
Effects of Malic Acid and Sucrose on the Fermentation Parameters, CNCPS Nitrogen Fractions, and Bacterial Community of Moringa oleifera Leaves Silage

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Silage Material and Preparation
2.2. Analyses of the Fermentation Parameter and Chemical Composition
2.3. Analyses of the Bacterial Community
2.4. Statistical Analysis
3. Results
3.1. Fermentation Parameters of MOL Silages
3.2. Chemical Compositions and Nitrogen Fractions of MOL Silage
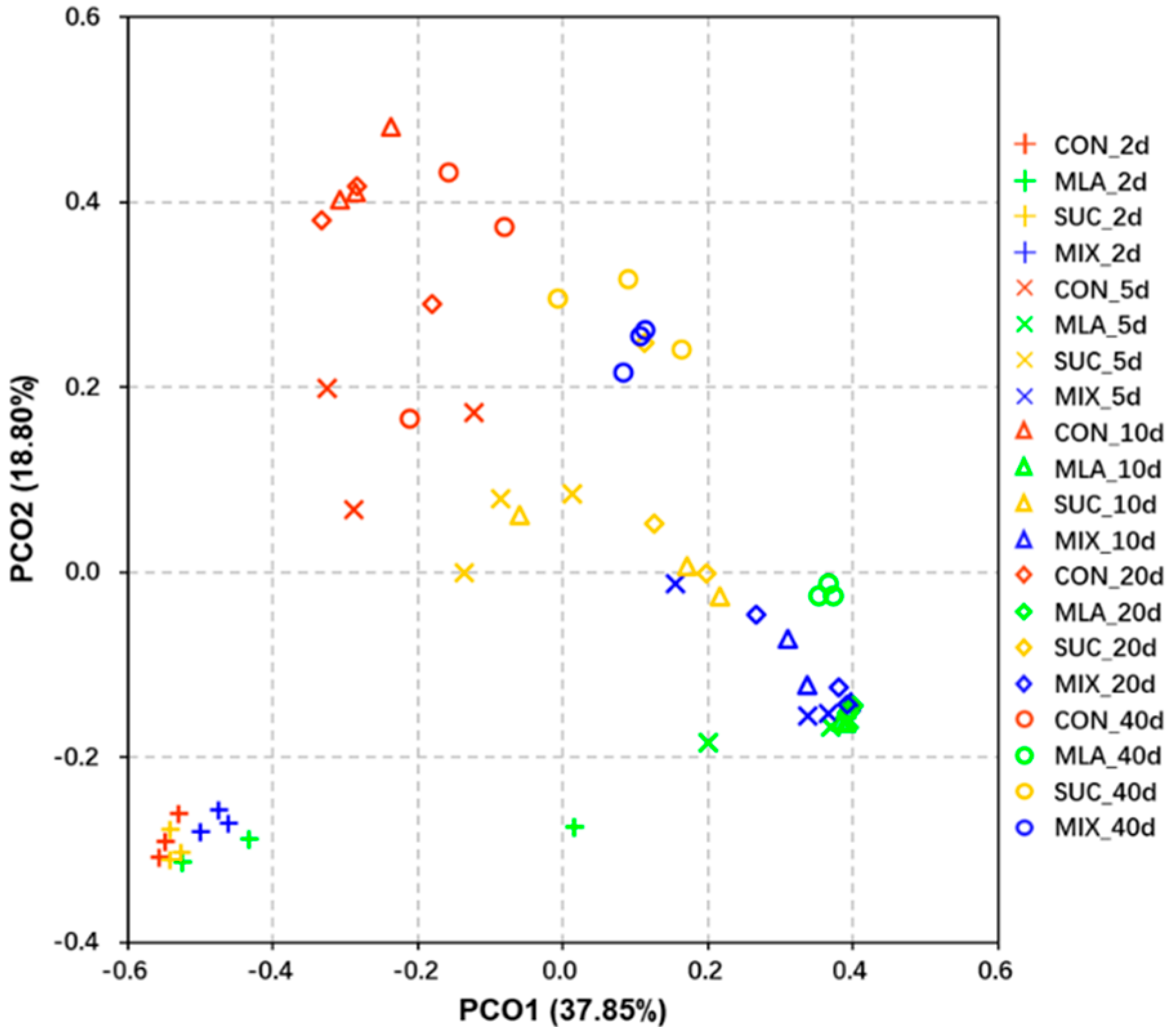
3.3. Diversity of Bacterial Community in MOL Silage
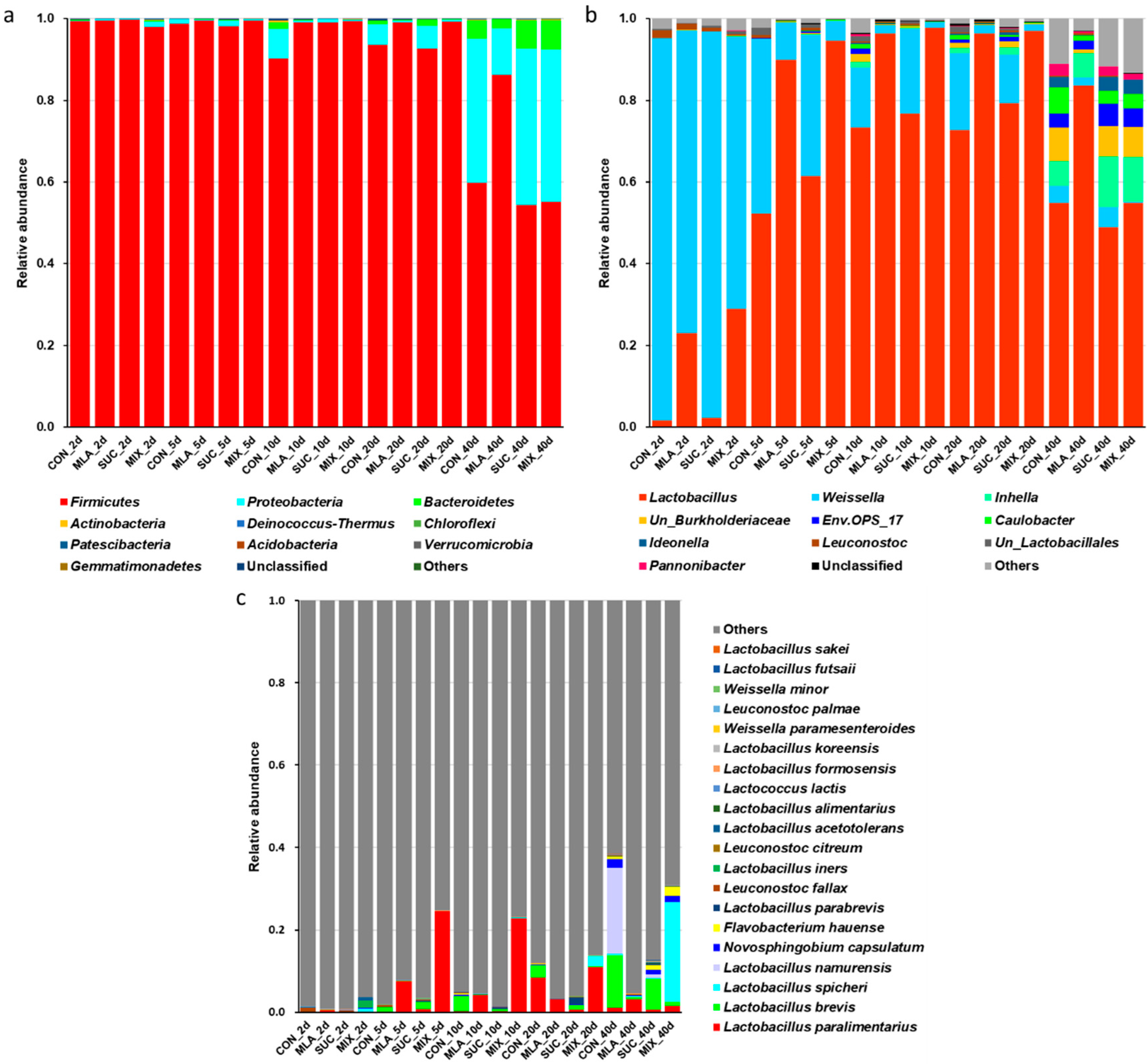
3.4. Composition of the Bacterial Community in MOL Silage
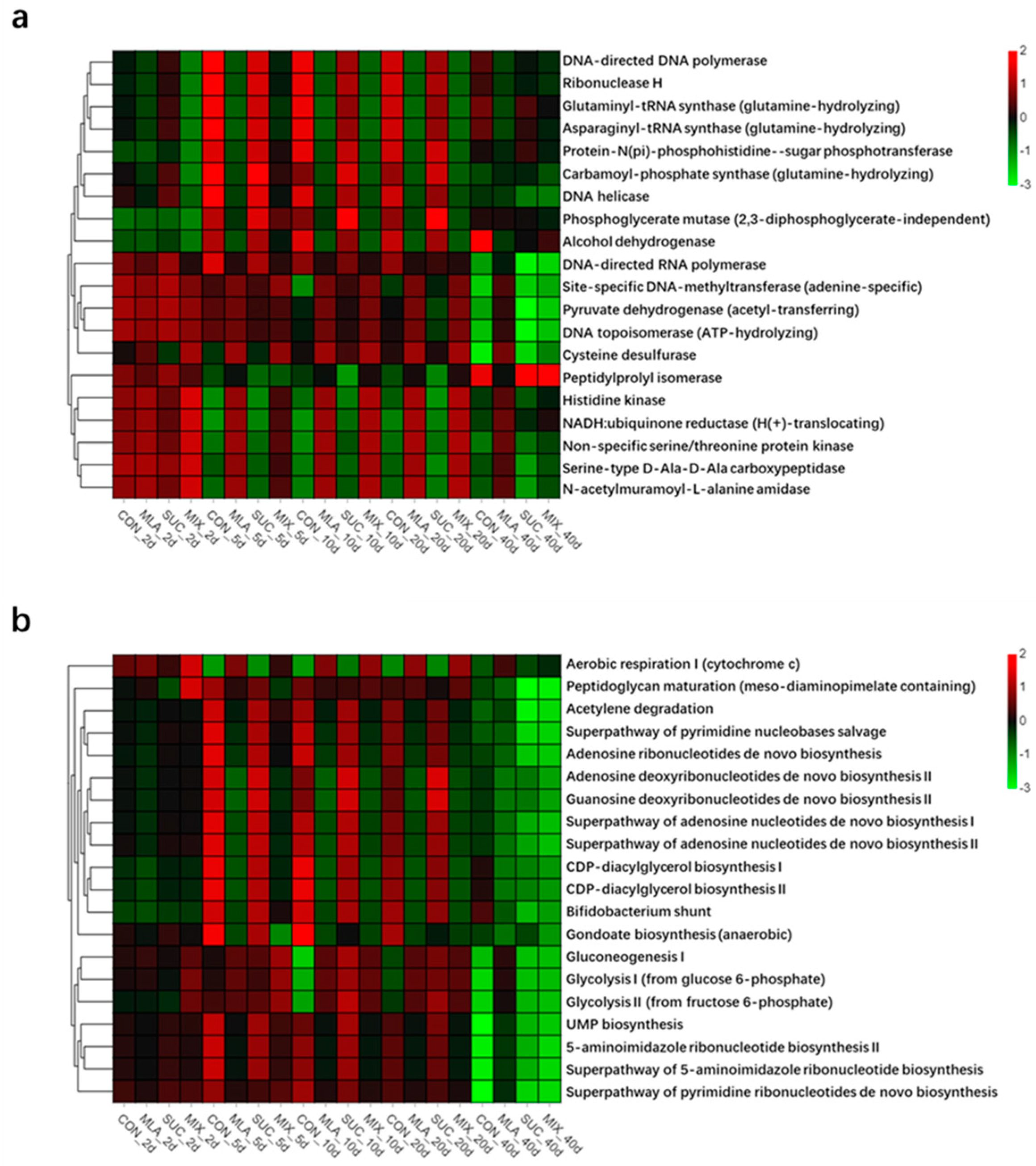
3.5. Predicted Functions and Pathways of Bacterial Community in MOL Silages
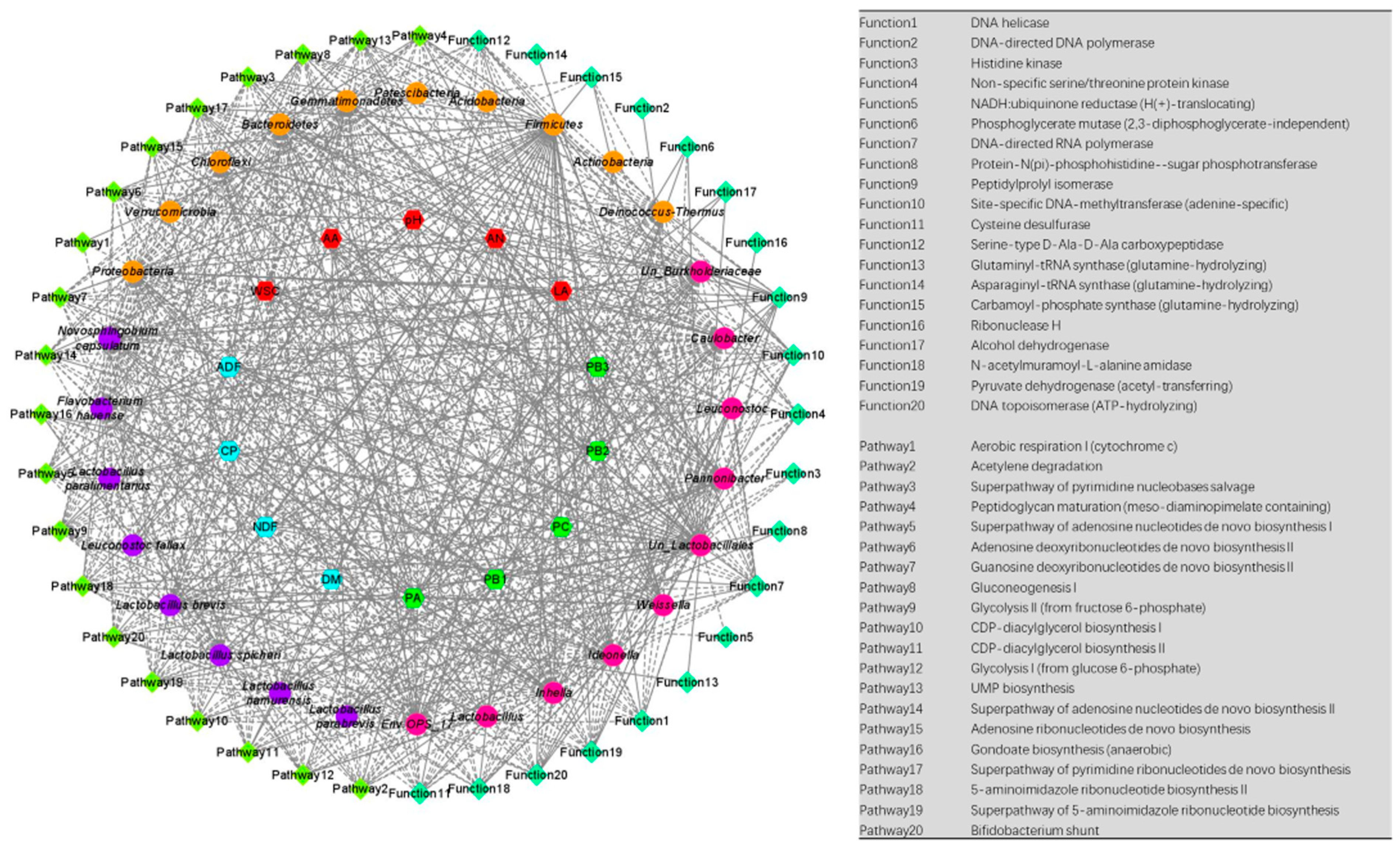
3.6. Correlation Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Çelekli, A.; Al-Nuaimi, A.I.; Bozkurt, H. Adsorption kinetic and isotherms of Reactive Red 120 on Moringa oleifera seed as an eco-friendly process. J. Mol. Struct. 2019, 1195, 168–178. [Google Scholar] [CrossRef]
- Pagano, C.; Perioli, L.; Baiocchi, C.; Bartoccini, A.; Beccari, T.; Blasi, F.; Calarco, P.; Ceccarini, M.R.; Cossignani, L.; di Michele, A.; et al. Preparation and characterization of polymeric microparticles loaded with Moringa oleifera leaf extract for exuding wound treatment. Int. J. Pharmaceut. 2020, 587, 119700. [Google Scholar] [CrossRef] [PubMed]
- Castro-López, C.; Espinoza-González, C.; Ramos-González, R.; Boone-Villa, V.D.; Aguilar-González, M.A.; Martínez-Ávila, G.C.G.; Aguilar, C.N.; Ventura-Sobrevilla, J.M. Spray-drying encapsulation of microwave-assisted extracted polyphenols from Moringa oleifera: Influence of tragacanth, locust bean, and carboxymethyl-cellulose formulations. Food. Res. Int. 2021, 144, 110291. [Google Scholar] [CrossRef] [PubMed]
- Tiloke, C.; Anand, K.; Gengan, R.M.; Chuturgoon, A.A. Moringa oleifera and their phytonanoparticles: Potential antiproliferative agents against cancer. Biomed. Pharmacother. 2018, 108, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Wang, J.; Lu, W.; Zhang, H.; Wu, S.; Qi, G. Effect of dietary supplementation with Moringa oleifera leaf on performance, meat quality, and oxidative stability of meat in broilers. Poult. Sci. 2018, 97, 2836–2844. [Google Scholar] [CrossRef] [PubMed]
- El-Kassas, S.; Abdo, S.E.; Abosheashaa, W.; Mohamed, R.; Moustafa, E.M.; Helal, M.A.; El-Naggar, K. Growth performance, serum lipid profile, intestinal morphometry, and growth and lipid indicator gene expression analysis of mono-sex Nile tilapia fed Moringa oleifera leaf powder. Aquacult. Rep. 2020, 18, 100422. [Google Scholar] [CrossRef]
- Kekana, T.W.; Marume, U.; Muya, M.C.; Nherera-Chokuda, F.V. Periparturient antioxidant enzymes, haematological profile and milk production of dairy cows supplemented with Moringa oleifera leaf meal. Anim. Feed Sci. Tech. 2020, 268, 114606. [Google Scholar] [CrossRef]
- N’nanle, O.; Tété-Bénissan, A.; Nideou, D.; Onagbesan, O.M.; Tona, K. Use of Moringa oleifera leaves in broiler production chain. 1—Effect on Sasso breeder hens performances, internal quality of hatching eggs and serum lipids. Vet. Med. Sci. 2020, 6, 485–490. [Google Scholar] [CrossRef]
- He, L.; Lv, H.; Chen, N.; Wang, C.; Zhou, W.; Chen, X.; Zhang, Q. Improving fermentation, protein preservation and antioxidant activity of Moringa oleifera leaves silage with gallic acid and tannin acid. Bioresour. Technol. 2020, 297, 122390. [Google Scholar] [CrossRef]
- Ren, H.; Feng, Y.; Pei, J.; Li, J.; Wang, Z.; Fu, S.; Zheng, Y.; Li, Z.; Peng, Z. Effects of Lactobacillus plantarum additive and temperature on the ensiling quality and microbial community dynamics of cauliflower leaf silages. Bioresour. Technol. 2020, 307, 123238. [Google Scholar] [CrossRef]
- Wu, Z.; Luo, Y.; Bao, J.; Luo, Y.; Yu, Z. Additives affect the distribution of metabolic profile, microbial communities and antibiotic resistance genes in high-moisture sweet corn kernel silage. Bioresour. Technol. 2020, 315, 123821. [Google Scholar] [CrossRef] [PubMed]
- Tekely, B.-E.; Martău, G.A.; Vodnar, D.C. Physicochemical effects of Lactobacillus plantarum and Lactobacillus casei cocultures on soy–wheat flour dough fermentation. Foods 2020, 9, 1894. [Google Scholar] [CrossRef]
- Ehteshami, S.; Abdollahi, F.; Ramezanian, A.; Rahimzadeh, M.; Dastjerdi, A.M. Maintenance of quality and bioactive compounds of cold stored pomegranate (Punica granatum L.) fruit by organic acids treatment. Food Sci. Technol. Int. 2020, 27, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Ke, W.C.; Ding, W.R.; Ding, L.M.; Xu, D.M.; Zhang, P.; Li, F.H.; Guo, X. Influences of malic acid isomers and their application levels on fermentation quality and biochemical characteristics of alfalfa silage. Anim. Feed Sci. Tech. 2018, 245, 1–9. [Google Scholar] [CrossRef]
- Wang, X.; Liu, H.; Xie, Y.; Zhang, Y.; Lin, Y.; Zheng, Y.; Yang, X.; Wang, N.; Ni, K.; Yang, F. Effect of sucrose and lactic acid bacteria additives on fermentation quality, chemical composition and protein fractions of two typical woody forage silages. Agriculture 2021, 11, 256. [Google Scholar] [CrossRef]
- Dineen, M.; McCarthy, B.; Ross, D.; Ortega, A.; Dillon, B.; Van Amburgh, M.E. Characterization of the nutritive value of perennial ryegrass (Lolium perenne L.) dominated pastures using updated chemical methods with application for the Cornell Net Carbohydrate and Protein System. Anim. Feed Sci. Tech. 2021, 272, 114752. [Google Scholar] [CrossRef]
- Nie, H.; Wang, Z.; You, J.; Zhu, G.; Wang, H.; Wang, F. Comparison of in vitro digestibility and chemical composition among four crop straws treated by Pleurotus ostreatus. Asian Austral. J. Anim. 2020, 33, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Gholizadeh, H.; Naserian, A.A.; Yari, M.; Jonker, A.; Yu, P. Crude protein fractionation, in situ ruminal degradability and FTIR protein molecular structures of different cultivars within barley, corn and sorghum cereal grains. Anim. Feed Sci. Tech. 2021, 275, 114855. [Google Scholar] [CrossRef]
- Rumsey, T.S.; Noller, C.H.; Rhykerd, C.L.; Burns, J.C. Measurement of Certain Metabolic Organic Acids in Forage, Silage, and Ruminal Fluid by Gas-Liquid Chromatography. J. Dairy Sci. 1967, 50, 214–219. [Google Scholar] [CrossRef]
- He, L.; Lv, H.; Xing, Y.; Chen, X.; Zhang, Q. Intrinsic tannins affect ensiling characteristics and proteolysis of Neolamarckia cadamba leaf silage by largely altering bacterial community. Bioresour. Technol. 2020, 311, 123496. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Gaithersburg, VA, USA, 2002. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Licitra, G.; Hernandez, T.M.; Van Soest, P.J. Standardization of procedures for nitrogen fractionation of ruminant feeds. Anim. Feed Sci. Tech. 1996, 57, 347–358. [Google Scholar] [CrossRef]
- Yang, L.; Christensen, D.A.; McKinnon, J.J.; Beattie, A.D.; Yu, P. Effect of altered carbohydrate traits in hulless barley (Hordeum vulgare L.) on nutrient profiles and availability and nitrogen to energy synchronization. J. Cereal Sci. 2013, 58, 182–190. [Google Scholar] [CrossRef]
- Gill, S.R.; Pop, M.; DeBoy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic analysis of the human distal gut microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Jiang, W. Application of high-throughput sequencing in understanding human oral microbiome related with health and disease. Front. Microbiol. 2014, 5, 508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, I.; Batra, T.; Das, S.; Kumar, V. Light at night affects gut microbial community and negatively impacts host physiology in diurnal animals: Evidence from captive zebra finches. Microbiol. Res. 2020, 241, 126597. [Google Scholar] [CrossRef]
- Urbanek, A.K.; Rybak, J.; Wróbel, M.; Leluk, K.; Mirończuk, A.M. A comprehensive assessment of microbiome diversity in Tenebrio molitor fed with polystyrene waste. Environ. Pollut. 2020, 262, 114281. [Google Scholar] [CrossRef]
- De Filippis, F.; Parente, E.; Zotta, T.; Ercolini, D. A comparison of bioinformatic approaches for 16S rRNA gene profiling of food bacterial microbiota. Int. J. Food Microbiol. 2018, 265, 9–17. [Google Scholar] [CrossRef]
- Hu, Z.; Yang, Y.; Lu, L.; Chen, Y.; Zhu, Z.; Huang, J. Kinetics of water absorption expansion of rice during soaking at different temperatures and correlation analysis upon the influential factors. Food Chem. 2021, 346, 128912. [Google Scholar] [CrossRef]
- Guyader, J.; Baron, V.S.; Beauchemin, K.A. Corn forage yield and quality for silage in short growing season areas of the Canadian prairies. Agronomy 2018, 8, 164. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; He, L.; Xing, Y.; Zheng, Y.; Zhou, W.; Pian, R.; Yang, F.; Chen, X.; Zhang, Q. Dynamics of bacterial community and fermentation quality during ensiling of wilted and unwilted Moringa oleifera leaf silage with or without lactic acid bacterial inoculants. mSphere 2019, 4, e00341-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; He, L.; Xing, Y.; Zhou, W.; Pian, R.; Yang, F.; Chen, X.; Zhang, Q. Bacterial diversity and fermentation quality of Moringa oleifera leaves silage prepared with lactic acid bacteria inoculants and stored at different temperatures. Bioresour. Technol. 2019, 284, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, L.; Yu, Z. Fermentation dynamics and bacterial diversity of mixed lucerne and sweet corn stalk silage ensiled at six ratios. Grass Forage Sci. 2019, 74, 264–273. [Google Scholar] [CrossRef]
- Guo, X.S.; Bai, J.; Li, F.H.; Xu, D.M.; Zhang, Y.X.; Bu, D.P.; Zhao, L.S. Effects of malate, citrate, succinate and fumarate on fermentation, chemical composition, aerobic stability and digestibility of alfalfa silage. Anim. Feed Sci. Tech. 2020, 268, 114604. [Google Scholar] [CrossRef]
- Bai, J.; Xu, D.; Xie, D.; Wang, M.; Li, Z.; Guo, X. Effects of antibacterial peptide-producing Bacillus subtilis and Lactobacillus buchneri on fermentation, aerobic stability, and microbial community of alfalfa silage. Bioresour. Technol. 2020, 315, 123881. [Google Scholar] [CrossRef]
- Mu, L.; Xie, Z.; Hu, L.; Chen, G.; Zhang, Z. Cellulase interacts with Lactobacillus plantarum to affect chemical composition, bacterial communities, and aerobic stability in mixed silage of high-moisture amaranth and rice straw. Bioresour. Technol. 2020, 315, 123772. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Wen, A.; Dong, Z.; Desta, S.T.; Shao, T. Effects of formic acid and potassium diformate on the fermentation quality, chemical composition and aerobic stability of alfalfa silage. Grass Forage Sci. 2017, 72, 833–839. [Google Scholar] [CrossRef]
- Wang, C.; Zheng, M.; Wu, S.; Zou, X.; Chen, X.; Ge, L.; Zhang, Q. Effects of Gallic Acid on Fermentation Parameters, Protein Fraction, and Bacterial Community of Whole Plant Soybean Silage. Front. Microbiol. 2021, 12, 662966. [Google Scholar] [CrossRef]
- Ni, K.; Wang, F.; Zhu, B.; Yang, J.; Zhou, G.; Pan, Y.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef]
- Li, M.; Zi, X.; Zhou, H.; Hou, G.; Cai, Y. Effects of sucrose, glucose, molasses and cellulase on fermentation quality and in vitro gas production of king grass silage. Anim. Feed Sci. Tech. 2014, 197, 206–212. [Google Scholar] [CrossRef]
- Zhao, J.; Dong, Z.; Li, J.; Chen, L.; Bai, Y.; Jia, Y.; Shao, T. Effects of lactic acid bacteria and molasses on fermentation dynamics, structural and nonstructural carbohydrate composition and in vitro ruminal fermentation of rice straw silage. Asian Austral. J. Anim. 2019, 32, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Martens, S.D.; Korn, U.; Roscher, S.; Pieper, B.; Schafft, H.; Steinhöfel, O. Effect of tannin extracts on protein degradation during ensiling of ryegrass or lucerne. Grass Forage Sci. 2019, 74, 284–296. [Google Scholar] [CrossRef]
- Dentinho, M.T.P.; Paulos, K.; Portugal, P.V.; Moreira, O.C.; Santos-Silva, J.; Bessa, R.J.B. Proteolysis and in situ ruminal degradation of lucerne ensiled with Cistus ladanifer tannins. Grass Forage Sci. 2019, 74, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Tian, J.; Zhang, Q.; Jiang, Y.; Hou, Z.; Wu, Z.; Yu, Z. Effects of applying Lactobacillus plantarum and Chinese gallnut tannin on the dynamics of protein degradation and proteases activity in alfalfa silage. Grass Forage Sci. 2018, 73, 648–659. [Google Scholar] [CrossRef]
- Brand, T.S.; Jordaan, L. Effect of extrusion on the rumen undegradable protein fraction of lupins. S. Afr. J. Anim. Sci. 2020, 50, 779–785. [Google Scholar]
- Spatharis, S.; Roelke, D.L.; Dimitrakopoulos, P.G.; Kokkoris, G.D. Analyzing the (mis)behavior of Shannon index in eutrophication studies using field and simulated phytoplankton assemblages. Ecol. Indic. 2011, 11, 697–703. [Google Scholar] [CrossRef]
- Sepehri, A.; Sarrafzadeh, M.-H. Activity enhancement of ammonia-oxidizing bacteria and nitrite-oxidizing bacteria in activated sludge process: Metabolite reduction and CO2 mitigation intensification process. Appl. Water Sci. 2019, 9, 131. [Google Scholar] [CrossRef] [Green Version]
- Zi, X.; Li, M.; Chen, Y.; Lv, R.; Zhou, H.; Tang, J. Effects of citric acid and Lactobacillus plantarum on silage quality and bacterial diversity of king grass silage. Front. Microbiol. 2021, 12, 631096. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, C.; Zhou, W.; Yang, F.; Chen, X.; Zhang, Q. Effects of Wilting and Lactobacillus plantarum Addition on the Fermentation Quality and Microbial Community of Moringa oleifera Leaf Silage. Front. Microbiol. 2018, 9, 1817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Dong, Z.; Liu, J.; Shao, T. Microbial community dynamics and their contributions to organic acid production during the early stage of the ensiling of Napier grass (Pennisetum purpureum). Grass Forage Sci. 2020, 75, 37–44. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, J.; Liu, J.; Yang, F.; Zhu, W.; Yuan, X.; Hu, Y.; Cui, Z.; Wang, X. Effect of ensiling and silage additives on biogas production and microbial community dynamics during anaerobic digestion of switchgrass. Bioresour. Technol. 2017, 241, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Sa, D.W.; Lu, Q.; Wang, Z.; Ge, G.; Sun, L.; Jia, Y. The potential and effects of saline-alkali alfalfa microbiota under salt stress on the fermentation quality and microbial. BMC Microbiol. 2021, 21, 149. [Google Scholar] [CrossRef]
- Dong, Z.; Shao, T.; Li, J.; Yang, L.; Yuan, X. Effect of alfalfa microbiota on fermentation quality and bacterial community succession in fresh or sterile Napier grass silages. J. Dairy Sci. 2020, 103, 4288–4301. [Google Scholar] [CrossRef]
- Graf, K.; Ulrich, A.; Idler, C.; Klocke, M. Bacterial community dynamics during ensiling of perennial ryegrass at two compaction levels monitored by terminal restriction fragment length polymorphism. J. Appl. Microbiol. 2016, 120, 1479–1491. [Google Scholar] [CrossRef] [Green Version]
- Dunière, L.; Sindou, J.; Chaucheyras-Durand, F.; Chevallier, I.; Thévenot-Sergentet, D. Silage processing and strategies to prevent persistence of undesirable microorganisms. Anim. Feed Sci. Tech. 2013, 182, 1–15. [Google Scholar] [CrossRef]
- Díaz-Montes, E.; Yáñez-Fernández, J.; Castro-Muñoz, R. Characterization of oligodextran produced by Leuconostoc mesenteroides SF3 and its effect on film-forming properties of chitosan. Mater. Today Commun. 2021, 28, 102487. [Google Scholar] [CrossRef]
- Muraro, G.B.; Carvalho-Estrada, P.D.; Pasetti, M.H.D.; Santos, M.A.; Nussio, L.G. Bacterial dynamics of sugarcane silage in the tropics. Environ. Microbial. 2021. [Google Scholar] [CrossRef]
- Koju, R.; Miao, S.; Liang, B.; Joshi, D.R.; Bai, Y.; Liu, R.; Qu, J. Transcriptional and metabolic response against hydroxyethane-(1,1-bisphosphonic acid) on bacterial denitrification by a halophilic Pannonibacter sp. strain DN. Chemosphere 2020, 252, 126478. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Pei, T.; Du, J.; Deng, M.-R.; Zhu, H. Inhella proteolytica sp. nov. and Inhella gelatinilytica sp. nov., two novel species of the genus Inhella isolated from aquaculture water. Arch. Microbiol. 2021, 203, 3191–3200. [Google Scholar] [CrossRef] [PubMed]
- Berrios, L.; Ely, B. Plant growth enhancement is not a conserved feature in the Caulobacter genus. Plant Soil 2020, 449, 81–95. [Google Scholar] [CrossRef]
- Abdelazez, A.; Abdelmotaal, H.; Evivie, S.E.; Bikheet, M.; Sami, R.; Mohamed, H.; Meng, X. Verification of Lactobacillus brevis tolerance to simulated gastric juice and the potential effects of postbiotic gamma-aminobutyric acid in streptozotocin-induced diabetic mice. Food Sci. Hum. Well. 2022, 11, 165–176. [Google Scholar] [CrossRef]
- Mu, L.; Wang, Q.; Cao, X.; Zhang, Z. Effects of fatty acid salts on fermentation characteristics, bacterial diversity and aerobic stability of mixed silage prepared with alfalfa, rice straw and wheat bran. J. Sci. Food Agric. 2021. [Google Scholar] [CrossRef] [PubMed]
- Barrangou, R.; Yoon, S.S.; Breidt, F.; Fleming, H.P.; Klaenhammer, T.R. Identification and characterization of Leuconostoc fallax strains isolated from an industrial sauerkraut fermentation. Appl. Environ. Microb. 2002, 68, 2877–2884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Wu, Z.; Yu, Z.; Na, R. Effect of lactic acid bacteria inoculants on fermentation characteristics and aerobic stability of three native grasses in Mongolian steppe. Grassl. Sci. 2018, 64, 199–206. [Google Scholar] [CrossRef]
- Daniel, J.L.P.; Checolli, M.; Zwielehner, J.; Junges, D.; Fernandes, J.; Nussio, L.G. The effects of Lactobacillus kefiri and L. brevis on the fermentation and aerobic stability of sugarcane silage. Anim. Feed Sci. Tech. 2015, 205, 69–74. [Google Scholar] [CrossRef]
- Chiş, M.S.; Păucean, A.; Man, S.M.; Mureşan, V.; Socaci, S.A.; Pop, A.; Stan, L.; Rusu, B.; Muste, S. Textural and sensory features changes of gluten free muffins based on rice sourdough fermented with Lactobacillus spicheri DSM 15429. Foods 2020, 9, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Thurber, R.L.V.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items 1 | Groups 2 | Days of Silage | SEM 3 | p-Value 4 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 5 | 10 | 20 | 40 | A | D | A × D | |||
| pH | CON | 4.99 Aa | 4.38 Ba | 4.22 Ca | 4.41 Ba | 4.40 Ba | 0.04 | <0.001 | <0.001 | <0.001 |
| MLA | 4.83 Ab | 4.00 BCc | 3.92 Cb | 4.03 Bc | 4.05 Bb | |||||
| SUC | 4.87 Ab | 4.23 Bb | 4.30 BCa | 4.29 BCb | 4.35 Ca | |||||
| MIX | 4.54 Ac | 3.97 Bc | 3.95 Bb | 3.98 Bc | 4.03 Bb | |||||
| LA (g/kg DM) | CON | 12.35 Eb | 18.29 Db | 34.84 Cc | 60.90 Bc | 68.39 Ab | 3.35 | <0.001 | <0.001 | <0.001 |
| MLA | 17.16 Ea | 25.81 Da | 52.53 Cb | 73.40 Bb | 83.13 Aa | |||||
| SUC | 18.32 Da | 25.48 Ca | 57.93 Ba | 79.81 Aa | 82.19 Aa | |||||
| MIX | 18.72 Da | 27.62 Ca | 57.84 Ba | 79.77 Aa | 81.65 Aa | |||||
| AA (g/kg DM) | CON | 13.82 Ca | 14.52 Ca | 21.45 Aa | 19.63 Bb | 21.06 Aa | 0.78 | <0.001 | <0.001 | <0.001 |
| MLA | 3.86 Bb | 7.97 Ab | 8.61 Ab | 8.03 Ac | 8.75 Ab | |||||
| SUC | 14.43 Ba | 13.99 Ba | 19.50 Aa | 22.14 Aa | 18.95 Aa | |||||
| MIX | 4.51 Bb | 4.72 Bc | 8.39 Ab | 9.28 Ac | 8.70 Ab | |||||
| WSC (g/kg DM) | CON | 26.48 Ab | 19.13 Bb | 14.89 Cb | 11.41 Dc | 9.28 Eb | 0.98 | <0.001 | <0.001 | 0.017 |
| MLA | 34.38 Da | 22.53 Ca | 17.06 Ba | 12.96 Aac | 11.13 Aa | |||||
| SUC | 29.51 Aab | 21.25 Bab | 14.77 Cb | 12.41 CDbc | 10.11 Dab | |||||
| MIX | 31.00 Aab | 24.28 Ba | 14.09 Cb | 13.80 Ca | 10.47 Dab | |||||
| AN (g/kg TN) | CON | 66.24 Ea | 106.51 Da | 95.60 Cb | 130.65 Ba | 155.23 Aa | 5.66 | <0.001 | <0.001 | 0.090 |
| MLA | 16.37 Cb | 38.05 Bb | 41.91 Bc | 54.25 Bb | 83.37 Ab | |||||
| SUC | 58.33 Ca | 94.99 Ba | 113.84 Ba | 140.18 Aa | 156.04 Aa | |||||
| MIX | 11.10 Db | 36.14 Cb | 49.05 BCc | 57.15 Bb | 81.28 Ab | |||||
| Items 1 | Groups 2 | Days of Silage | SEM 3 | p-Value 4 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 5 | 10 | 20 | 40 | A | D | A × D | |||
| DM (g/kg FW) | CON | 224.52 Abc | 221.92 Ac | 216.75 Bb | 226.14 Ab | 226.88 Ab | 1.01 | <0.001 | <0.001 | 0.012 |
| MLA | 237.71 Aab | 228.71 Bb | 229.23 Ba | 233.51 ABa | 234.05 ABb | |||||
| SUC | 221.77 Bc | 229.05 ABb | 225.43 ABa | 231.82 ABa | 234.01 Ab | |||||
| MIX | 244.27 Aa | 236.61 ABa | 228.17 BCa | 233.17 BCa | 244.89 Aa | |||||
| NDF (g/kg DM) | CON | 252.90 Aa | 242.14 Aa | 222.35 Ba | 218.08 BCa | 205.88 Ca | 2.03 | <0.001 | <0.001 | 0.275 |
| MLA | 229.62 Ab | 227.95 ABab | 217.76 ABCa | 210.98 BCab | 203.34 Ca | |||||
| SUC | 211.32 Ac | 211.76 Ab | 207.12 ABa | 204.13 ABb | 194.67 Ba | |||||
| MIX | 227.37 Ab | 221.05 Ab | 215.91 ABa | 217.02 Aab | 199.24 Ba | |||||
| ADF (g/kg DM) | CON | 179.68 Aa | 166.31 Ba | 152.21 Ca | 150.65 Ca | 143.66 Ca | 1.49 | <0.001 | <0.001 | 0.027 |
| MLA | 165.74 Aab | 156.00 ABab | 144.79 BCa | 145.46 BCab | 139.65 Cab | |||||
| SUC | 146.56 Ac | 145.73 Ab | 144.77 Aa | 143.70 Ab | 137.24 Ab | |||||
| MIX | 158.28 Abc | 144.75 BCb | 146.19 Ba | 146.44 Bab | 138.22 Cb | |||||
| CP (g/kg DM) | CON | 273.51 Cc | 278.1 1BCc | 284.83 ABb | 284.57 ABb | 290.49 Aa | 0.96 | <0.001 | <0.001 | 0.163 |
| MLA | 278.00 Db | 283.98 CDb | 287.78 BCb | 291.37 ABab | 294.25 Aa | |||||
| SUC | 287.81 BCa | 284.48 Cb | 292.38 ABab | 295.03 Aa | 293.00 ABa | |||||
| MIX | 284.86 Ba | 294.60 Aa | 297.19 Aa | 297.54 Aa | 295.69 Aa | |||||
| Items 1 | Groups 2 | Days of Silage | SEM 3 | p-Value 4 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 5 | 10 | 20 | 40 | A | D | A × D | |||
| TP (g/kg CP) | CON | 755.66 Ab | 685.51 Bc | 585.91 Cb | 473.88 Db | 386.88 Eb | 16.53 | <0.001 | <0.001 | <0.001 |
| MLA | 813.01 Aa | 770.89 Ba | 745.91 Ba | 655.95 Ca | 576.54 Da | |||||
| SUC | 724.31 Ac | 663.41 Bc | 589.16 Cb | 427.64 Dc | 381.90 Eb | |||||
| MIX | 747.76 Ab | 725.25 Ab | 733.52 Aa | 643.34 Ba | 592.47 Ca | |||||
| NPN (g/kg CP) | CON | 244.34 Eb | 314.49 Da | 414.09 Ca | 526.12 Bb | 613.12 Aa | 16.53 | <0.001 | <0.001 | <0.001 |
| MLA | 186.99 Dc | 229.11 Cc | 254.09 Cb | 344.05 Bc | 423.46 Ab | |||||
| SUC | 275.69 Ea | 336.59 Da | 410.84 Ca | 572.36 Ba | 618.10 Aa | |||||
| MIX | 252.24 Cb | 274.75 Cb | 266.48 Cb | 356.66 Bc | 407.53 Ab | |||||
| SCP (g/kg CP) | CON | 299.67 Eb | 377.28 Da | 479.60 Ca | 594.66 Bb | 689.65 Aa | 17.62 | <0.001 | <0.001 | <0.001 |
| MLA | 228.39 Dc | 270.97 Cc | 298.73 Cb | 392.23 Bc | 474.31 Ab | |||||
| SUC | 334.20 Ea | 392.03 Da | 480.55 Ca | 640.44 Ba | 687.79 Aa | |||||
| MIX | 301.30 Cb | 321.05 Cb | 319.24 Cb | 407.73 Bc | 461.81 Ab | |||||
| NDICP (g/kg CP) | CON | 121.23 Ac | 109.30 Bc | 102.13 Cb | 76.29 Db | 70.15 Ec | 4.26 | <0.001 | <0.001 | <0.001 |
| MLA | 172.86 Aa | 158.49 Ba | 149.73 Ca | 140.49 Da | 128.51 Ea | |||||
| SUC | 120.23 Ac | 92.68 Bd | 76.87 Cc | 68.27 Dc | 65.91 Dc | |||||
| MIX | 164.31 Ab | 132.04 Cb | 141.21 Ba | 135.70 BCa | 104.95 Db | |||||
| ADICP (g/kg CP) | CON | 45.96 Ab | 45.21 Ab | 38.46 Bb | 32.23 Cb | 30.69 Cb | 1.06 | <0.001 | <0.001 | <0.001 |
| MLA | 55.18 Aa | 51.96 Ba | 50.17 Ba | 49.10 BCa | 48.12 Ca | |||||
| SUC | 45.10 Ab | 36.46 Bc | 33.30 Cc | 30.98 Db | 30.96 Db | |||||
| MIX | 54.10 Aa | 50.15 Ba | 49.64 Ba | 48.27 Ba | 45.99 Ca | |||||
| Items 1 | Groups 2 | Days of Silage | SEM 3 | p-Value 4 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 5 | 10 | 20 | 40 | A | D | A × D | |||
| PA (g/kg CP) | CON | 244.34 Eb | 314.49 Da | 414.09 Ca | 526.12 Bb | 613.12 Aa | 16.53 | <0.001 | <0.001 | <0.001 |
| MLA | 186.99 Dc | 229.11 Cc | 254.09 Cb | 344.05 Bc | 423.46 Ab | |||||
| SUC | 275.69 Ea | 336.59 Da | 410.84 Ca | 572.36 Ba | 618.10 Aa | |||||
| MIX | 252.24 Cb | 274.75 Cb | 266.48 Cb | 356.66 Bc | 407.53 Ab | |||||
| PB1 (g/kg CP) | CON | 55.33 Cab | 62.79 BCa | 65.51 Ba | 68.54 ABa | 76.53 Aa | 1.40 | <0.001 | <0.001 | 0.246 |
| MLA | 41.40 Ac | 41.86 Ab | 44.63 Ab | 48.18 Ab | 50.85 Ab | |||||
| SUC | 58.51 Ba | 55.45 Ba | 69.70 Aa | 68.08 Aa | 69.69 Aa | |||||
| MIX | 49.06 ABbc | 46.30 Bb | 52.77 ABb | 51.07 ABb | 54.28 Ab | |||||
| PB2 (g/kg CP) | CON | 579.10 Ab | 513.42 Bb | 418.27 Cb | 329.05 Db | 240.20 Eb | 14.08 | <0.001 | <0.001 | <0.001 |
| MLA | 598.75 Aa | 570.54 ABa | 551.55 Ba | 467.29 Ca | 397.18 Da | |||||
| SUC | 545.56 Ac | 515.28 Bb | 442.58 Cb | 291.29 Db | 246.30 Eb | |||||
| MIX | 534.40 Ac | 546.91 Aab | 539.55 Aa | 456.57 Ba | 433.24 Ba | |||||
| PB3 (g/kg CP) | CON | 75.28 Aa | 64.09 Bc | 63.67 Bb | 44.06 Cb | 39.46 Cc | 3.26 | <0.001 | <0.001 | <0.001 |
| MLA | 117.68 Aa | 106.53 Ba | 99.56 BCa | 91.39 Ca | 80.39 Da | |||||
| SUC | 75.13 Ac | 56.22 Bc | 43.57 Cc | 37.29 CDc | 34.96 Dc | |||||
| MIX | 110.21 Ab | 81.89 Cb | 91.57 Ba | 87.43 BCa | 58.96 Db | |||||
| PC (g/kg CP) | CON | 45.96 Ab | 45.21 Ab | 38.46 Bb | 32.23 Cb | 30.69 Cb | 1.06 | <0.001 | <0.001 | <0.001 |
| MLA | 55.18 Aa | 51.96 Ba | 50.17 Ba | 49.10 BCa | 48.12 Ca | |||||
| SUC | 45.10 Ab | 36.46 Bc | 33.30 Cc | 30.98 Db | 30.96 Db | |||||
| MIX | 54.10 Aa | 50.15 Ba | 49.64 Ba | 48.27 Ba | 45.99 Ca | |||||
| Items | Groups 1 | Days of Silage | SEM 2 | p-Value 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 5 | 10 | 20 | 40 | A | D | A × D | |||
| Observed species | CON | 93.57 Ca | 224.70 Aa | 141.00 BCa | 173.30 ABa | 86.20 Ca | 7.57 | <0.001 | <0.001 | <0.001 |
| MLA | 75.07 Aa | 50.10 ABb | 50.53 ABb | 44.83 Bc | 58.13 ABb | |||||
| SUC | 87.53 Ba | 206.50 Aa | 172.53 Aa | 166.90 Aa | 91.67 Ba | |||||
| MIX | 52.87 Ba | 53.93 ABb | 56.90 ABb | 75.33 Ab | 71.07 ABb | |||||
| Shannon index | CON | 3.13 Ca | 6.11 Aa | 4.58 Ba | 5.29 ABa | 4.14 BCa | 0.18 | <0.001 | 0.005 | <0.001 |
| MLA | 3.39 Aa | 2.30 Bb | 1.73 Bb | 1.55 Bc | 2.20 Bb | |||||
| SUC | 2.64 Ca | 5.68 Aa | 4.89 ABa | 4.79 Ba | 4.50 Ba | |||||
| MIX | 3.48 ABa | 2.46 Cb | 2.72 BCb | 2.99 BCb | 4.11 Aa | |||||
| Simpson index | CON | 0.63 Bab | 0.95 Aa | 0.83 Aab | 0.90 Aa | 0.86 Aa | 0.03 | <0.001 | 0.019 | <0.001 |
| MLA | 0.73 Aab | 0.54 Bb | 0.38 BCc | 0.36 Cc | 0.50 BCb | |||||
| SUC | 0.54 Bb | 0.93 Aa | 0.86 Aa | 0.85 Aa | 0.90 Aa | |||||
| MIX | 0.80 ABa | 0.60 Bb | 0.69 ABb | 0.64 Bb | 0.88 Aa | |||||
| Chao1 index | CON | 178.40 CDa | 419.44 Aa | 252.91 BCa | 320.24 ABa | 143.34 Da | 14.94 | <0.001 | <0.001 | <0.001 |
| MLA | 120.03 Aa | 82.79 Ab | 89.39 Ab | 88.24 Ac | 107.31 Ab | |||||
| SUC | 146.95 Ba | 399.11 Aa | 316.61 Aa | 320.58 Aa | 148.33 Ba | |||||
| MIX | 57.31 Bb | 88.82 ABb | 101.87 ABb | 131.66 Ab | 124.16 Aab | |||||
| Pielou evenness | CON | 0.48 Bab | 0.78 Aa | 0.64 Aa | 0.71 Aa | 0.65 Aa | 0.02 | <0.001 | 0.010 | <0.001 |
| MLA | 0.55 Aab | 0.41 Bb | 0.31 Bc | 0.28 Bc | 0.38 Bb | |||||
| SUC | 0.41 Cb | 0.74 Aa | 0.66 ABa | 0.65 Ba | 0.69 ABa | |||||
| MIX | 0.61 ABa | 0.43 Cb | 0.47 BCb | 0.48 BCb | 0.67 Aa | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, H.; Wang, Y.; Liu, Z.; Hu, Z.; Guo, Y.; Deng, M.; Liu, G.; Sun, B. Effects of Malic Acid and Sucrose on the Fermentation Parameters, CNCPS Nitrogen Fractions, and Bacterial Community of Moringa oleifera Leaves Silage. Microorganisms 2021, 9, 2102. https://doi.org/10.3390/microorganisms9102102
Tian H, Wang Y, Liu Z, Hu Z, Guo Y, Deng M, Liu G, Sun B. Effects of Malic Acid and Sucrose on the Fermentation Parameters, CNCPS Nitrogen Fractions, and Bacterial Community of Moringa oleifera Leaves Silage. Microorganisms. 2021; 9(10):2102. https://doi.org/10.3390/microorganisms9102102
Chicago/Turabian StyleTian, Hanchen, Yue Wang, Zichong Liu, Zhaoying Hu, Yongqing Guo, Ming Deng, Guangbin Liu, and Baoli Sun. 2021. "Effects of Malic Acid and Sucrose on the Fermentation Parameters, CNCPS Nitrogen Fractions, and Bacterial Community of Moringa oleifera Leaves Silage" Microorganisms 9, no. 10: 2102. https://doi.org/10.3390/microorganisms9102102
APA StyleTian, H., Wang, Y., Liu, Z., Hu, Z., Guo, Y., Deng, M., Liu, G., & Sun, B. (2021). Effects of Malic Acid and Sucrose on the Fermentation Parameters, CNCPS Nitrogen Fractions, and Bacterial Community of Moringa oleifera Leaves Silage. Microorganisms, 9(10), 2102. https://doi.org/10.3390/microorganisms9102102