
Malaria Diagnosis in Non-Endemic Settings: The European Experience in the Last 22 Years

 ,
, 


 , and
, and
Abstract
1. Introduction
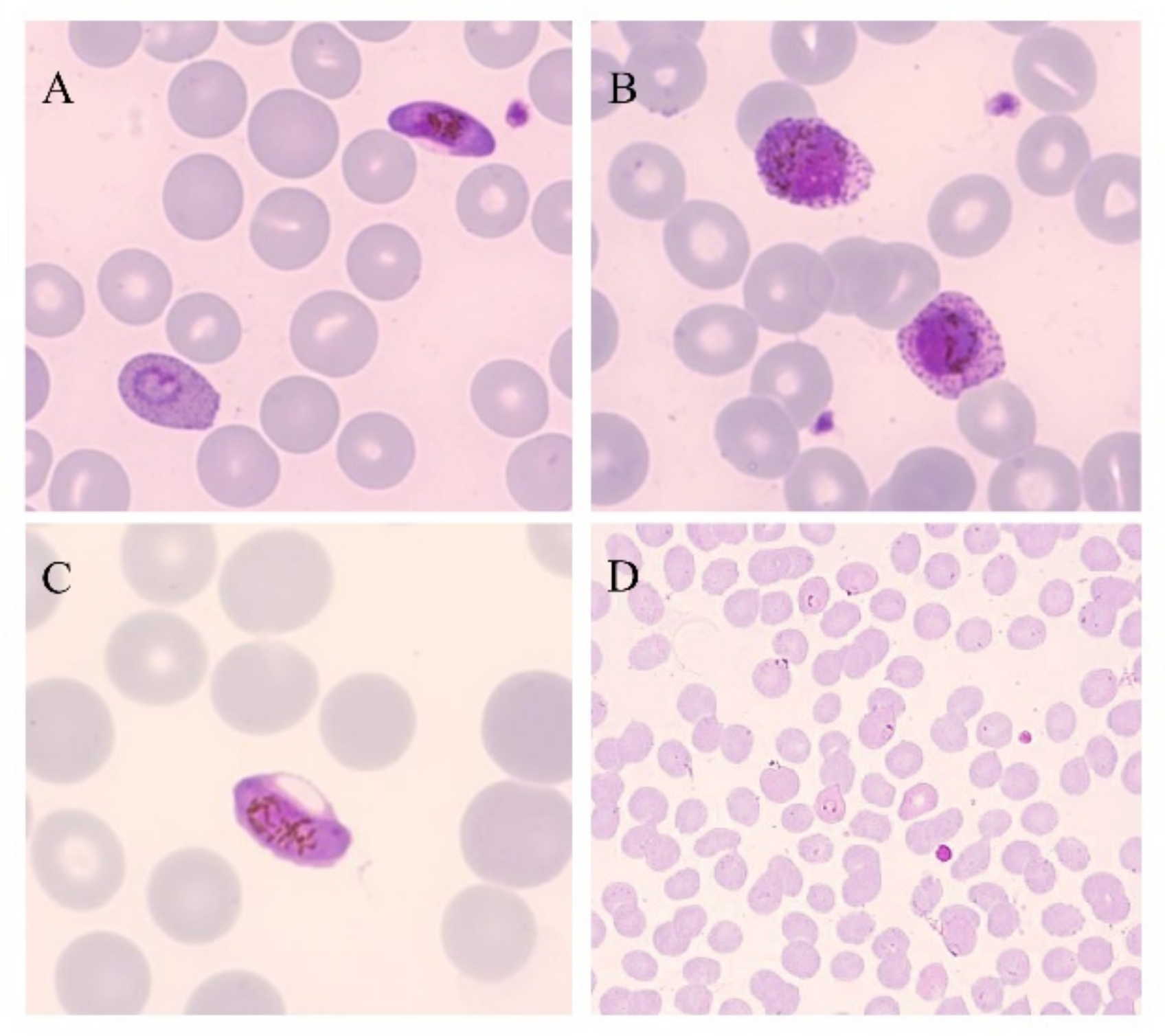
2. Gold-Standard Method
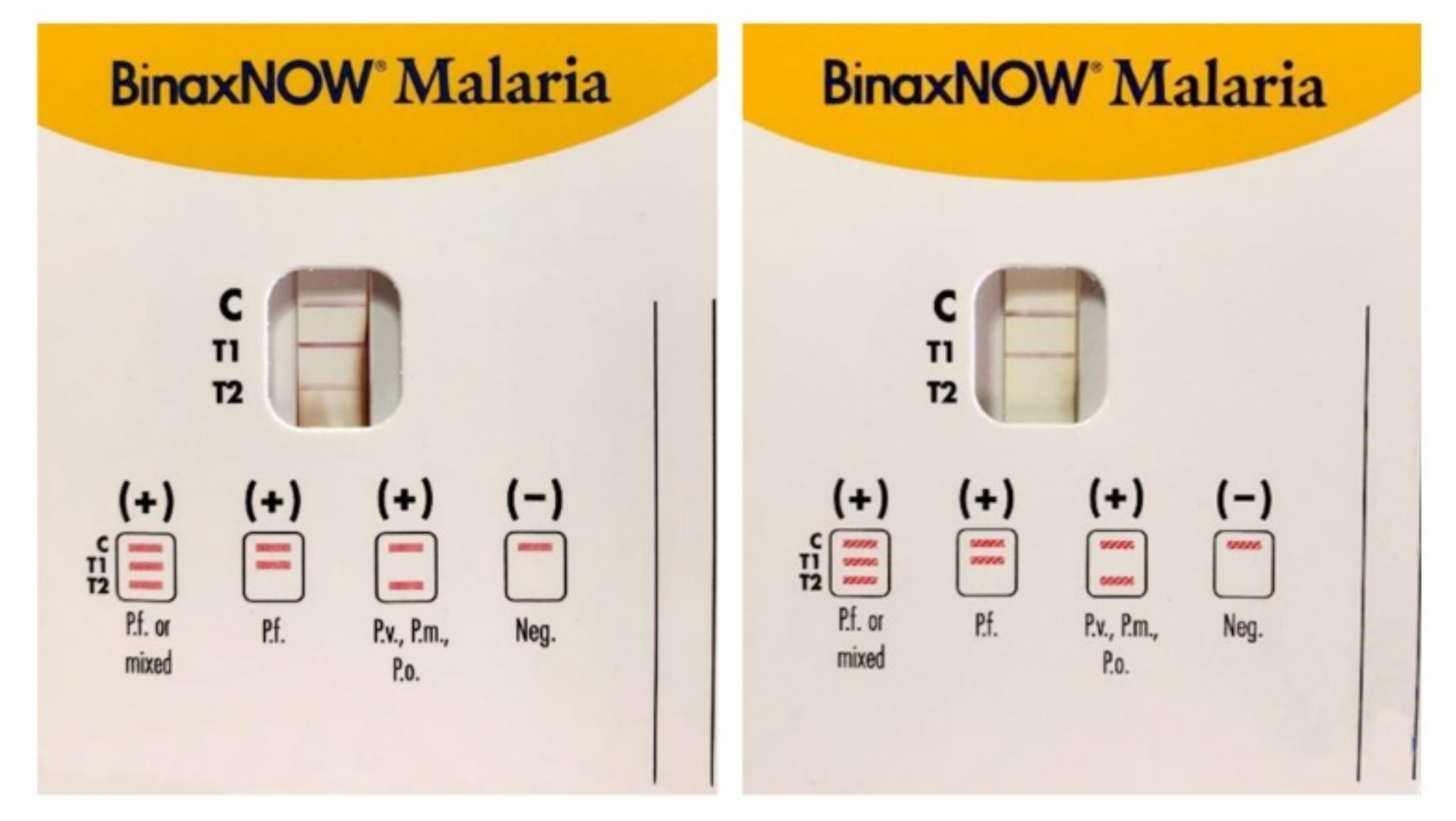
3. Rapid Diagnostic Tests
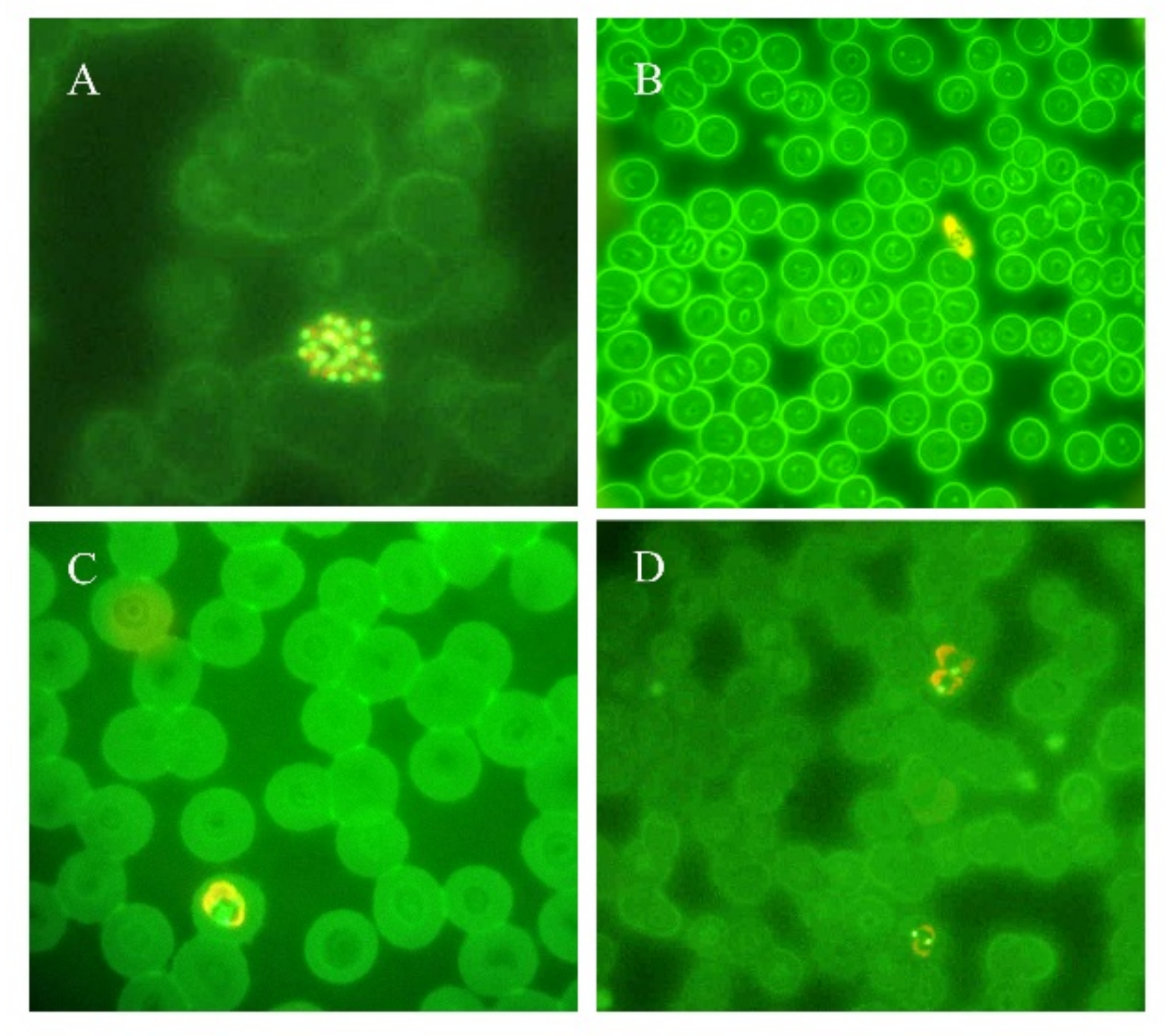
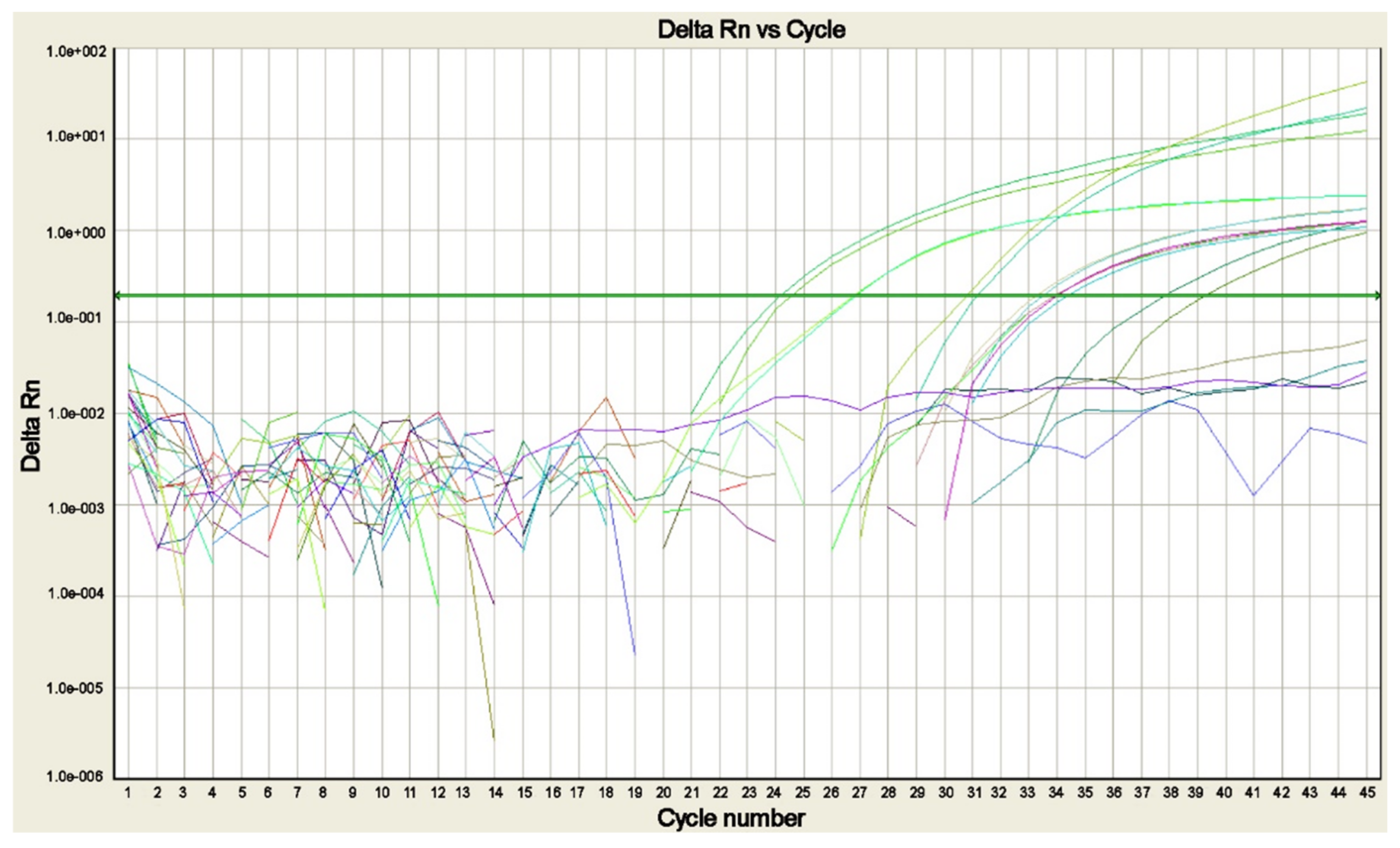
4. Molecular Assays

{kind=link}
{kind=link}
{kind=link}
{kind=link}
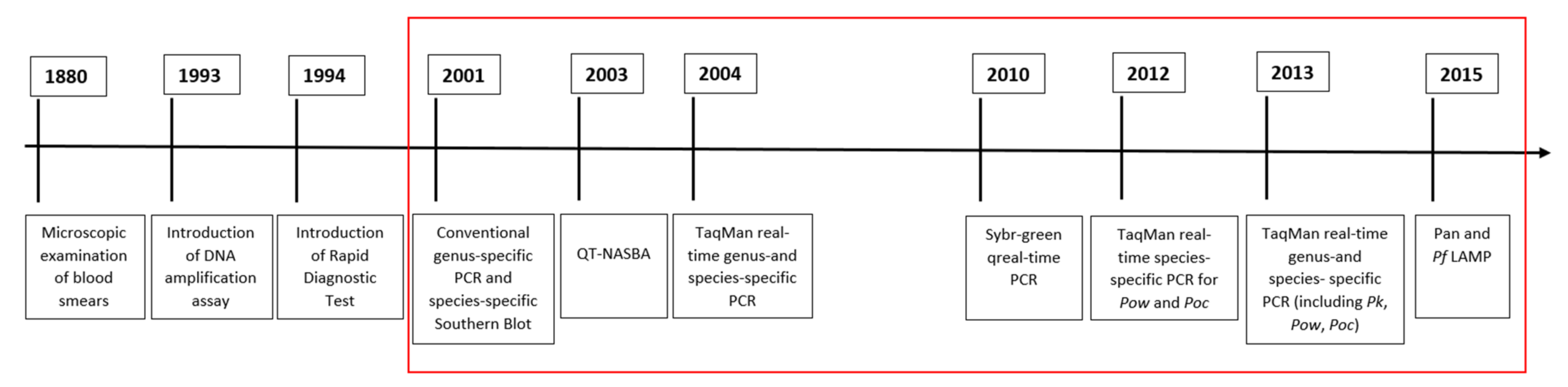{kind=link}
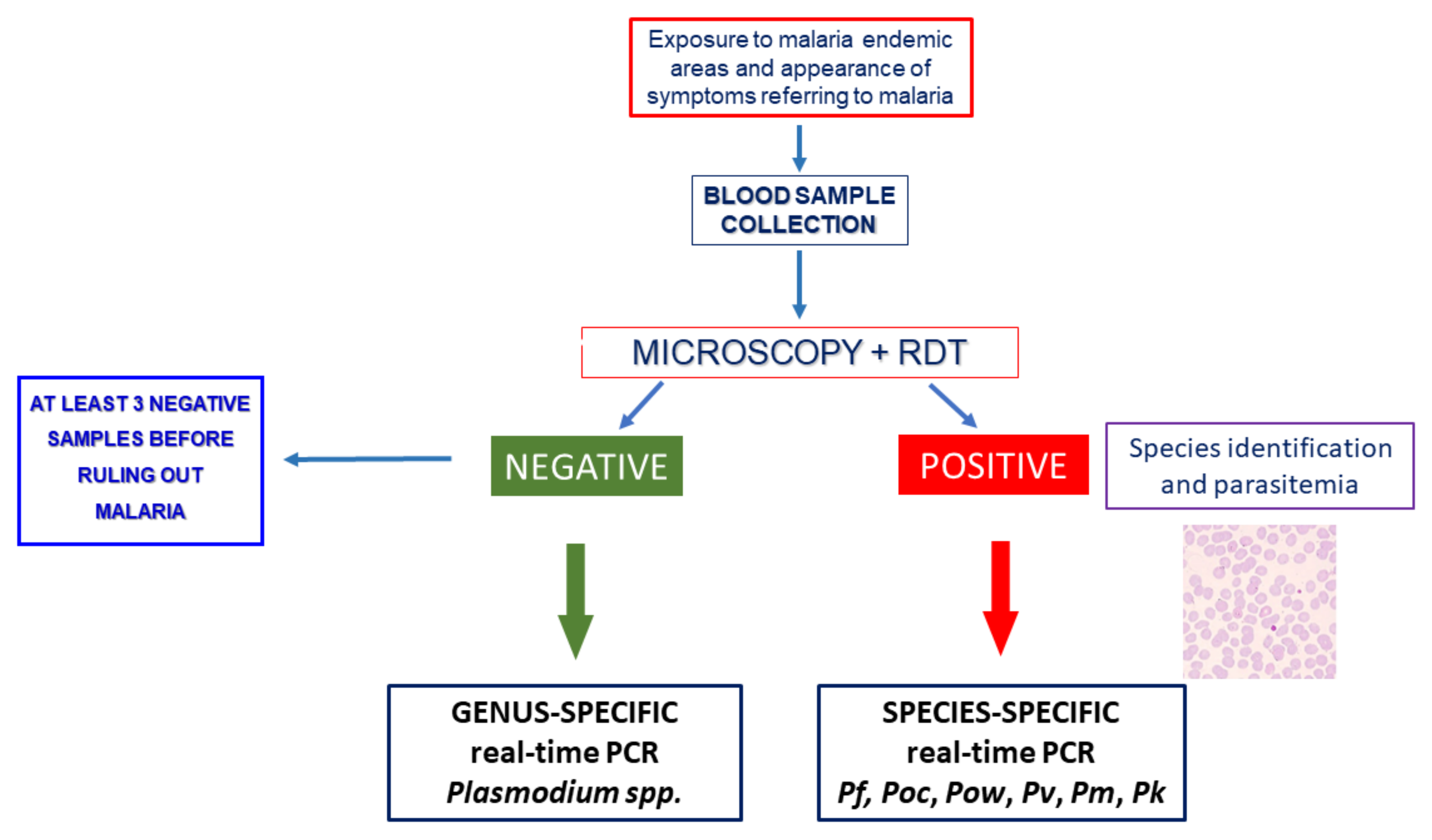{kind=link}
| Evaluated Assay | Type of Amplification | Target | Country | Samples/ Patients Tested | Period | Reference Test | Performance/ Agreement with Reference Test | Reference |
|---|---|---|---|---|---|---|---|---|
| In-house species-specific PCR | Nested PCR | 18S rRNA | Spain | 192 samples/patients with suspected malaria | 1997–1998 | Microscopy | 12.4% more malaria cases detected by PCR | [37] |
| In-house genus-specific PCR | Conventional genus-specific PCR | 18S rRNA | Italy | 101 samples/ patients with suspected malaria | 1994–1999 | Microscopy | Sensitivity 100% Specificity 100% | [49] |
| species-specific PCR by [67] and by [68] | species-specific Southern blot | Agreement 94% | ||||||
| In-house species-specific PCR by [58] | Nested PCR | 18S rRNA | Poland | 216 patients with suspected malaria | / | Microscopy | Agreement 83.8% | [50] |
| In-house genus-specific PCR | QT-NASBA | 18S rRNA | The Netherlands | 113 patients with suspected malaria | 4 months | Microscopy | Sensitivity 100% Specificity 94% Agreement 94.7% | [51] |
| In-house genus-specific qPCR | TaqMan | 18S rRNA | Switzerland | 97 samples/ 66 from patients with suspected malaria + 31 from patients with known Pf malaria | 2002–2003 | Microscopy | 86% agreement | [52] |
| In-house species-specific qPCR | 71% agreement | |||||||
| In-house genus-specific PCR+ species-specific PCR (Pf, Pv, Po) | TaqMan | 18S rRNA | Italy | 122 samples/patients with suspected malaria | / | 18S rRNA nested PCR | Sensitivity 100% Specificity 100% | [53] |
| In-house species-specific PCR | QT-NASBA | 18S rRNA | The Netherlands | 79 samples of patients with malaria | / | Microscopy | Perfect agreement (evaluated by Cohen’s Kappa coefficient) | [54] |
| Pf qReal Time-PCR by [69] | Sybr Green | Pf CoxI gene | France | 192 patients with suspected malaria | 2005–2007 | Microscopy +RDT | 93.3% agreement | [55] |
| In-house genus-specific qPCR | Plasmodium mitochondrial sequence | 99% agreement | ||||||
| In-house species-specific qPCR | 18S rRNA | 98% agreement | ||||||
| Genus-specific qPCR by [52] with modifications | TaqMan | 18S rRNA Plasmodium gene | Belgium | 351 samples | 1995–2009 | Microscopy | 8.3% cases detected only by PCR | [56] |
| In-house species-specific PCR | 18S rRNA specific genes | 1.3% species identification only by PCR | ||||||
| In-house species-specific PCR by [70] | Seminested PCR | 18S rRNA | Italy | 1226 patients with suspected malaria | 1998–2003 | Microscopy | Sensitivity 100% Specificity 100% PPV 100% NPV 100% | [40] |
| In-house species-specific PCR for Poc and Pow | TaqMan | 18S rRNA | Italy | 31 samples from patients with P.ovale malaria | / | 18S rRNA nested PCR | 100% agreement | [39] |
| Genus-specific PCR by [58] | TaqMan | 18S rRNA | Norway | 135 samples/patients with suspected malaria | 2006–2011 | Nested SSU rRNA PCR | 93% agreement | [42] |
| Genus-specific PCR by [71] | Mitochondrial DNA sequence | 97% agreement | ||||||
| Species-specific PCR [72] | 18S rRNA | 87% agreement | ||||||
| In-house genus-specific PCR + species -specific PCR | TaqMan | 18S rRNA (including Pk, Poc and Pow) | Italy | 398 samples/ patients with suspected malaria | 2000–2012 | Microscopy | 6.3% species identification only by PCR:1.5% samples disagreement with microscopy | [41] |
| Genus-specific qPCR [52] | TaqMan | 18S rRNA Plasmodium gene | Serbia | 109 samples/patients with suspected malaria | 2010–2013 | Microscopy | 95.5% agreement | [38] |
| Species-specific qPCR by [53] (for Pf, Pv, Po) and by [52] (for Pm) | 18S rRNA specific genes | 73.3% agreement | ||||||
| Pan and Pf LoopAMP® (Eiken Chemical Co.) | LAMP | Mitochondrial DNA sequence | Switzerland | 210 samples/patients with suspected malaria | March–October 2012 | Microscopy | Sensitivity 100% Specificity 97.5% PPV 91.5% NPV 100% | [43] |
| 18S rRNA qPCR | Sensitivity 100% Specificity 100% PPV 100% NPV 100% | |||||||
| Pan and Pf LoopAMP® (Eiken Chemical Co.) | LAMP | Mitochondrial DNA sequence | Spain | 427 samples: 29 Po positive samples+ 398 negative samples stored | 2014–2016 | Nested SSU rRNA PCR | Sensitivity 100% Specificity 97.2% PPV 72.5% NPV 100% | [44] |
| Genus-specific PCR and Species-specific PCR (FTD Malaria, Fast-Track Diagnostics®) | TaqMan | / | Spain | 250 patients: 86 with suspected malaria+ 164 asymptomatic immigrants from endemic areas | 2015–2017 | In-house genus-and species-specific PCR | Sensitivity 96% Specificity 97.4% PPV 93.6% NPV 98% | [45] |
| Species-specific PCR by RealStar Malaria S&T PCR Kit 1.0 | TaqMan | Plasmodium spp. DNA (including Pk) | Germany | 179 samples positive by microscopy and genus-specific PCR | April–December 2017 | Microscopy+ genus-specific PCR | Sensitivity 95.1% | [46] |
| Genus-specific PCR and Species-specific PCR (FTD Malaria, Fast-Track Diagnostics®) | Sensitivity 96.8% | |||||||
| In-house species-specific duplex PCR for Poc and Pow by [73] | TaqMan | 18S rRNA | Germany | 77 samples/patients with P.ovale malaria | 2010–2019 | / | 100% agreement among the 2 evaluated assays | [63] |
| In-house species-specific singleplex PCR for Poc and Pow described by [74] and [39] | ||||||||
| Alethia assay (Meridian Bioscience) | LAMP | Undeclared target: segments of the Plasmodium genome | France | 331 samples/patients with suspected malaria | 2017–2018 | Real-time PCR | Sensitivity 97.3% Specificity 99.6% PPV 94.8% NPV 99.8% | [47] |
| In-house Pfhrp2 PCR | TaqMan | Pfhrp2 gene | United Kingdom/ Switzerland/ Portugal | 50 DNA samples from suspected Pf patients from Eritrea | / | Conventional qPCR | Sensitivity 100% | [48] |
| In-house Pfhrp3 PCR | Pfhrp3 gene | Specificity 100% |
5. Other Diagnostic Methods
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ashley, E.A.; Pyae Phyo, A.; Woodrow, C.J. Malaria. Lancet 2018, 391, 1608–1621. [Google Scholar] [CrossRef]
- Anstey, N.M.; Grigg, M.J. Zoonotic Malaria: The Better You Look, the More You Find. J. Infect. Dis. 2019, 219, 679–681. [Google Scholar] [CrossRef] [PubMed]
- Imwong, M.; Madmanee, W.; Suwannasin, K.; Kunasol, C.; Peto, T.J.; Tripura, R.; von Seidlein, L.; Nguon, C.; Davoeung, C.; Day, N.P.J.; et al. Asymptomatic Natural Human Infections with the Simian Malaria Parasites Plasmodium cynomolgi and Plasmodium knowlesi. J. Infect. Dis. 2019, 219, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Grigg, M.J.; Snounou, G. Plasmodium simium: A Brazilian focus of anthropozoonotic vivax malaria? Lancet Glob. Health 2017, 5, e961–e962. [Google Scholar] [CrossRef]
- WHO. World Malaria Report 2020. Available online: https://www.who.int/publications/i/item/9789240015791 (accessed on 28 September 2021).
- WHO. Severe malaria. Trop. Med. Int. Health 2014, 19 (Suppl. 1), 7–131. [Google Scholar] [CrossRef] [PubMed]
- Kamaliddin, C.; Le Bouar, M.; Berry, A.; Fenneteau, O.; Gillet, P.; Godineau, N.; Candolfi, E.; Houzé, S. Assessment of diagnostic methods for imported malaria in mainland France. Med. Mal. Infect. 2020, 50, 141–160. [Google Scholar] [CrossRef] [PubMed]
- WHO. Global Technical Strategy for Malaria 2016–2030. 2021. Available online: https://www.who.int/publications/i/item/9789240031357 (accessed on 28 September 2021).
- WHO. WHO Malaria Terminology. 2019. Available online: https://www.who.int/publications/i/item/WHO-HTM-GMP-2016.6 (accessed on 29 August 2021).
- Centers for Disease Control and Prevention. Malaria’s Impact Worldwide. 2021. Available online: https://www.cdc.gov/malaria/malaria_worldwide/impact.html (accessed on 28 September 2021).
- Piperaki, E.T.; Daikos, G.L. Malaria in Europe: Emerging threat or minor nuisance? Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2016, 22, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Tseroni, M.; Pervanidou, D.; Tserkezou, P.; Rachiotis, G.; Pinaka, O.; Baka, A.; Georgakopoulou, T.; Vakali, A.; Dionysopoulou, M.; Terzaki, I.; et al. Field application of SD bioline malaria Ag Pf/Pan rapid diagnostic test for malaria in Greece. PLoS ONE 2015, 10, e0120367. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. 2021 Malaria-Annual Epidemiological Report for 2019. Available online: https://www.ecdc.europa.eu/en/publications-data/malaria-annual-epidemiological-report-2019#no-link (accessed on 28 September 2021).
- Calderaro, A.; Gorrini, C.; Peruzzi, S.; Piccolo, G.; Dettori, G.; Chezzi, C. An 8-year survey on the occurrence of imported malaria in a nonendemic area by microscopy and molecular assays. Diagn. Microbiol. Infect. Dis. 2008, 61, 434–439. [Google Scholar] [CrossRef]
- Rossi, I.A.; D’Acremont, V.; Prod’Hom, G.; Genton, B. Safety of falciparum malaria diagnostic strategy based on rapid diagnostic tests in returning travelers and migrants: A retrospective study. Malar. J. 2012, 11, 377. [Google Scholar] [CrossRef]
- Kamaliddin, C.; Salnot, V.; Leduc, M.; Ezinmegnon, S.; Broussard, C.; Fievet, N.; Deloron, P.; Guillonneau, F.; Bertin, G.I. PFI1785w: A highly conserved protein associated with pregnancy associated malaria. PLoS ONE 2017, 12, e0187817. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. 2020 Malaria Diagnostic Tests. Available online: https://www.cdc.gov/malaria/diagnosis_treatment/diagnostic_tools.html (accessed on 28 September 2021).
- Moody, A. Rapid diagnostic tests for malaria parasites. Clin. Microbiol. Rev. 2002, 15, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Dhorda, M.; Ba, E.H.; Kevin Baird, J.; Barnwell, J.; Bell, D.; Carter, J.Y.; Dondorp, A.; Ekawati, L.; Gatton, M.; González, I.; et al. Towards harmonization of microscopy methods for malaria clinical research studies. Malar. J. 2020, 19, 324. [Google Scholar] [CrossRef]
- Calderaro, A.; Piccolo, G.; Montecchini, S.; Buttrini, M.; Rossi, S.; Dell’Anna, M.L.; De Remigis, V.; Arcangeletti, M.C.; Chezzi, C.; De Conto, F. High prevalence of malaria in a non-endemic setting: Comparison of diagnostic tools and patient outcome during a four-year survey (2013–2017). Malar. J. 2018, 17, 63. [Google Scholar] [CrossRef]
- Eibach, D.; Traore, B.; Bouchrik, M.; Coulibaly, B.; Coulibaly, N.; Siby, F.; Bonnot, G.; Bienvenu, A.-L.; Picot, S. Evaluation of the malaria rapid diagnostic test VIKIA malaria Ag Pf/PanTM in endemic and non-endemic settings. Malar. J. 2013, 12, 188. [Google Scholar] [CrossRef]
- Garcia, L.S. Diagnostic Medical Parasitology; ASM Press: Washington, DC, USA, 2016. [Google Scholar]
- Gimenez, A.M.; Marques, R.F.; Regiart, M.; Bargieri, D.Y. Diagnostic Methods for Non-Falciparum Malaria. Front. Cell. Infect. Microbiol. 2021, 11, 681063. [Google Scholar] [CrossRef]
- WHO. Malaria Rapid Diagnostic Test Performance. Results of WHO Product Testing of Malaria RDTs: Round 8 (2016–2018). 2018. Available online: https://www.who.int/publications/i/item/9789241514965 (accessed on 25 October 2021).
- Houzé, S.; Boutron, I.; Marmorat, A.; Dalichampt, M.; Choquet, C.; Poilane, I.; Godineau, N.; Le Guern, A.-S.; Thellier, M.; Broutier, H.; et al. Performance of rapid diagnostic tests for imported malaria in clinical practice: Results of a national multicenter study. PLoS ONE 2013, 8, e75486. [Google Scholar] [CrossRef]
- Pasquier, G.; Azoury, V.; Sasso, M.; Laroche, L.; Varlet-Marie, E.; Houzé, S.; Lachaud, L.; Bastien, P.; Sterkers, Y.; Leveque, M.F. Rapid diagnostic tests failing to detect infections by Plasmodium falciparum encoding pfhrp2 and pfhrp3 genes in a non-endemic setting. Malar. J. 2020, 19, 179. [Google Scholar] [CrossRef]
- Cropley, I.M.; Lockwood, D.N.; Mack, D.; Pasvol, G.; Davidson, R.N. Rapid diagnosis of Falciparum malaria by using the ParaSight F test in travellers returning to the United Kingdom: Prospective study. BMJ 2000, 321, 484–485. [Google Scholar] [CrossRef][Green Version]
- Jelinek, T.; Grobusch, M.P.; Nothdurft, H.D. Use of dipstick tests for the rapid diagnosis of malaria in nonimmune travelers. J. Travel Med. 2000, 7, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Ricci, L.; Viani, I.; Piccolo, G.; Fabio, A.; Calderaro, A.; Galati, L.; Perandin, F.; Vecchia, L.; Manca, N.; Dettori, G.; et al. Evaluation of OptiMAL Assay test to detect imported malaria in Italy. New Microbiol. 2000, 23, 391–398. [Google Scholar]
- Rubio, J.M.; Buhigas, I.; Subirats, M.; Baquero, M.; Puente, S.; Benito, A. Limited level of accuracy provided by available rapid diagnosis tests for malaria enhances the need for PCR-based reference laboratories. J. Clin. Microbiol. 2001, 39, 2736–2737. [Google Scholar] [CrossRef] [PubMed]
- Grobusch, M.P.; Hänscheid, T.; Göbels, K.; Slevogt, H.; Zoller, T.; Rögler, G.; Teichmann, D. Comparison of three antigen detection tests for diagnosis and follow-up of falciparum malaria in travellers returning to Berlin, Germany. Parasitol. Res. 2003, 89, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Durand, F.; Crassous, B.; Fricker-Hidalgo, H.; Carpentier, F.; Brion, J.-P.; Grillot, R.; Pelloux, H. Performance of the Now Malaria rapid diagnostic test with returned travellers: A 2-year retrospective study in a French teaching hospital. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2005, 11, 903–907. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, D.P.J.; Gillet, P.; Vlieghe, E.; Cnops, L.; van Esbroeck, M.; Jacobs, J. Evaluation of the Palutop+4 malaria rapid diagnostic test in a non-endemic setting. Malar. J. 2009, 8, 293. [Google Scholar] [CrossRef]
- Bailey, J.W.; Williams, J.; Bain, B.J.; Parker-Williams, J.; Chiodini, P.L. Guideline: The laboratory diagnosis of malaria. General Haematology Task Force of the British Committee for Standards in Haematology. Br. J. Haematol. 2013, 163, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Chilton, D.; Malik, A.N.J.; Armstrong, M.; Kettelhut, M.; Parker-Williams, J.; Chiodini, P.L. Use of rapid diagnostic tests for diagnosis of malaria in the UK. J. Clin. Pathol. 2006, 59, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Berthod, D.; Rochat, J.; Voumard, R.; Rochat, L.; Genton, B.; D’Acremont, V. Self-diagnosis of malaria by travellers: A cohort study on the use of malaria rapid diagnostic tests provided by a Swiss travel clinic. Malar. J. 2017, 16, 436. [Google Scholar] [CrossRef]
- Rubio, J.M.; Benito, A.; Berzosa, P.J.; Roche, J.; Puente, S.; Subirats, M.; López-Vélez, R.; García, L.; Alvar, J. Usefulness of seminested multiplex PCR in surveillance of imported malaria in Spain. J. Clin. Microbiol. 1999, 37, 3260–3264. [Google Scholar] [CrossRef]
- Dakić, Z.; Ivović, V.; Pavlović, M.; Lavadinović, L.; Marković, M.; Djurković-Djaković, O. Clinical significance of molecular methods in the diagnosis of imported malaria in returning travelers in Serbia. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2014, 29, 24–30. [Google Scholar] [CrossRef]
- Calderaro, A.; Piccolo, G.; Gorrini, C.; Montecchini, S.; Rossi, S.; Medici, M.C.; Chezzi, C.; Snounou, G. A new real-time PCR for the detection of Plasmodium ovale wallikeri. PLoS ONE 2012, 7, e48033. [Google Scholar] [CrossRef]
- Paglia, M.G.; Vairo, F.; Bevilacqua, N.; Ghirga, P.; Narciso, P.; Severini, C.; Nicastri, E. Molecular diagnosis and species identification of imported malaria in returning travellers in Italy. Diagn. Microbiol. Infect. Dis. 2012, 72, 175–180. [Google Scholar] [CrossRef]
- Calderaro, A.; Piccolo, G.; Gorrini, C.; Rossi, S.; Montecchini, S.; Dell’Anna, M.L.; De Conto, F.; Medici, M.C.; Chezzi, C.; Arcangeletti, M.C. Accurate identification of the six human Plasmodium spp. causing imported malaria, including Plasmodium ovale wallikeri and Plasmodium knowlesi. Malar. J. 2013, 12, 321. [Google Scholar] [CrossRef] [PubMed]
- Haanshuus, C.G.; Mohn, S.C.; Mørch, K.; Langeland, N.; Blomberg, B.; Hanevik, K. A novel, single-amplification PCR targeting mitochondrial genome highly sensitive and specific in diagnosing malaria among returned travellers in Bergen, Norway. Malar. J. 2013, 12, 26. [Google Scholar] [CrossRef]
- Marti, H.; Stalder, C.; González, I.J. Diagnostic accuracy of a LAMP kit for diagnosis of imported malaria in Switzerland. Travel Med. Infect. Dis. 2015, 13, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Cuadros, J.; Martin Ramírez, A.; González, I.J.; Ding, X.C.; Perez Tanoira, R.; Rojo-Marcos, G.; Gómez-Herruz, P.; Rubio, J.M. LAMP kit for diagnosis of non-falciparum malaria in Plasmodium ovale infected patients. Malar. J. 2017, 16, 20. [Google Scholar] [CrossRef] [PubMed]
- Martín-Díaz, A.; Rubio, J.M.; Herrero-Martínez, J.M.; Lizasoain, M.; Ruiz-Giardin, J.M.; Jaqueti, J.; Cuadros, J.; Rojo-Marcos, G.; Martín-Rabadán, P.; Calderón, M.; et al. Study of the diagnostic accuracy of microbiological techniques in the diagnosis of malaria in the immigrant population in Madrid. Malar. J. 2018, 17, 314. [Google Scholar] [CrossRef]
- Frickmann, H.; Wegner, C.; Ruben, S.; Behrens, C.; Kollenda, H.; Hinz, R.; Rojak, S.; Schwarz, N.G.; Hagen, R.M.; Tannich, E. Evaluation of the multiplex real-time PCR assays RealStar malaria S&T PCR kit 1.0 and FTD malaria differentiation for the differentiation of Plasmodium species in clinical samples. Travel Med. Infect. Dis. 2019, 31, 101442. [Google Scholar] [CrossRef]
- Charpentier, E.; Benichou, E.; Pagès, A.; Chauvin, P.; Fillaux, J.; Valentin, A.; Guegan, H.; Guemas, E.; Salabert, A.-S.; Armengol, C.; et al. Performance evaluation of different strategies based on microscopy techniques, rapid diagnostic test and molecular loop-mediated isothermal amplification assay for the diagnosis of imported malaria. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2020, 26, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Grignard, L.; Nolder, D.; Sepúlveda, N.; Berhane, A.; Mihreteab, S.; Kaaya, R.; Phelan, J.; Moser, K.; van Schalkwyk, D.A.; Campino, S.; et al. A novel multiplex qPCR assay for detection of Plasmodium falciparum with histidine-rich protein 2 and 3 (pfhrp2 and pfhrp3) deletions in polyclonal infections. EBioMedicine 2020, 55, 102757. [Google Scholar] [CrossRef]
- Perandin, F.; Manca, N.; Galati, L.; Piccolo, G.; Calderaro, A.; Viani, I.; Ricci, L.; Dettori, G.; Chezzi, C.; Turano, A. Usefulness of genus-specific PCR and Southern blot species-specific hybridization for the detection of imported malaria cases in Italy. New Microbiol. 2001, 24, 69–76. [Google Scholar]
- Myjak, P.; Nahorski, W.; Pieniazek, N.J.; Pietkiewicz, H. Usefulness of PCR for diagnosis of imported malaria in Poland. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2002, 21, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Schallig, H.D.F.H.; Schoone, G.J.; Lommerse, E.J.M.; Kroon, C.C.M.; de Vries, P.J.; van Gool, T. Usefulness of quantitative nucleic Acid sequence-based amplification for diagnosis of malaria in an academic hospital setting. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2003, 22, 555–557. [Google Scholar] [CrossRef] [PubMed]
- Rougemont, M.; Van Saanen, M.; Sahli, R.; Hinrikson, H.P.; Bille, J.; Jaton, K. Detection of four Plasmodium species in blood from humans by 18S rRNA gene subunit-based and species-specific real-time PCR assays. J. Clin. Microbiol. 2004, 42, 5636–5643. [Google Scholar] [CrossRef] [PubMed]
- Perandin, F.; Manca, N.; Calderaro, A.; Piccolo, G.; Galati, L.; Ricci, L.; Medici, M.C.; Arcangeletti, M.C.; Snounou, G.; Dettori, G.; et al. Development of a real-time PCR assay for detection of Plasmodium falciparum, Plasmodium vivax, and Plasmodium ovale for routine clinical diagnosis. J. Clin. Microbiol. 2004, 42, 1214–1219. [Google Scholar] [CrossRef] [PubMed]
- Mens, P.F.; Schoone, G.J.; Kager, P.A.; Schallig, H.D.F.H. Detection and identification of human Plasmodium species with real-time quantitative nucleic acid sequence-based amplification. Malar. J. 2006, 5, 80. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, N.; Boutet, A.; Bousquet, P.-J.; Basset, D.; Douard-Enault, C.; Charachon, S.; Lachaud, L. Comparison of three real-time PCR methods with blood smears and rapid diagnostic test in Plasmodium sp. infection. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2010, 16, 1305–1311. [Google Scholar] [CrossRef]
- Cnops, L.; Jacobs, J.; Van Esbroeck, M. Validation of a four-primer real-time PCR as a diagnostic tool for single and mixed Plasmodium infections. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2011, 17, 1101–1107. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Evidence Review Group on Malaria Diagnosis in Low Transmission Settings. 2014. Available online: https://www.who.int/malaria/mpac/mpac_mar2014_diagnosis_low_transmission_settings_report.pdf?ua=1 (accessed on 25 October 2021).
- Snounou, G.; Viriyakosol, S.; Zhu, X.P.; Jarra, W.; Pinheiro, L.; do Rosario, V.E.; Thaithong, S.; Brown, K.N. High sensitivity of detection of human malaria parasites by the use of nested polymerase chain reaction. Mol. Biochem. Parasitol. 1993, 61, 315–320. [Google Scholar] [CrossRef]
- Singh, B.; Sung, L.K.; Matusop, A.; Radhakrishnan, A.; Shamsul, S.S.G.; Cox-singh, J.; Thomas, A. A large focus of naturally acquired Plasmodium knowlesi infections in human beings. Lancet 2003, 362, 1504. [Google Scholar] [CrossRef]
- Zimmerman, P.A.; Howes, R.E. Malaria diagnosis for malaria elimination. Curr. Opin. Infect. Dis. 2015, 28, 446–454. [Google Scholar] [CrossRef]
- Haanshuus, C.G.; Mørch, K. Detection of remaining Plasmodium DNA and gametocytes during follow up after curative malaria treatment among returned travellers in Norway. Malar. J. 2020, 19, 296. [Google Scholar] [CrossRef]
- Mischlinger, J.; Pitzinger, P.; Veletzky, L.; Groger, M.; Zoleko-Manego, R.; Adegnika, A.A.; Agnandji, S.T.; Lell, B.; Kremsner, P.G.; Mombo-Ngoma, G.; et al. Validity and reliability of methods to microscopically detect and quantify malaria parasitaemia. Trop. Med. Int. Health 2018, 23, 980–991. [Google Scholar] [CrossRef]
- Frickmann, H.; Wegner, C.; Ruben, S.; Loderstädt, U.; Tannich, E. A comparison of two PCR protocols for the differentiation of Plasmodium ovale species and implications for clinical management in travellers returning to Germany: A 10-year cross-sectional study. Malar. J. 2019, 18, 272. [Google Scholar] [CrossRef] [PubMed]
- Lau, Y.-L.; Lai, M.-Y.; Fong, M.-Y.; Jelip, J.; Mahmud, R. Loop-Mediated Isothermal Amplification Assay for Identification of Five Human Plasmodium Species in Malaysia. Am. J. Trop. Med. Hyg. 2016, 94, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Jarra, W.; Snounou, G. Only viable parasites are detected by PCR following clearance of rodent malarial infections by drug treatment or immune responses. Infect. Immun. 1998, 66, 3783–3787. [Google Scholar] [CrossRef] [PubMed]
- Padley, D.J.; Heath, A.B.; Sutherland, C.; Chiodini, P.L.; Baylis, S.A. Establishment of the 1st World Health Organization International Standard for Plasmodium falciparum DNA for nucleic acid amplification technique (NAT)-based assays. Malar. J. 2008, 7, 139. [Google Scholar] [CrossRef] [PubMed]
- Arai, M.; Kunisada, K.; Kawai, S.; Kimura, M.; Wataya, Y. DNA Diagnosis of Ovale Malaria and Malariae Malaria Using Microtiter Plate-Hybridization. Nucleosides Nucleotides 1994, 13, 1363–1374. [Google Scholar] [CrossRef]
- Kawai, S.; Maekawajiri, S.; Yamane, A. A simple method of detecting amplified DNA with immobilized probes on microtiter wells. Anal. Biochem. 1993, 209, 63–69. [Google Scholar] [CrossRef]
- Elsayed, S.; Plewes, K.; Church, D.; Chow, B.; Zhang, K. Use of molecular beacon probes for real-time PCR detection of Plasmodium falciparum and other plasmodium species in peripheral blood specimens. J. Clin. Microbiol. 2006, 44, 622–624. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, J.; Wirtz, R.A.; McConkey, G.A.; Sattabongkot, J.; Waters, A.P.; Rogers, M.J.; McCutchan, T.F. Plasmodium: Genus-conserved primers for species identification and quantitation. Exp. Parasitol. 1995, 81, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Polley, S.D.; Mori, Y.; Watson, J.; Perkins, M.D.; González, I.J.; Notomi, T.; Chiodini, P.L.; Sutherland, C.J. Mitochondrial DNA targets increase sensitivity of malaria detection using loop-mediated isothermal amplification. J. Clin. Microbiol. 2010, 48, 2866–2871. [Google Scholar] [CrossRef]
- Padley, D.; Moody, A.H.; Chiodini, P.L.; Saldanha, J. Use of a rapid, single-round, multiplex PCR to detect malarial parasites and identify the species present. Ann. Trop. Med. Parasitol. 2003, 97, 131–137. [Google Scholar] [CrossRef]
- Bauffe, F.; Desplans, J.; Fraisier, C.; Parzy, D. Real-time PCR assay for discrimination of Plasmodium ovale curtisi and Plasmodium ovale wallikeri in the Ivory Coast and in the Comoros Islands. Malar. J. 2012, 11, 307. [Google Scholar] [CrossRef] [PubMed]
- Calderaro, A.; Piccolo, G.; Perandin, F.; Gorrini, C.; Peruzzi, S.; Zuelli, C.; Ricci, L.; Manca, N.; Dettori, G.; Chezzi, C.; et al. Genetic polymorphisms influence Plasmodium ovale PCR detection accuracy. J. Clin. Microbiol. 2007, 45, 1624–1627. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Laveran and the Discovery of the Malaria Parasite. 2015. Available online: https://www.cdc.gov/malaria/about/history/laveran.html (accessed on 25 October 2021).
- Mens, P.F.; Matelon, R.J.; Nour, B.Y.M.; Newman, D.M.; Schallig, H.D.F.H. Laboratory evaluation on the sensitivity and specificity of a novel and rapid detection method for malaria diagnosis based on magneto-optical technology (MOT). Malar. J. 2010, 9, 207. [Google Scholar] [CrossRef]
- Hänscheid, T.; Valadas, E.; Grobusch, M.P. Automated malaria diagnosis using pigment detection. Parasitol. Today 2000, 16, 549–551. [Google Scholar] [CrossRef]
- Arndt, L.; Koleala, T.; Orbán, Á.; Ibam, C.; Lufele, E.; Timinao, L.; Lorry, L.; Butykai, Á.; Kaman, P.; Molnár, A.P.; et al. Magneto-optical diagnosis of symptomatic malaria in Papua New Guinea. Nat. Commun. 2021, 12, 969. [Google Scholar] [CrossRef] [PubMed]
- Di Gregorio, E.; Ferrauto, G.; Schwarzer, E.; Gianolio, E.; Valente, E.; Ulliers, D.; Aime, S.; Skorokhod, O. Relaxometric studies of erythrocyte suspensions infected by Plasmodium falciparum: A tool for staging infection and testing anti-malarial drugs. Magn. Reson. Med. 2020, 84, 3366–3378. [Google Scholar] [CrossRef]
- Gupta, M.; Singh, K.; Lobiyal, D.K.; Safvan, C.P.; Sahu, B.K.; Yadav, P.; Singh, S. A sensitive on-chip probe–based portable nuclear magnetic resonance for detecting low parasitaemia plasmodium falciparum in human blood. Med. Devices Sens. 2020, 3, e10098. [Google Scholar] [CrossRef]
- Wang, S.; Yang, C.; Preiser, P.; Zheng, Y. A Photoacoustic-Surface-Acoustic-Wave Sensor for Ring-Stage Malaria Parasite Detection. IEEE Trans. Circuits Syst. II Express Briefs 2020, 67, 881–885. [Google Scholar] [CrossRef]
- Hänscheid, T.; Melo-Cristino, J.; Pinto, B.G. Automated detection of malaria pigment in white blood cells for the diagnosis of malaria in Portugal. Am. J. Trop. Med. Hyg. 2001, 64, 290–292. [Google Scholar] [CrossRef] [PubMed][Green Version]
- WHO. Universal Access to Malaria Diagnostic Testing: An Operational Manual. 2013. Available online: https://apps.who.int/iris/bitstream/handle/10665/44657/9789241502092_eng.pdf?sequence=1&isAllowed=y (accessed on 25 October 2021).
- Mbanefo, A.; Kumar, N. Evaluation of Malaria Diagnostic Methods as a Key for Successful Control and Elimination Programs. Trop. Med. Infect. Dis. 2020, 5, 102. [Google Scholar] [CrossRef] [PubMed]
- WHO. New Perspectives Malaria Diagnosis. 2000. Available online: https://www.who.int/tdr/publications/documents/malaria-diagnosis.pdf (accessed on 25 October 2021).
- Jain, S.; Nehra, M.; Kumar, R.; Dilbaghi, N.; Hu, T.; Kumar, S.; Kaushik, A.; Li, C.-Z. Internet of medical things (IoMT)-integrated biosensors for point-of-care testing of infectious diseases. Biosens. Bioelectron. 2021, 179, 113074. [Google Scholar] [CrossRef]






| Evaluated Assay | Antigen Detected | Country | Samples/Patients Tested | Period | Reference Test | Performance/ Agreement with Reference Test | Reference |
|---|---|---|---|---|---|---|---|
| ParaSight-F (Becton Dickinson) | PfHRP2 | Spain | 206 samples/ 169 patients with suspected imported malaria | 1998–1999 | Microscopy PCR | Sensitivity 76.8% Specificity 76.7% PPV 65.4% NPV 85.2% Sensitivity 67.8% Specificity 77.1% PPV 70.4% NPV 75% | [30] |
| ICT Malaria Pf/Pv (Amrad) | PfHRP2/ pan-aldolase | Microscopy PCR | Sensitivity 81.2% Specificity 97.4% PPV 92.8% NPV 92.5% Sensitivity 68.4% Specificity 97.1% PPV 92.8% NPV 85% | ||||
| OPTIMAL® (Flow Incorporated) | PfLDH/pLDH | Microscopy PCR | Sensitivity 66.1% Specificity 81.5% PPV 62.1% NPV 84% Sensitivity 62.7% Specificity 84.4% PPV 71.2% NPV 78.6% | ||||
| ParaSight F (Becton Dickinson) | PfHRP2 | United Kingdom | 160 patients with suspected malaria | 1994–1996 | Microscopy | Sensitivity 93.3% Specificity 98.3% PPV 95.6% NPV 97.4% | [27] |
| ParaSight-F (Becton Dickinson) | PfHRP2 | Italy | 139 samples/patients with suspected imported malaria | 1994–1999 | Microscopy | Sensitivity 50–100% | [29] |
| OPTIMAL® (Flow Incorporated) | PfLDH/pLDH | Sensitivity 0–100% | |||||
| ICT Malaria Pf® (ICT Diagnostics) | PfHRP2/ pan-aldolase | Germany | 231 patients with suspected imported malaria | 10 years | Microscopy | Sensitivity 92.5% Specificity 98.3% PPV 94.2% NPV 97.8% | [28] |
| OPTIMAL® (Flow Inc.) | PfLDH/pLDH | Sensitivity 88.7% Specificity 99.4% PPV 97.9% NPV 96.7% | |||||
| ParaSight-F (Becton Dickinson) | PfHRP2 | Germany | 1073 samples/ 850 patients with suspected imported malaria | 1998–2001 | Microscopy | Sensitivity 95.1% Specificity 97.1% | [31] |
| ICT Malaria Pf (ICT Diagnostics) | PfHRP2 | Sensitivity 90.6% Specificity 99.4% | |||||
| ICT Malaria Pf/Pv (ICT Diagnostics) | PfHRP2/pan-aldolase | Sensitivity 97.7% Specificity 98.8% | |||||
| OPTIMAL® (Flow Inc.) | PfLDH/pLDH | Sensitivity 76.2% Specificity 99.7% | |||||
| Now Malaria dipstick (Binax) | PfHRP2/ pan-aldolase | France | 413 patients with suspected imported malaria | 2002–2004 | Microscopy/ QBC | Pf Sensitivity 96.4% Specificity 97% PPV 84% NPV 99.4% | [32] |
| Non-Pf Sensitivity 66.7% Specificity 100% PPV 100% NPV 98.2% | |||||||
| Palutop+4 (All.Diag) | PfHRP2/ PvpLDH/pLDH | Belgium | 613 samples selected from international travellers | 1995–2008 | Microscopy corrected by PCR | Pf Sensitivity 85.1% Specificity 96.9% Pv Sensitivity 66% Specificity 100% Po Sensitivity 5.5% Pm Sensitivity 32% Po/Pm Specificity 100% | [33] |
| ICT Malaria Pf/Pv (Amrad) | PfHRP2/ pan-aldolase | Switzerland | 2139 adult patientswith malaria | 1999–2007 | Microscopy | Full agreement: 94% | [15] |
| OPTIMAL® (Diamed) | PfLDH and pLDH | ||||||
| Now ICT Malaria (Binax) | PfHRP2/ pan-aldolase | France | 1311 patients with suspected malaria | 2006–2008 | Microscopy | Sensitivity 93% Specificity 97% | [25] |
| Core Malaria Pan/Pv/Pf (Ivagen) | PfHRP2/pLDH/ PvLDH | Sensitivity 94% Specificity 96% | |||||
| Palutop+4 (All Diag) | PfHRP2/pLDH/ PvLDH | Sensitivity 94% Specificity 97% | |||||
| Optimal-IT (Diamed) | PfLDH/pLDH | Sensitivity 83% Specificity 99% | |||||
| VIKIA Malaria Ag Pf/Pan™ | PfHRP2/ pan-aldolase | France | 155 patients with suspected malaria | 2011 | Microscopy corrected by PCR | Pf or mixed Sensitivity 98% Specificity 93.1% PPV 87.3% NPV 98.9% | [21] |
| Non-Pf Sensitivity 60% Specificity 100% PPV 100% NPV 95.7% | |||||||
| SD Bioline Malaria Ag Pf/Pan | PfHRP2/pLDH | Greece | 955 samples/patients with suspected malaria, residents around the case’s house and residents in regions where autochthonous cases occurred | 2012 | Microscopy | Sensitivity 97.3% Specificity 99.4% PPV 86.1% NPV 99.9% | [12] |
| PCR | Sensitivity 95.6% Specificity 100% PPV 100% NPV 99.8% | ||||||
| ICT Malaria Combo Cassette (ICT Diagnostics)+ SD Bioline Malaria Ag Pf/Pan malaria (Standard Diagnostics) | PfHRP2/ pan-aldolase PfHRP2/ pLDH | France | 446 samples/ patients with imported malaria | 2006–2018 | Microscopy (from 2017 a LAMP assay-Alethia Malaria kit, Meridian®-was associated) | Full agreement: 99.3% | [26] |
| Key Features | Microscopic Examination | Rapid Diagnostic Tests (RDT) | Nucleic Acid Amplification Tests (NAATs) | Future Desired Improvements |
|---|---|---|---|---|
| Sensitivity | 50–200 parasites/µL blood | 200–2000 parasites/µL blood | 0.2–6 parasites/µL blood | RDT sensitivity increasing |
| Specificity | 98% | 68.4–100% (depending on the assay and the species) | 94–100% | RDT discrimination capability for Plasmodia species other than Pf |
| Identification of parasites at species level | Yes (Not distinguish between Poc and Pow) | Not distinguish among Pv/Poc/Pow/Pm. Not able to detect Pk | Yes (distinguish also between Poc and Pow) | RDT discrimination capability increasing and introduction of Pk identification |
| Detection of mixed infections | Low capability | Low capability | High capability | RDT discrimination capability increasing |
| Identification of parasites stages | Yes | No | No | Improvement of the skills of the microscopists in non-endemic settings |
| Turn-around-time | Rapid (suitable to be performed in emergency) | Rapid (suitable to be performed in emergency) | Rapid | NAAT suitable to be performed in emergency |
| Determination of parasitemia | Yes | No | No | NA |
| Monitoring of patients with malaria including follow-up during specific therapy | Yes | No | No | NA |
| Automatization | No | No | Yes | Extension to a wide number of laboratories of automatized tools |
| Instrument required | Yes | No | Yes | NA |
| Trained personnel | Yes | No | No | Improvement of the skills of the microscopists in non-endemic setting |
| Costs | Low | Moderate | High | Reduction of the costs particularly for endemic settings |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calderaro, A.; Montecchini, S.; Buttrini, M.; Piccolo, G.; Rossi, S.; Arcangeletti, M.C.; Farina, B.; De Conto, F.; Chezzi, C. Malaria Diagnosis in Non-Endemic Settings: The European Experience in the Last 22 Years. Microorganisms 2021, 9, 2265. https://doi.org/10.3390/microorganisms9112265
Calderaro A, Montecchini S, Buttrini M, Piccolo G, Rossi S, Arcangeletti MC, Farina B, De Conto F, Chezzi C. Malaria Diagnosis in Non-Endemic Settings: The European Experience in the Last 22 Years. Microorganisms. 2021; 9(11):2265. https://doi.org/10.3390/microorganisms9112265
Chicago/Turabian StyleCalderaro, Adriana, Sara Montecchini, Mirko Buttrini, Giovanna Piccolo, Sabina Rossi, Maria Cristina Arcangeletti, Benedetta Farina, Flora De Conto, and Carlo Chezzi. 2021. "Malaria Diagnosis in Non-Endemic Settings: The European Experience in the Last 22 Years" Microorganisms 9, no. 11: 2265. https://doi.org/10.3390/microorganisms9112265
APA StyleCalderaro, A., Montecchini, S., Buttrini, M., Piccolo, G., Rossi, S., Arcangeletti, M. C., Farina, B., De Conto, F., & Chezzi, C. (2021). Malaria Diagnosis in Non-Endemic Settings: The European Experience in the Last 22 Years. Microorganisms, 9(11), 2265. https://doi.org/10.3390/microorganisms9112265