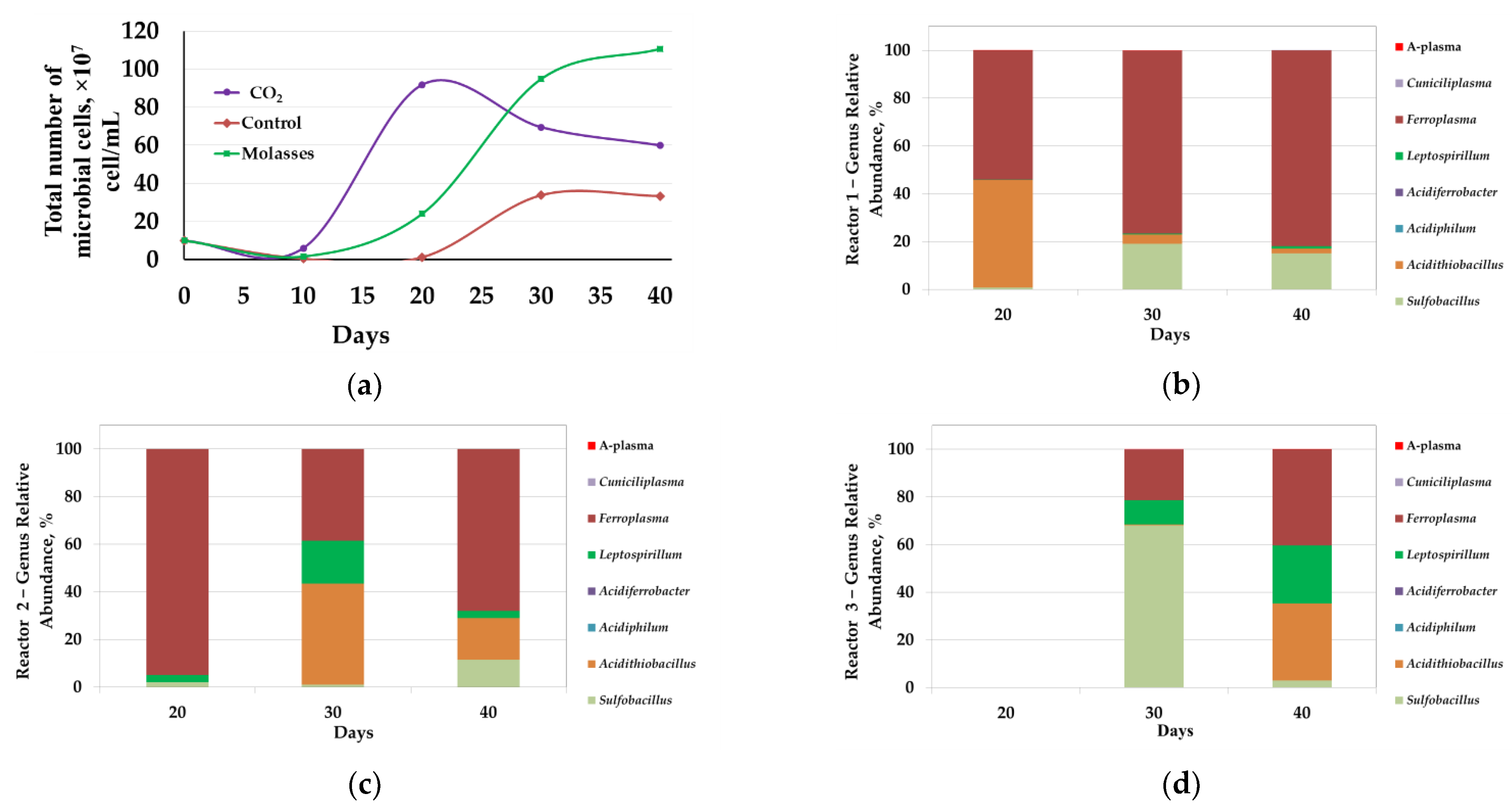
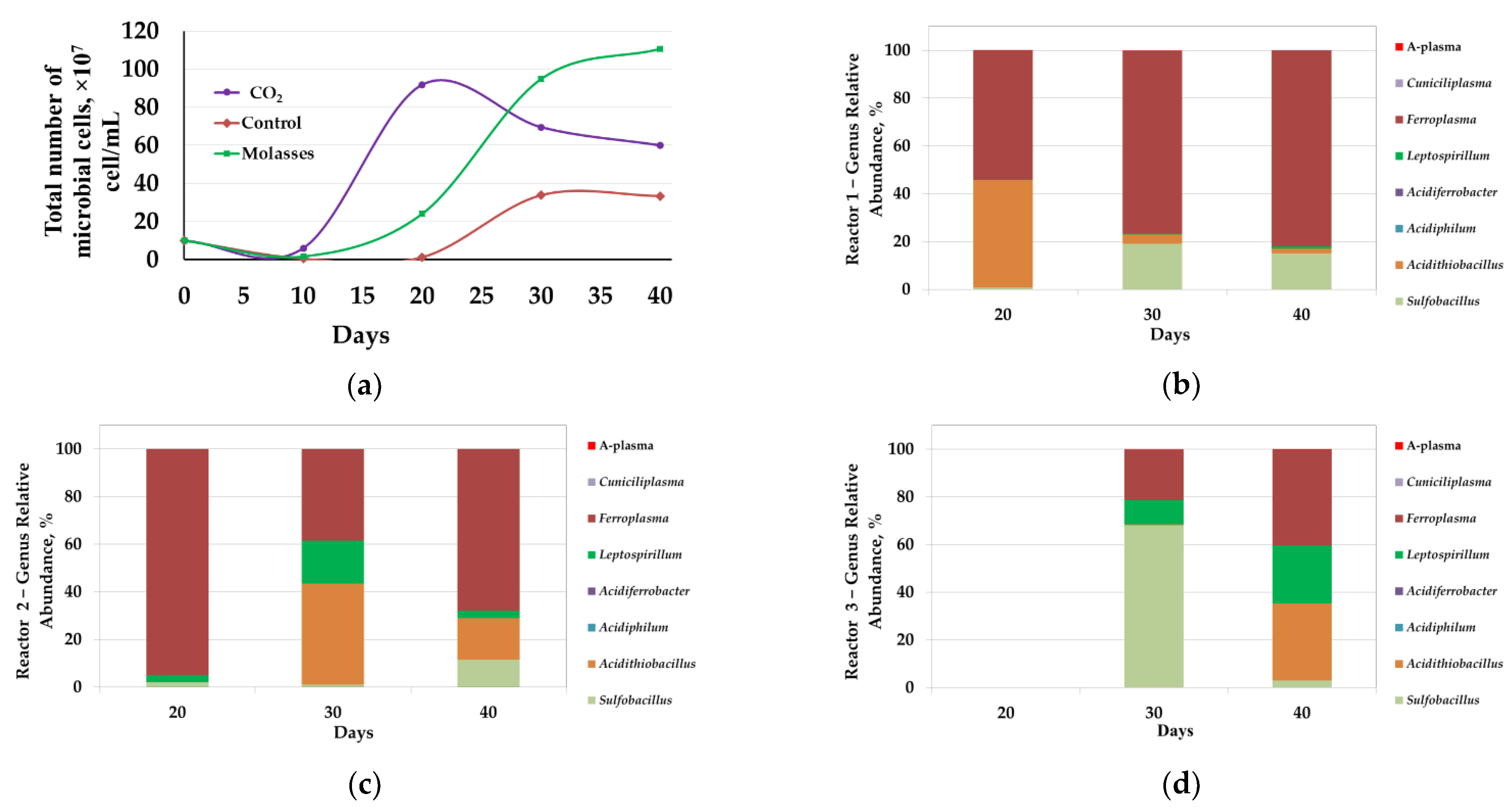
3.1.3. Microbial Population Analysis
3.2. Biooxidation under Thermophilic Conditons
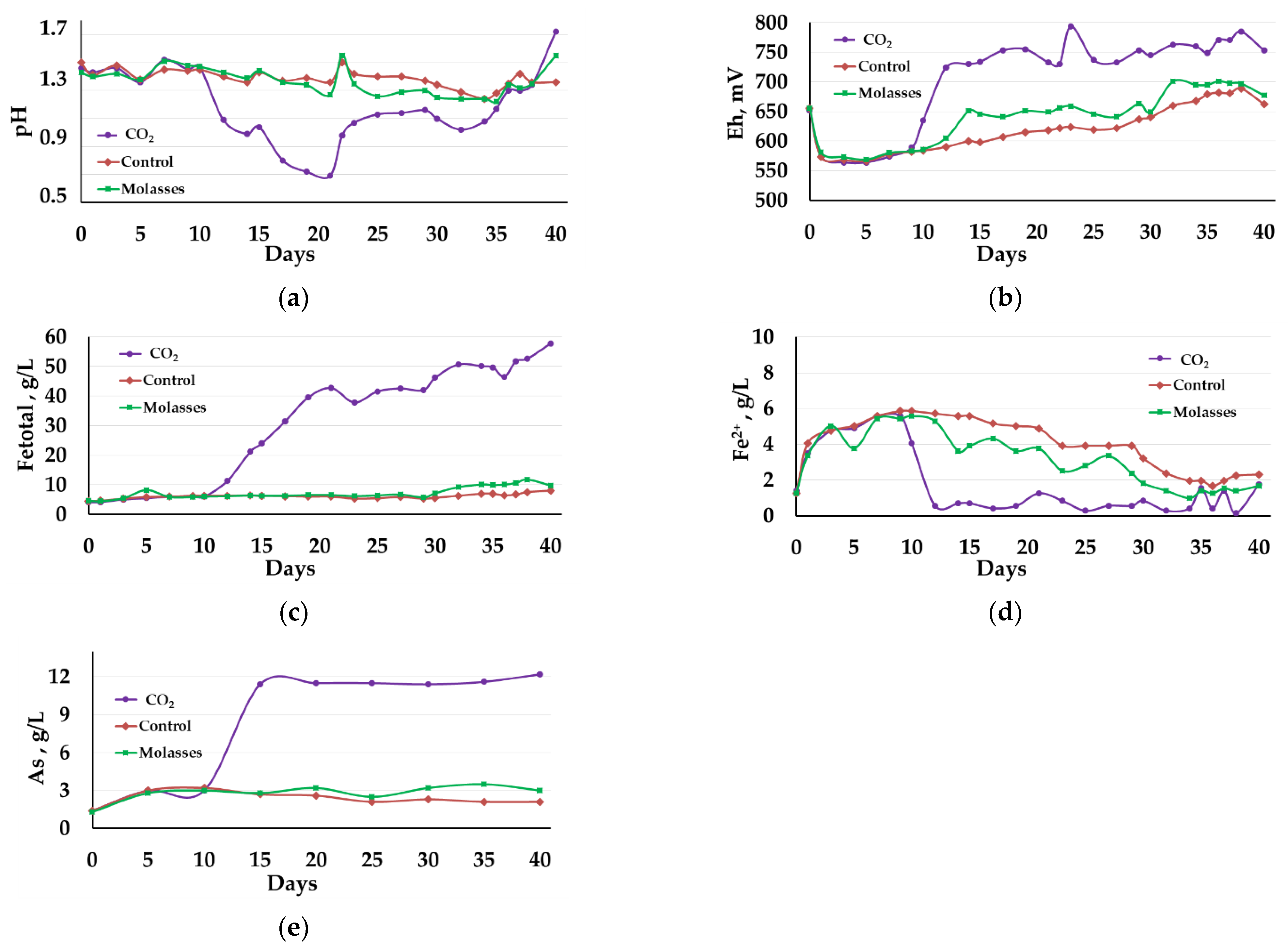
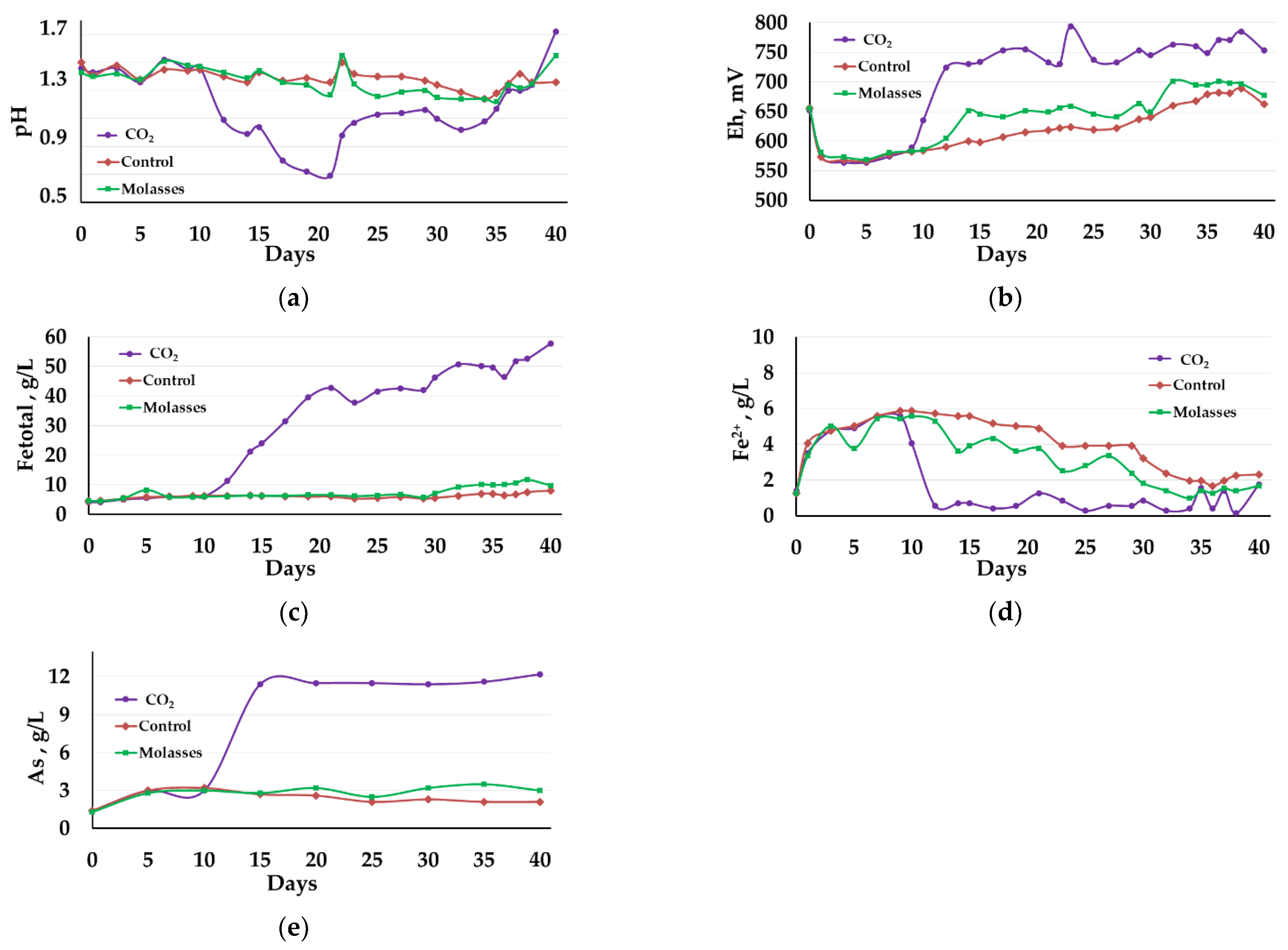
3.2.1. Liquid Phase Analysis
3.2.2. Solid Residue Analysis
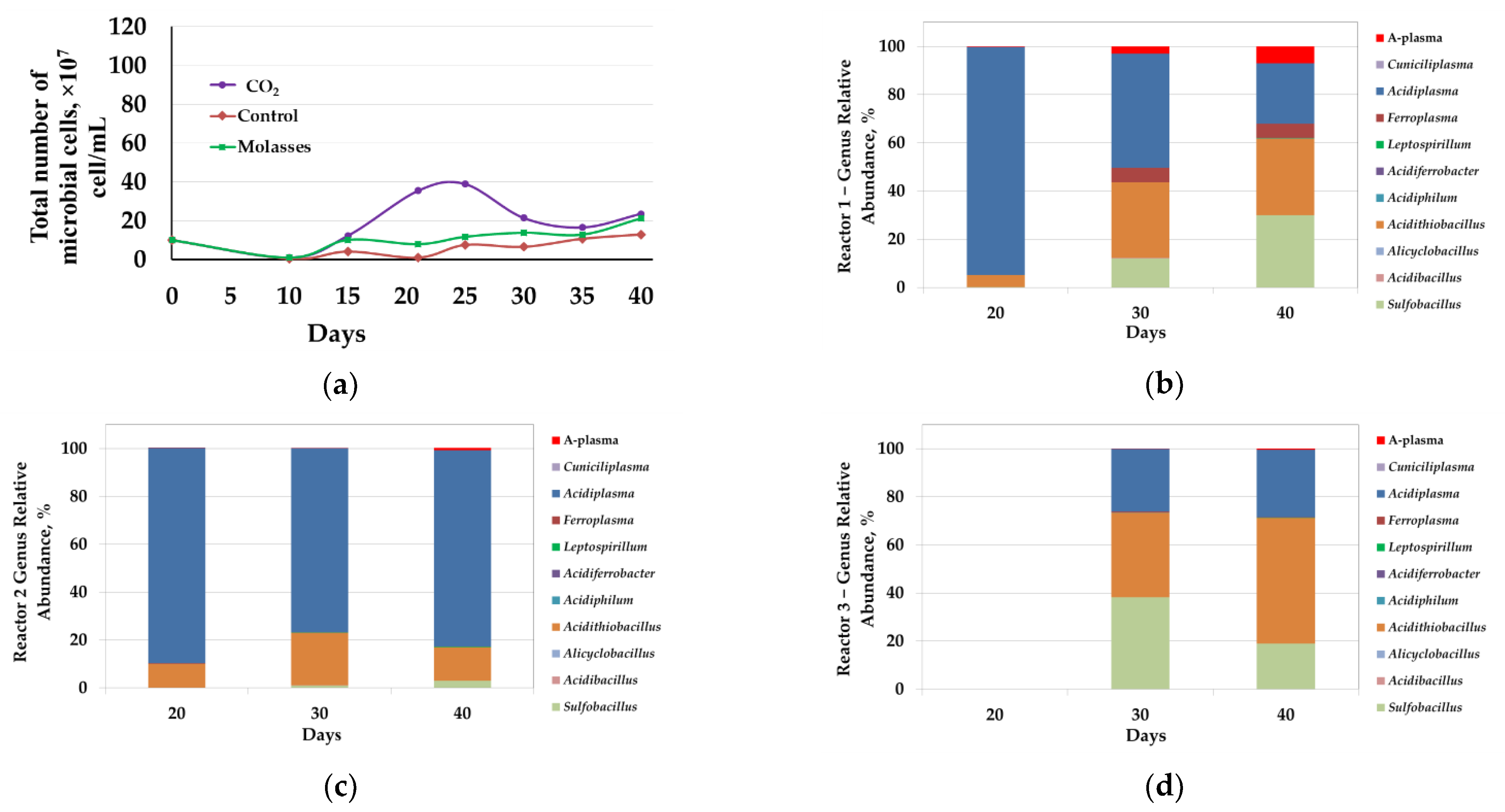
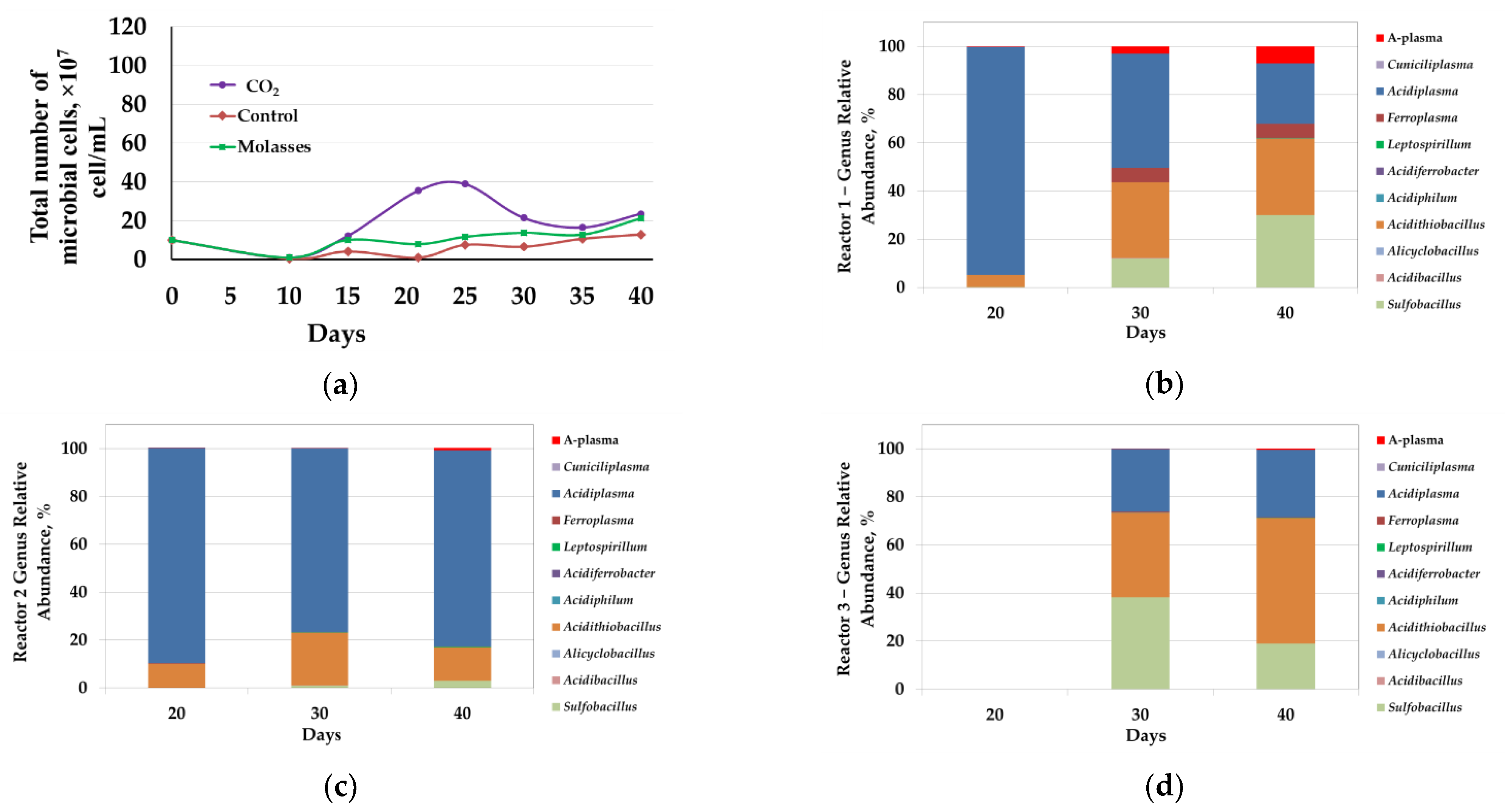
3.2.3. Microbial Population Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Johnson, D.B. Biomining—Biotechnologies for extracting and recovering metals from ores and waste materials. Curr. Opin. Biotechnol. 2014, 30, 24–31. [Google Scholar] [CrossRef]
- Johnson, D.B. The Evolution, Current Status, and Future Prospects of Using Biotechnologies in the Mineral Extraction and Metal Recovery Sectors. Minerals 2018, 8, 343. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, A.; Cezac, P.; Hoadley, A.F.A.; Contaminea, F.; D’Hugues, P. A review of sulfide minerals microbially assisted leaching in stirredtank reactors. Int. Biodeteriorat. Biodegrad. 2017, 119, 118–146. [Google Scholar] [CrossRef]
- Marsden, J.O.; House, C.I. The Chemistry of Gold Extraction, 2nd ed.; Society for Mining, Metallurgy, and Exploration, Inc.: Littleton, CO, USA, 2006; 625p. [Google Scholar]
- Thomas, K.G. Pressure oxidation overview. In Developments in Mineral Processing, Advances in Gold Ore Processing; Adams, M.D., Wills, B.A., Eds.; Elsevier B.V: Amsterdam, The Netherlands, 2006; Volume 15, pp. 346–369. [Google Scholar]
- van Aswegen, P.C.; van Niekerk, J.; Olivier, W. The BIOX process for the treatment of refractory gold concentrate. In Biomining; Rawlings, D.E., Johnson, B.D., Eds.; Springer: Berlin, Germany, 2007; pp. 1–35. [Google Scholar] [CrossRef]
- Gericke, M.; Neale, J.W.; van Staden, P.J. A Mintek perspective of the past 25 years in minerals bioleaching. J. S. Afr. Inst. Min. Metall. 2017, 109, 567–585. [Google Scholar]
- Belyi, A.V.; Chernov, D.V.; Solopova, N.V. Development of BIONORD® technology on Olimpiada deposit refractory arsenic–gold ores treatment in conditions of Extreme North. Hydrometallurgy 2018, 179, 188–191. [Google Scholar] [CrossRef]
- Rawlings, D.E.; Coram, N.J.; Gardner, M.N.; Deane, S.M. Thiobacillus caldus and Leptospirillum ferrooxidans are widely distributed in continuous flow biooxidation tanks used to treat a variety of metal containing ores and concentrates. In Biohydrometallurgy and the Environment: Toward the Mining of the 21st Century. Part A; Amils, R., Ballester, A., Eds.; Elsevier: Amsterdam, The Netherlands, 1999; pp. 777–786. [Google Scholar] [CrossRef]
- Coram, N.J.; Rawlings, D.E. Molecular relationship between two groups of the genus Leptospirillum and the finding that Leptospirillum ferriphilum sp. nov. dominates South African commercial biooxidation tanks that operate at 40 °C. Appl. Environ. Microbiol. 2002, 68, 838–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okibe, N.; Gericke, M.; Hallberg, K.B.; Johnson, D.B. Enumeration and characterization of acidophilic microorganisms isolated from a pilot plant stirred tank bioleaching operation. Appl. Environ. Microbiol 2003, 69, 1936–1943. [Google Scholar] [CrossRef] [Green Version]
- Dopson, M.; Lindstrom, E.B. Analysis of community composition during moderately thermophilic bioleaching of pyrite, arsenical pyrite, and chalcopyrite. Microb. Ecol. 2004, 48, 19–28. [Google Scholar] [CrossRef]
- Morin, D.H.R.; d’Hugues, P. Bioleaching of a cobalt containing pyrite in stirred reactors: A case study from laboratory scale to industrial application. In Biomining; Rawlings, D.E., Johnson, B.D., Eds.; Springer: Berlin, Germany, 2007; pp. 35–55. [Google Scholar] [CrossRef]
- Spolaore, P.; Joulian, C.; Gouin, J.; Ibáñez, A.; Auge, T.; Morin, D.; d’Hugues, P. Bioleaching of an organic–rich polymetallic concentrate using stirred–tank technology. Hydrometallurgy 2009, 99, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Spolaore, P.; Joulian, C.; Gouin, J.; Morin, D.; d’ Hugues, P. Relationship between bioleaching performance, bacterial community structure and mineralogy in the bioleaching of a copper concentrate in stirred–tank reactors. Appl. Microbiol. Biotechnol. 2010, 89, 441–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, W.; Qiu, G.; Zhou, H.; Peng, J.; Chen, M.; Tan, S.N.; Chao, W.; Liu, X.; Zhang, Y. Community structure and dynamics of the free and attached microorganisms during moderately thermophilic bioleaching of chalcopyrite concentrate. Bioresour. Technol. 2010, 101, 7068–7075. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Su, L.; Zhang, L.; Zeng, W.; Wu, J.; Wan, L.; Qiu, G.; Chen, X.; Zhou, H. Bioleaching of chalcopyrite by defined mixed moderately thermophilic consortium including a marine acidophilic halotolerant bacterium. Bioresour. Technol. 2012, 121, 348–354. [Google Scholar] [CrossRef]
- van Hille, R.P.; van Wyk, N.; Froneman, T.; Harrison, S.T.L. Dynamic evolution of the microbial community in BIOX leaching tanks. Adv. Mater. Res. 2013, 825, 331–334. [Google Scholar] [CrossRef]
- Kondrat’eva, T.F.; Pivovarova, T.A.; Bulaev, A.G.; Moshchanetskii, P.V.; Tsaplina, I.A.; Grigor’eva, N.V.; Zhuravleva, A.E.; Melamud, V.S.; Belyi, A.V. Selection of a community of acidochemolithotrophic microorganisms with a high oxidation rate of pyrrhotite–containing sulphide ore flotation concentrate. Appl. Biochem. Microbiol. 2013, 49, 495–501. [Google Scholar] [CrossRef]
- Muravyov, M.I.; Bulaev, A.G. Two–step oxidation of a refractory gold–bearing sulfidic concentrate and the effect of organic nutrients on its biooxidation. Miner. Eng. 2013, 45, 108–114. [Google Scholar] [CrossRef]
- Wang, Y.; Zeng, W.; Qiu, G.; Chen, X.; Zhou, H. A moderately thermophilic mixed microbial culture for bioleaching of chalcopyrite concentrate at high pulp density. Appl. Environ. Microbiol. 2014, 80, 741–750. [Google Scholar] [CrossRef] [Green Version]
- Hedrich, S.; Guézennec, A.-G.; Charron, M.; Schippers, A.; Joulian, C. Quantitative monitoring of microbial species during bioleaching of a copper concentrate. Front. Microbiol. 2016, 7, 20441. [Google Scholar] [CrossRef] [PubMed]
- Bulaev, A.; Belyi, A.; Panyushkina, A.; Solopova, N.; Pivovarova, T. Microbial population of industrial biooxidation reactors. Solid State Phenom. Submitt. 2017, 262, 48–52. [Google Scholar] [CrossRef]
- Bulaev, A.; Melamud, V.; Boduen, A. Bioleaching of non-ferrous metals from arsenic-bearing sulfide concentrate. Solid State Phenom. 2020, 299, 1064–1068. [Google Scholar] [CrossRef]
- Karavaiko, G.I.; Dubinina, G.A.; Kondrateva, T.F. Lithotrophic microorganisms of the oxidative cycles of sulfur and iron. Microbiology (Mikrobiologiya) 2006, 75, 512–545. [Google Scholar] [CrossRef]
- Muravyov, M.I.; Bulaev, A.G. A two-step process for the treatment of refractory sulphidic concentrate. Adv. Mater. Res. 2013, 825, 246–249. [Google Scholar] [CrossRef]
- Schippers, A. Microorganisms involved in bioleaching and nucleic acid-based molecular methods for their identification and quantification. In Microbial Processing of Metal Sulfides; Donati, E.R., Sand, W., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 3–33. [Google Scholar] [CrossRef]
- Gonzalez-Tori, E.; Llobet-Brossa, E.; Casamayor, E.O.; Amann, R.; Amils, R. Microbial ecology of an extreme acidic environment, the Tinto River. Appl. Environ. Microbiol. 2003, 69, 4853–4865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Druschel, G.K.; Baker, B.J.; Gihring, T.H.; Banfield, J.F. Acid mine drainage biogeochemistry at Iron Mountain. Calif. Geochem. Trans. 2004, 5, 13–32. [Google Scholar] [CrossRef] [Green Version]
- Schnaitman, C.; Lundgren, D.G. Organic compounds in the spent medium of Ferrobacillus ferrooxidans. Can. J. Microbiol 1965, 11, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Borichewski, R.M. Keto acids as growth-limiting factors in autotrophic growth of Thiobacillus thiooxidans. J. Bacteriol 1967, 93, 597–599. [Google Scholar] [CrossRef] [Green Version]
- Okibe, N.; Johnson, D.B. Biooxidation of pyrite by defined mixed cultures of moderately thermophilic acidophiles in pH-controlled bioreactors: Significance of microbial interactions. Biotechnol. Bioeng. 2004, 87, 574–583. [Google Scholar] [CrossRef]
- Nancucheo, I.; Johnson, D.B. Production of glycolic acid by chemolithotrophic iron- and sulfur-oxidizing bacteria and its role in delineating and sustaining acidophilic sulfide mineral-oxidizing consortia. Appl. Environ. Microbiol. 2010, 76, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Bulaev, A.G. Biooxidation of refractory pyrite-arsenopyrite gold bearing sulfide concentrate. In Proceedings of the19th International Multidisciplinary Scientific GeoConference (SGEM 2019), Albena, Bulgaria, 28 June–7 July 2019; Volume 6.3, pp. 67–74. [Google Scholar] [CrossRef]
- Reznikov, A.A.; Mulikovskaya, E.P.; Sokolov, I.Y. Metody analiza prirodnykh vod (Methods for Analysis of Natural Waters); Nedra: Moscow, Russia, 1970; 140p. (In Russian) [Google Scholar]
- Surovskaya, I.A.; Titov, V.I.; Brodskaya, V.M.; Vasil’ev, P.I.; Lipshits, B.M.; Elentukh, B.M. Tekhnicheskii analiz tsvetnoi metallurgii (Technical Analysis in Nonferrous Metallurgy); Metallurgizdat: Moscow, Russia, 1957; p. 567. (In Russian) [Google Scholar]
- Filippova, N.A. Fazovyi analiz rud i produktov ikh pererabotki (Phase Analysis of Ores and Products of Their Processing); Khimiya: Moscow, Russia, 1975; 280p. (In Russian) [Google Scholar]
- Pimenov, N.V.; Merkel, A.Y.; Samylina, O.S.; Kanapatskii, T.A.; Tikhonova, E.N.; Vlasova, M.A.; Tarnovetskii, I.Y.; Malakhova, T.V. Structure of Microbial Mats in the Mramornaya Bay (Crimea) Coastal Areas. Microbiology (Mikrobiologiya) 2018, 87, 681–691. [Google Scholar] [CrossRef]
- Das, B. Theoretical study of formation of secondary arsenic minerals: Scorodite and pharmacosiderite. ACS Earth Space Chem. 2019, 3, 192–201. [Google Scholar] [CrossRef]
- Deng, S.; Gu, G.; He, G.; Li, L. Catalytic effect of pyrite on the leaching of arsenopyrite in sulfuric acid and acid culture medium. Electrochim. Acta 2018, 263, 8–16. [Google Scholar] [CrossRef]
- Xu, J.-N.; Shi, W.-G.; Ma, P.-C.; Lu, L.-S.; Chen, G.-M.; Yang, H.-Y. Corrosion behavior of a pyrite and arsenopyrite galvanic pair in the presence of sulfuric acid, ferric ions and HQ0211 bacterial strain. Minerals 2019, 9, 169. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, D.E.; Tributsch, H.; Hansford, G. Reasons why ‘Leptospirillum’-like species rather than Thiobacillus ferrooxidans are the dominant iron-oxidizing bacteria in many commercial processes for the biooxidation of pyrite and related ores. Microbiology 1999, 145, 5–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, R.; Gentina, J.C.; Acevedo, F. Attachment behaviour of Thiobacillus ferrooxidans cells to refractory gold concentrate particles. Biotechnol. Lett. 1999, 21, 715–718. [Google Scholar] [CrossRef]
- Lizama, H.M.; Fairweather, M.J.; Dai, Z.; Allegretto, T.D. How does bioleaching start? Hydrometallurgy 2003, 69, 109–1161. [Google Scholar] [CrossRef]
- Deveci, H. Effect of particle size and shape of solids on the viability of acidophilic bacteria during mixing in stirred tank reactors. Hydrometallurgy 2004, 71, 385–396. [Google Scholar] [CrossRef]
- Melamud, V.S.; Pivovarova, T.A. Specific features of the growth of the type strain of Sulfobacillus thermosulfidooxidans in medium 9K. Appl. Biochem. Microbiol. 1998, 34, 314–315. [Google Scholar]
- Vaughan, D.J.; Corkhill, C.L. Mineralogy of Sulfides. Elements 2017, 13, 81–87. [Google Scholar] [CrossRef]
- Bulaev, A.G. Effect of organic carbon source on pyrite biooxidation by moderately thermophilic acidophilic microorganisms. Microbiology (Moscow) 2020, 89, 301–308. [Google Scholar] [CrossRef]
- Golyshina, O.V.; Yakimov, M.M.; Lünsdorf, H.; Ferrer, M.; Nimtz, M.; Timmis, K.N.; Wray, V.; Tindall, B.J.; Golyshin, P.N. Acidiplasma aeolicum gen. nov., sp. nov., a euryarchaeon of the family Ferroplasmaceae isolated from a hydrothermal pool, and transfer of Ferroplasma cupricumulans to Acidiplasma cupricumulans comb. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 2815–2824. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Content, % |
|---|---|
| Fetot | 31.8 |
| Fes | 29.1 |
| Stot | 34.7 |
| Ss | 34.4 |
| Ssulfate | 0.2 |
| S0 | 0.1 |
| Astot | 6.9 |
| Ass | 6.5 |
| Au, g/t | 45.5 |
| Temperature | Carbon Source | pH | Eh | Fe3+, g/L | Fe2+, g/L | As, g/L | Microbial Cell Number, × 107 Cell/mL |
|---|---|---|---|---|---|---|---|
| 40 °C | CO2 | 1.40 | 730 | 41.4 | 1.4 | 10.9 | 60 |
| Molasses | 1.37 | 736 | 37.8 | 1.4 | 10.9 | 110 | |
| Control | 1.26 | 730 | 19.6 | 0.1 | 10.3 | 33 | |
| 50 °C | CO2 | 1.72 | 753 | 56.0 | 1.8 | 12.0 | 24 |
| Molasses | 1.55 | 667 | 8.1 | 1.7 | 3.0 | 22 | |
| Control | 1.36 | 662 | 5.7 | 2.3 | 2.1 | 12 |
| Mineral | Content, % | |||||
|---|---|---|---|---|---|---|
| 40 °C | 50 °C | |||||
| Reactor 1 | Reactor 2 | Reactor 3 | Reactor 1 | Reactor 2 | Reactor 3 | |
| Fetot | 7.20 | 8.50 | 24.50 | 3.90 | 24.50 | 26.10 |
| Fes | 8.62 | 16.28 | 27.00 | 1.60 | 20.37 | 22.57 |
| Stot | 20.10 | 19.90 | 31.40 | 16.20 | 28.50 | 29.90 |
| Ss | 7.60 | 8.80 | 27.40 | 2.30 | 24.50 | 26.50 |
| Ssulfate | 12.10 | 10.70 | 3.60 | 13.70 | 3.50 | 2.90 |
| S0 | 0.40 | 0.40 | 0.40 | 0.10 | 0.50 | 0.50 |
| Astot | 0.21 | 0.26 | 1.20 | 0.85 | 5.30 | 5.85 |
| Ass | 0.14 | 0.16 | 0.57 | 0.03 | 0.41 | 0.47 |
| Temperature | Carbon Source | Yield, % | Oxidation Rate, % | ||
|---|---|---|---|---|---|
| SS | Pyrite | Arsenopyrite | |||
| 40 °C | CO2 | 85.3 | 81 | 77 | 98 |
| Molasses | 85.9 | 78 | 73 | 98 | |
| Control | 78.7 | 37 | 27 | 93 | |
| 50 °C | CO2 | 73.7 | 95 | 94 | 99 |
| Molasses | 98.7 | 29 | 21 | 94 | |
| Control | 103.9 | 19 | 10 | 92 | |
| Mineral | Content, % | ||||||
|---|---|---|---|---|---|---|---|
| Concentrate | 40 °C | 50 °C | |||||
| Reactor 1 | Reactor 2 | Reactor 3 | Reactor 1 | Reactor 2 | Reactor 3 | ||
| Pyrite (FeS2) | 48 | 10 | 10 | 30 | <1 | 30 | 35 |
| Arsenopyrite (FeAsS) | 9 | <1 | <1 | <1 | <1 | <1 | <1 |
| Quartz (SiO2) | 25 | 20 | 15 | 20 | 30 | 35 | 35 |
| Mica | 18 | 10 | 10 | 5 | 15 | 25 | 20 |
| Gypsum (CaSO4·2H2O) | 0 | 55 | 65 | 45 | 15 | 0 | 0 |
| Bassanite (CaSO4·0.5H2O) | 0 | 0 | 0 | 0 | 40 | 10 | 10 |
| Anhydrite (CaSO4) | 0 | 5 | 0 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bulaev, A.; Nechaeva, A.; Elkina, Y.; Melamud, V. Effect of Carbon Sources on Pyrite-Arsenopyrite Concentrate Bio-oxidation and Growth of Microbial Population in Stirred Tank Reactors. Microorganisms 2021, 9, 2350. https://doi.org/10.3390/microorganisms9112350
Bulaev A, Nechaeva A, Elkina Y, Melamud V. Effect of Carbon Sources on Pyrite-Arsenopyrite Concentrate Bio-oxidation and Growth of Microbial Population in Stirred Tank Reactors. Microorganisms. 2021; 9(11):2350. https://doi.org/10.3390/microorganisms9112350
Chicago/Turabian StyleBulaev, Aleksandr, Aleksandra Nechaeva, Yuliya Elkina, and Vitaliy Melamud. 2021. "Effect of Carbon Sources on Pyrite-Arsenopyrite Concentrate Bio-oxidation and Growth of Microbial Population in Stirred Tank Reactors" Microorganisms 9, no. 11: 2350. https://doi.org/10.3390/microorganisms9112350
APA StyleBulaev, A., Nechaeva, A., Elkina, Y., & Melamud, V. (2021). Effect of Carbon Sources on Pyrite-Arsenopyrite Concentrate Bio-oxidation and Growth of Microbial Population in Stirred Tank Reactors. Microorganisms, 9(11), 2350. https://doi.org/10.3390/microorganisms9112350