Abstract
Borrelia miyamotoi is classified as a relapsing fever spirochete. Although B. miyamotoi is genetically and ecologically distinct from Borrelia burgdorferi sensu lato, both microorganisms are transmitted by the same Ixodes tick species. B. miyamotoi was detected in I. persulcatus ticks in 1994 in Japan. A phylogenetic analysis based on selected sequences of B. miyamotoi genome revealed genetic differences between isolates from Asia, North America, and Europe, which are clearly separated into three genotypes. Symptomatic human cases of Borrelia miyamotoi disease (BMD) were first reported in 2011 in Russia and then in North America, Europe, and Asia. The most common clinical manifestation of BMD is fever with flu-like symptoms. Several differences in rare symptoms (thrombocytopenia, monocytosis, cerebrospinal fluid pleocytosis, or symptoms related to the central nervous system) have been noted among cases caused by Asian, European, and American types of B. miyamotoi. BMD should be considered in the diagnosis of patients after tick bites, particularly with meningoencephalitis, without anti-Borrelia antibodies in the cerebrospinal fluid. This review describes the biology, ecology, and potential of B. miyamotoi as a tick-borne pathogen of public health concern, with particular emphasis on Europe.
1. Introduction
In Europe, tick-borne diseases transmitted by Ixodes ricinus are the most common zoonoses with significant medical and veterinary importance [1]. This hematophagous arthropod is a reservoir and vector of many pathogenic microorganisms, including the bacteria Borrelia burgdorferi sensu lato (s.l.) complex—the causative agent of Lyme borreliosis (LB), Rickettsia spp., and Anaplasma spp., as well as the flavivirus responsible for tick-borne encephalitis (TBE) and the etiological protozoan agents of babesiosis [2,3]. With advanced methods of molecular biology, new tick-borne microorganism species and their genetic variants with confirmed or potential pathogenicity for humans and animals are still being identified [4]. One of the emerging Ixodes-borne diseases in the northern temperate climate zones of the world, including Europe, is Borrelia miyamotoi disease (BMD), caused by spirochete from the relapsing fever (RF) group of Borrelia [5,6]. Since 1994, when B. miyamotoi was first isolated from questing I. presulcatus ticks and mouse Apodemus argentus in Japan [7], it was considered to be a non-pathogenic endosymbiont. However, since 2011 many symptomatic B. miyamotoi infections in humans have been noted in Asia, North America, and Europe [8,9,10,11,12,13,14].
This review presents data on the biology, ecology, and the potential of B. miyamotoi as a human tick-borne pathogen of public health concern, with particular emphasis on Europe.
2. Review
2.1. Taxonomic Position
B. miyamotoi is a Gram-negative bacteria included in the genus Borrelia from the family Spirochaetaceae, within the phylum Spirochaetes and the order Spirochaetales [15]. Borrelia species are obligate parasites, transmitted by arthropod vectors to vertebrate hosts. The biological feature that distinguishes B. miyamotoi and several other relapsing fever species from B. burgdorferi s.l. is transovarial transmission [16].
The Borrelia spirochete cells are 0.2–0.5 mm in diameter by 3–30 mm in length, with 15–20 periplasmic flagella (endoflagella) located in the periplasmic space between the outer membrane and the protoplasmic cylinder. These cells can move actively with frequent reversal of direction [15,17]. Due to limited B. miyamotoi biosynthetic potential, its in vitro culture is difficult (as other Borrelia species) and requires microaerophilic conditions and complex nutrition. However, it can be propagated in Kelly-Pettenkofer medium with fetal calf serum (MKP-F) [18].
Although the Borrelia species share spirochetal morphology, they have different biological, clinical, and epidemiological features. Based on their arthropod vectors and genetic characteristics two major groups of Borrelia were distinguished. The first group contains 20 Borrelia species, including the B. burgdorferi s.l. complex, an agent of LB, and are transmitted by Ixodes hard ticks. The second group includes 25 Borrelia species associated with human RF and mostly found in soft ticks (Argasidae) but also in lice (B. recurensis) and hard ticks (B. miyamotoi, B. lonestari, B. theileri). In RF-Borrelia complex only B. miyamotoi is transmitted by Ixodes ticks—a vector of B. burgdorferi s.l. complex [15,19,20]. These two groups are genetically similar but form distinct, independent monophyletic clades and share a common ancestor. In 2014, Adeolu and Gupta [21] proposed splitting the spirochetes from the genus Borrelia into two separate genera: a novel genus, Borreliella gen. nov., containing the causative agents of Lyme disease and a revised genus Borrelia, with spirochetes causing RF, including B. miyamotoi. However, the proposed change in the name of this pathogenic bacteria species proved controversial and did not receive support among scientists, clinicians or public health authorities, who felt it would lead to confusion and pose a risk to patient safety [20,22,23].
2.2. Genome Organization and Genetic Diversity
The first information about the organization of the B. miyamotoi genome and its differences in relation to the known species from the LB- and RF-Borrelia groups was published in 1995 [7]. Later, more advanced molecular analysis of Asian, American, and European B. miyamotoi isolates from Ixodes ticks and clinical samples revealed the complexity of the genome structure typical of Borrelia spirochetes [24,25,26,27,28,29]. However, the most information was obtained by sequencing the genome of B. miyamotoi Izh-4 isolate from a Russian patient [30]. The complete genome of a single B. miyamotoi cell consists of one linear chromosome (~900 kb) and 12 linear and two circular plasmids (from 6 to 73 kb). Two of the plasmids (lp70 and lp64) had not previously been found in other Borrelia species. A total of 1362 genes, including 1222 protein-coding genes, 103 pseudogenes, 31 genes for transfer RNA (tRNA), a cluster of three genes of ribosomal RNA (rRNA), and three genes of non-coding RNA (ncRNA) were identified. In B. miyamotoi virulence, a significant role is played by plasmid lp4, which includes genes of variable membrane proteins (VMPs), necessary to mask the bacteria from the host immune system and prolong the infection [30,31,32]. A comparison of different B. miyamotoi isolates revealed that the number and order of VMPs genes were unique for each of them [30].
Phylogenetic analysis based on genome sequences of B. miyamotoi showed genetic differences between isolates from Asia, North America and Europe which are clearly separated into three types (genotypes) and form a monophyletic clade inside the RF-Borrelia spirochetes [30]. However, the genetic differences between the B. miyamotoi isolates are probably not connected with geographic origin, but rather with pathogenicity, vector competence, and host range [16,24].
The B. miyamotoi genetic distance from other LB species and the relationship with the species from the RF group is evidenced by the carriage and expression of a glpQ gene, coding the immunoreactive protein glycerophosphodiester phosphodiesterase [33,34]. The glpQ gene and GlpQ protein are conserved among the members of the genus Borrelia, except LB spirochetes (Figure 1). Therefore, GlpQ is usually used as a marker in molecular and serological tests to detect RF spirochete infections and to distinguish cases of LB and other tick-borne infections (e.g., anaplasmosis, babesiosis) [8,35,36,37].
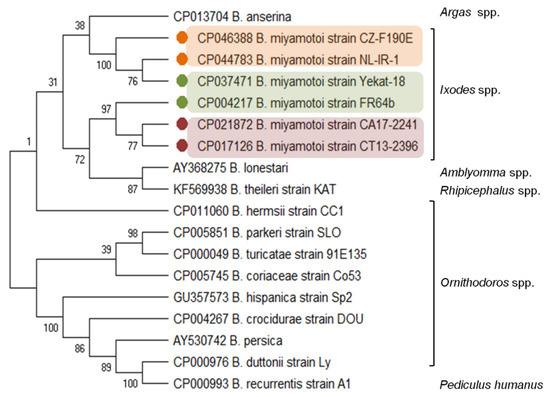
Figure 1.
Molecular relationships between B. miyamotoi and other RF Borrelia species based on the sequences of the glpQ gene selected from GenBank. The consensus tree constructed using the neighbor-joining method and the maximum composite likelihood as the distance method; numbers at the tree nodes indicate bootstrap value from 1000 replicates; analyses were conducted in MEGA X [38]. Marks: orange—European type, green—Asian type, red—American type of B. miyamotoi. The genus names of the vectors were added.
2.3. Vectors and Reservoirs
Since the first detection in 1994 in questing adult I. persulcatus in Japan [7], B. miyamotoi has been recorded in Ixodes ticks from many countries in Asia (Russia, Japan, China, Mongolia, Korea), North America (USA, Canada), and Europe. In Asia, the main vector of B. miyamotoi is I. persulcatus, which is also detected in I. ovatus, I. pavlovskyi, I. nipponensis, and Haemaphysalis concinna [7,39,40,41,42]. I. pacificus is known as a B. miyamotoi vector in the western USA and I. scapularis in the north-central USA and Canada [43,44,45,46,47]. In Europe, vector competence for B. miyamotoi has been demonstrated for I. ricinus and I. persulcatus [12,36,45,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81] (Table 1). Worldwide, B. miyamotoi prevalence in questing Ixodes ticks ranges from 0.2 to 10% [42,43,44,77,82]. This pathogen has been detected in all three tick life stages (larvae, nymphs and adults) [68,83,84]. In European populations of I. ricinus, B. miyamotoi was identified in 0.1–2% larvae [78,85], 0.4–2.8% nymphs [12,36,78,79] and 3.0–4.3% of adults [78,79].

Table 1.
Borrelia miyamotoi in host-seeking and feeding Ixodes ricinus and Ixodes persulcatus ticks in Europe.
The relatively high percentage of naturally infected I. ricinus larvae is an effect of well-documented B. miyamotoi efficient transovarial (vertical) transmission from female ticks to their offspring [43,83]. Van Duijvendijk et al. [85] also reported that the larvae of I. ricinus can transmit B. miyamotoi into nymphs. Transmission and acquisition of the pathogen from rodent hosts to larvae is possible [54]. Ixodes nymphs and adults can also be naturally infected by B. miyamotoi during feeding on vertebrate hosts [85].
Competence as a B. miyamotoi reservoir was demonstrated for Apodemus spp. mice, Myodes glareolus (the bank vole), and Peromyscus leucopus (the white-footed mouse) [43,86]. However, DNA detection of spirochetes in mammals (e.g., squirrels, raccoons, hedgehogs, wild boar, roe deer) and birds (e.g., blackbirds, European robins, European greenfinches, wild turkeys) did not exclude these species as competent reservoirs [6,82,87,88].
Despite demonstrated effective routes of B. miyamotoi transmission, the low infection rate of Ixodes ticks is still unclear. This can be explained by the negative effect of spirochetes on the survival rate of infected ticks or by the low rate of ticks acquired during feeding on infected hosts [44,89]. Moreover, laboratory and field studies revealed that B. miyamotoi does not cause a persistent infection in wild rodents and probably provokes the production of antibodies against B. miyamotoi, making the rodents resistant to infections [85,90]. This may indicate that wild rodents are able to eliminate B. miyamotoi and do not play a significant role in its spread. However, this does not change the fact that B. miyamotoi is constantly present in Ixodes tick populations and can infect humans in all life stages.
2.4. Borrelia miyamotoi Disease (BMD) in Europe
B. miyamotoi is increasingly documented as a human pathogen, especially in the northern hemisphere of the world, where it is co-circulated with B. burgdorferi s.l., bacterium causing LB. Both use the same hard tick species as vectors [6]. In 2011, the first series of 46 patients with febrile diseases caused by B. miyamotoi was described in Yekaterinburg, in the Asian part of Russia [8]. Evidence of B. miyamotoi human infection was then confirmed in the USA, Japan, and China [5,9,10,11,40,91,92].
In Europe, several single cases of BMD have been described [13,14,93,94,95,96]. Human cases of a positive PCR for B. miyamotoi in Europe are summarized in Table 2. BMD is usually manifested by several episodes of fever (~40 °C) and flu-like symptoms [8]. However, symptoms of B. miyamotoi disease are very often non-specific. Meningoencephalitis is one of the potentially dangerous consequences in the course of BMD. In 2013, Hovius et al. [13] reported, for the first time in Europe, meningitis caused by B. miyamotoi infection in an immunocompromised patient (Table 2). A similar case of an immunocompromised individual from New Jersey, USA, was described the same year [92]. Another two cases of meningitis during human BMD were diagnosed in Sweden in 2018. Particularly noteworthy is that, in one of these cases, central nervous system (CNS) symptoms were first diagnosed in an immunocompetent patient [94] (Table 2). Symptoms of BMD were also noted in a seropositive immunocompetent patient in the Netherlands. In this patient, B. miyamotoi–specific PCR of the blood was negative. Moreover, testing for anti-GlpQ and the anti–variable major proteins (VMPs) IgM and IgG using ELISA and Western blot in serum samples demonstrated a clear seroconversion, predominantly for IgG against GlpQ [37]. In turn, BMD-associated neuroborreliosis in an immunocompromised patient was found in 2015 in Germany [93]. In a preliminary report, one case of BMD was demonstrated in the course of neuroborreliosis in Poland [96]. A human case of B. miyamotoi infection was diagnosed in Austria as well (Table 2). Although a phylogenetic analysis of the B. miyamotoi isolate indicated an infection by a European type, the patient claimed a tick bite in the United States. Therefore, this case origin is unclear [95]. Additionally, one blood sample in the Netherlands was found to be PCR-positive and the patient reported no symptoms of BMD but had erythema migrans [14] (Table 2). Due to the coexistence of B. burgdorferi s.l. and B. miyamotoi spirochetes, the correct diagnosis of LB and BMD may cause many difficulties caused by the overlapping manifestation of symptoms of both diseases.
Table 2.
Borrelia miyamotoi disease (BMD) DNA-positive cases in Europe.
An increase in B. miyamotoi infections has been recently recorded during screening tests. A positive Real-Time PCR for B. miyamotoi in the blood from 43 symptomatic patients was revealed in 2020 (Table 2). These studies included 824 patients in expressing signs and symptoms compatible with a persistent polymorphic syndrome, possibly due to a tick bite (neurological/musculoskeletal pain, and cognitive dysfunction, sleep disturbance, and fatigue, lasting for at least six months) and living in different regions of France. According to the authors of the study, the data suggest that B. miyamotoi infection may be persistent and long-term [97].
Few studies in Europe have dealt with the serological evidence of B. miyamotoi. This exposure was found among forestry workers in the Netherlands and febrile patients in Alsace, France [65,98]. More recently, a case of post-tick bite febrile syndrome has been reported in western Europe and serological results suggest that B. miyamotoi was the causative agent of the patient’s symptoms [37]. Reiter et al. [99] studied difference in seroprevalence between distinct populations in Austria by various immunoblotting methods. Antibodies were detected in the sera of 7/53 hunters and in 1/11 sera of Lyme neuroborreliosis patients, 17/74 sera of cases with high concentrations of anti-B. burgdorferi s.l. (α-Bbsl), 7/50 in α-Bbsl negative cases, 5/14 in healthy blood donors from commercial suppliers, and 10/35 from the Austrian Red Cross workers. In the same studies, GlpQ serology was negative in two PCR-positive cases.
2.5. Clinical Manifestation
A diagnosis of BMD should be considered in patients who experience fever attacks and live or spend time in regions where environmental conditions are favorable for ticks and their hosts. [95]. B. miyamotoi infection does not present specific symptoms of the relapsing fever group. Typically, patients experience fever with flu-like symptoms such as chills, headaches, muscle, and joint aches and general fatigue [89,95]. In the available literature, cases with recurrent febrile episodes interspersed with fever-free periods characterizing classic relapsing fever have been rarely reported [6,8,16]. In cases reported in Russia, up to three fever episodes were observed [8]. However, since patients usually receive antibacterial treatment after diagnosis the number of registered febrile episodes may be underestimated. In the case of BMD, there are no other typical symptoms of recurrent fever caused by other spirochetes of this group, such as epistaxis, abortion, jaundice, or severe organ failure. The common features of B. miyamotoi infection and classic relapsing fever include headache, chills, muscle aches, joint pain, and nausea/vomiting [6].
Several differences in rare clinical presentation have been noted between cases caused by Asian, European and American types of B. miyamotoi (Figure 2). One of them is cytopenia (especially thrombocytopenia) which has not been observed in patients infected by the Asian type of B. miyamotoi in Russia, but was recorded in half of the American cases [11,100]. On the other hand, thrombocytopenia was reported in a patient on the island of Hokkaido in Japan, where the Asian type of B. miyamotoi should be genetically close to the Russian isolate. However, BMD was confirmed only by serological test in this case [101]. Increased leukocyte count and thrombocytopenia also occurred occasionally in patients diagnosed with BMD in Northeastern China [40]. A characteristic symptom of the infection of the American type is also monocytosis [91,100]. Erythema migrans, also present in LB, was recorded in Russian and Japanese patients, in which the infection was caused by the Asian type of B. miyamotoi [9,102], as well as in the case of the European type of B. miyamotoi in the Netherlands and France [14,97] (Figure 2).
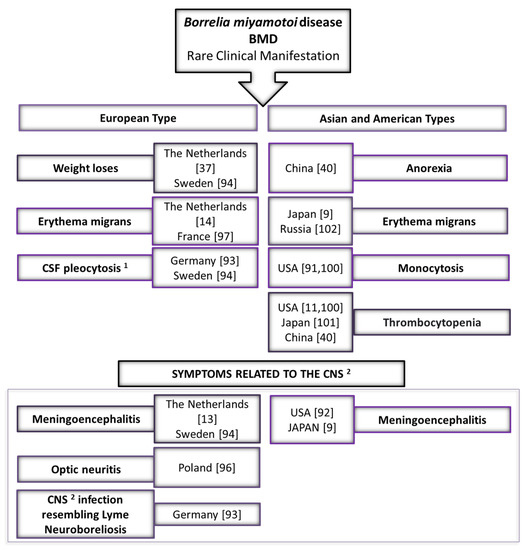
Figure 2.
Rare clinical manifestations of Borrelia miyamotoi disease (BMD)—comparing the European type with Asian (Siberian) and American types. Notes: 1 CSF—cerebrospinal fluid; 2 CNS—central nervous system.
Meningoencephalitis is a common complication in the course of BMD of all three B. miyamotoi types. Cases of meningitis have been reported in the USA, Japan, and in Europe (the Netherlands and Sweden) [9,13,92,94]. Some of the symptoms characteristic for BMD caused by the European type have been observed (Figure 2). For example, two patients lost weight. In the case of a 72-year-old female patient from the Netherlands, it was 2.5 kg within three weeks of a tick bite, while a 66-year-old woman from Stockholm (Sweden) lost 15 kg within six months of illness [37,94] (Figure 2). Although Chinese patients diagnosed with Siberian B. miyamotoi infections have reported anorexia, there is no specific weight loss data available [40]. Cerebrospinal fluid (CSF) pleocytosis was observed in a patient in Germany and a patient in Sweden. Both analyses of CSF showed a total leucocyte count significantly above normal [93,94]. A preliminary report case of BMD has been recently reported in Poland. DNA of B. miyamotoi was detected in the CSF of a patient who had suffered for three months from blurred vision in the left eye. Although the patient did not report typical symptoms of BMD (such as relapsing fever) and did not indicate a tick bite habit in the medical interview, further diagnostics showed that the left eye exhibited extraocular optic neuritis. Brain magnetic resonance imaging (MRI) revealed hyperintense signal abnormalities in the white matter of the brain hemispheres. The optic nerve was thinned and obliterated, which was indicative of fibrosis of the nerve and its sheath. In addition, some demyelinating changes were found in both hemispheres. The authors of the report point to patients with neurological symptoms and questionable serological findings presenting a serious diagnostic problem. This indicates the need for further studies of patients with signs of central nervous system infection [96]. In Germany, another case of B. miyamotoi infection of the CNS resembling neuroborreliosis was investigated [93] (Figure 2).
It appears that co-infection of B. miyamotoi with other tick-borne pathogens does not exacerbate the symptoms of the disease. Patients diagnosed in China who were co-infected with Candidatus Rickettsia tarasevichiae and Anaplasma capra had no more complicated symptoms or prolonged course of BMD [40].
2.6. Diagnosis
The diagnosis of B. miyamotoi infection should always be considered in all patients who live or visit endemic areas in North America, Asia, and Europe [16]. Different mechanisms of infection transmission should be taken into account in distinguishing BMD from other tick-borne diseases. B. miyamotoi infection can be acquired in humans by a bite in any stage of tick development, including the larval stage due to transovarial transmission [41]. Common BMD symptoms, such as recurrent fever, flu-like symptoms, and fatigue are the most important in providing support for a diagnosis. The correct diagnosis can be misleading because patients with other tick-borne diseases, such as LB, human granulocytic anaplasmosis and babesiosis, have similar symptoms [16].
BMD diagnosis is possible through several methods, e.g., by blood smear, different types of PCR, determination of IgG and IgM antibodies, in vitro culture, and/or isolation by animal inoculation. However, two diagnostic methods have been most commonly used to detect B. miyamotoi infections in humans—B. miyamotoi DNA/RNA detection and serodiagnosis [82]. In Europe, enzyme-linked immunosorbent assay (ELISA) and Western-blot serodiagnostic tests and PCR tests (Table 2) are used, either in combination or separately, to detect B. miyamotoi spirochete infection in humans. In ELISA and a confirmatory Western blot, specific antibodies against GlpQ protein, as a non-reactive antigen from B. burgdorferi s.l. [65,94,98], in human serum [10,65,96,98] and in CSF samples [94] were used. Sometimes, however, GlpQ-based B. miyamotoi serology as a single marker does not hold specificity or sensitivity [13,99]. Therefore, searching for other antigens, such as the variable major proteins (VMPs) of B. miyamotoi, should be evaluated as diagnostic additional markers to ensure sufficient specificity for an accurate diagnosis [99]. In such cases, it appears that PCR methods are also more reliable. The amplification of various genes (16S rDNA, fla, p66, 16S–23S internal spacer region) was used by the real-time PCR method in serum and CSF assays [13,95,103]. However, the glpQ gene was most often tested in combination with other genes or individually by qPCR. To determine the B. miyamotoi type, nested PCR [96] or sequencing [95,97] is used.
2.7. Treatment and Prevention
B. miyamotoi infection is generally effectively treated with antibiotics following guidelines used for the treatment of LB [6]. In cases reported in Europe, doses of 200 mg doxycycline have been used successfully once or twice daily for two weeks [94,95]. Intravenous ceftriaxone 2000 mg once a day provided for two weeks has been effectively used in the case of meningoencephalitis diagnosed with BMD [13].
Unfortunately, since no vaccine against Borrelia spirochetes, including B. miyamotoi, has yet been developed or approved [104], it seems that the only effective preventive measures will be the same as for other diseases transmitted by Ixodes ticks, such as LB. These include personal protective measures to avoid tick bites, as well as environmental modification to reduce the number of ticks [16].
3. Conclusions
In Europe, BMD represents an emerging tick-borne disease with an increasing number of diagnosed cases in humans. In the last seven years, B. miyamotoi infection has been noted in both immunocompetent and immunocompromised patients. Fever and other flu-like symptoms suggest a mild infection course. However, serious symptoms related to the central nervous system can be observed. BMD should be considered in the diagnoses of patients after tick bites, particularly with meningoencephalitis, without anti-Borrelia antibodies in CSF. Currently, since there is no specific, reliable serological marker, serodiagnostics should be combined with molecular methods (such as different types of PCR) for a correct diagnosis.
Author Contributions
Conceptualization, K.K. and M.D.; writing—original draft preparation, K.K. and M.D.; writing—review and editing, K.K., M.S. and M.D.; visualization, M.S., K.K.; supervision, K.K. and M.D.; project administration, M.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data presented in this study are openly available.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Mysterud, A.; Jore, S.; Østerås, O.; Viljugrein, H. Emergence of tick-borne diseases at northern latitudes in Europe: A comparative approach. Sci. Rep. 2017, 7, 16316. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F.; Chomel, B.; Otranto, D. Ticks and tick-borne diseases: A One Health perspective. Trends Parasitol. 2012, 28, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubálek, Z.; Földvári, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Špitalská, E.; et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: New hazards and relevance for public health. Front. Public Health 2014, 2, 251. [Google Scholar] [CrossRef] [PubMed]
- Azagi, T.; Hoornstra, D.; Kremer, K.; Hovius, J.W.R.; Sprong, H. Evaluation of disease causality of rare Ixodes ricinus-borne infections in Europe. Pathogens 2020, 9, 150. [Google Scholar] [CrossRef]
- Telford, S.R.; Goethert, H.K.; Molloy, P.J.; Berardi, V.P.; Chowdri, H.R.; Gugliotta, J.L.; Lepore, T.J. Borrelia miyamotoi disease: Neither Lyme disease nor relapsing fever. Clin. Lab. Med. 2015, 35, 867–882. [Google Scholar] [CrossRef]
- Cutler, S.J.; Vayssier-Taussat, M.; Estrada-Peña, A.; Potkonjak, A.; Mihalca, A.D.; Zeller, H. A new Borrelia on the block: Borrelia miyamotoi—A human health risk? Eurosurveillance 2019, 24, 1800170. [Google Scholar] [CrossRef]
- Fukunaga, M.; Takahashi, Y.; Tsuruta, Y.; Matsushita, O.; Ralph, D.; McClelland, M.; Nakao, M. Genetic and phenotypic analysis of Borrelia miyamotoi sp. nov., isolated from the ixodid tick Ixodes persulcatus, the vector for Lyme disease in Japan. Int. J. Syst. Bacteriol. 1995, 45, 804–810. [Google Scholar] [CrossRef]
- Platonov, A.E.; Karan, L.S.; Kolyasnikova, N.M.; Makhneva, N.A.; Toporkova, M.G.; Maleev, V.V.; Fish, D.; Krause, P.J. Humans infected with relapsing fever spirochete Borrelia miyamotoi, Russia. Emerg. Infect. Dis. 2011, 17, 1816–1823. [Google Scholar] [CrossRef]
- Sato, K.; Takano, A.; Konnai, S.; Nakao, M.; Ito, T.; Koyama, K.; Kaneko, M.; Ohnishi, M.; Kawabata, H. Human infections with Borrelia miyamotoi, Japan. Emerg. Infect. Dis. 2014, 20, 1391–1393. [Google Scholar] [CrossRef]
- Krause, P.J.; Narasimhan, S.; Wormser, G.P.; Rollend, L.; Fikrig, E.; Lepore, T.; Barbour, A.; Fish, D. Human Borrelia miyamotoi infection in the United States. N. Engl. J. Med. 2013, 368, 291–293. [Google Scholar] [CrossRef]
- Molloy, P.J.; Telford, S.R., III; Chowdri, H.R.; Lepore, T.J.; Gugliotta, J.L.; Weeks, K.E.; Hewins, M.E.; Goethert, H.K.; Berardi, V.P. Borrelia miyamotoi disease in the Northeastern United States: A case series. Ann. Intern. Med. 2015, 163, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Hansford, K.M.; Fonville, M.; Jahfari, S.; Sprong, H.; Medlock, J.M. Borrelia miyamotoi in host-seeking Ixodes ricinus ticks in England. Epidemiol. Infect. 2015, 143, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Hovius, J.W.R.; De Wever, B.; Sohne, M.; Brouwer, M.C.; Coumou, J.; Wagemakers, A.; Oei, A.; Knol, H.; Narasimhan, S.; Hodiamont, C.J.; et al. A case of meningoencephalitis by the relapsing fever spirochaete Borrelia miyamotoi in Europe. Lancet 2013, 382, 658. [Google Scholar] [CrossRef]
- Jahfari, S.; Hofhuis, A.; Fonville, M.; van der Giessen, J.; van Pelt, W.; Sprong, H. Molecular detection of tick-borne pathogens in humans with tick bites and erythema migrans, in the Netherlands. PLoS Negl. Trop. Dis. 2016, 10, e0005042. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Schwartz, I. Genus Borrelia. In Bergey’s Manual of Systematic Bacteriology Vol. 4: The Bacteroidetes, Spirochaetes, Tenericutes (Mollicutes), Acidobacteria, Fibrobacteres, Fusobacteria, Dictyoglomi, Gemmatimonadetes, Lentisphaerae, Verrucomicrobia, Chlamydiae, and Planctomycetes; Krieg, N., Staley, J., Brown, D., Hedlund, B., Paster, B., Ward, N., Ludwig, W., Whitman, W., Eds.; Springer: New York, NY, USA, 2011; pp. 484–531. ISBN 978-0-387-68572-4. [Google Scholar]
- Krause, P.J.; Fish, D.; Narasimhan, S.; Barbour, A.G. Borrelia miyamotoi infection in nature and in humans. Clin. Microbiol. Infect. 2015, 21, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Wang, G. Borrelia burgdorferi and other Borrelia species. In Molecular Medical Microbiology, 2nd ed.; Tang, Y.-W., Sussman, M., Liu, D., Poxton, I., Schwartzman, J., Eds.; Academic Press: Cambridge, MA, USA, 2015; pp. 1867–1909. ISBN 9780123971692. [Google Scholar]
- Wagemakers, A.; Oei, A.; Fikrig, M.M.; Miellet, W.R.; Hovius, J.W. The relapsing fever spirochete Borrelia miyamotoi is cultivable in a modified Kelly-Pettenkofer medium, and is resistant to human complement. Parasites Vectors 2014, 7, 418. [Google Scholar] [CrossRef]
- Stanek, G.; Reiter, M. The expanding Lyme Borrelia complex-clinical significance of genomic species? Clin. Microbiol. Infect. 2011, 17, 487–493. [Google Scholar] [CrossRef]
- Margos, G.; Marosevic, D.; Cutler, S.; Derdakova, M.; Diuk-Wasser, M.; Emler, S.; Fish, D.; Gray, J.; Hunfeldt, K.P.; Jaulhac, B.; et al. There is inadequate evidence to support the division of the genus Borrelia. Int. J. Syst. Evol. Microbiol. 2017, 67, 1081–1084. [Google Scholar] [CrossRef]
- Adeolu, M.; Gupta, R.S. A phylogenomic and molecular marker based proposal for the division of the genus Borrelia into two genera: The emended genus Borrelia containing only the members of the relapsing fever Borrelia, and the genus Borreliella gen. nov. containing the members of the Lyme disease Borrelia (Borrelia burgdorferi sensu lato complex). Antonie Leeuwenhoek 2014, 105, 1049–1072. [Google Scholar] [CrossRef]
- Margos, G.; Fingerle, V.; Oskam, C.; Stevenson, B.; Gofton, A. Comment on: Gupta, 2019, Distinction between Borrelia and Borreliella is more robustly supported by molecular and phenotypic characteristics than all other neighbouring prokaryotic genera: Response to Margos’ et al. “The genus Borrelia reloaded” PLoS ONE. Ticks Tick Borne Dis. 2020, 11, 101320. [Google Scholar] [CrossRef]
- Stevenson, B.; Fingerle, V.; Wormser, G.P.; Margos, G. Public health and patient safety concerns merit retention of Lyme borreliosis-associated spirochetes within the genus Borrelia, and rejection of the genus novum Borreliella. Ticks Tick Borne Dis. 2019, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Barbour, A.G. Phylogeny of a relapsing fever Borrelia species transmitted by the hard tick Ixodes scapularis. Infect. Genet. Evol. 2014, 27, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Hue, F.; Langeroudi, A.G.; Barbour, A.G. Borne agent of human infection. Genome Announc. 2013, 1, 5–6. [Google Scholar] [CrossRef]
- Kuleshov, K.V.; Koetsveld, J.; Goptar, I.A.; Markelov, M.L.; Kolyasnikova, N.M. Whole-genome sequencing of six Borrelia miyamotoi clinical strains isolated in Russia. Genome Announc. 2018, 6, e01424-17. [Google Scholar] [CrossRef] [PubMed]
- Kingry, L.C.; Replogle, A.; Dolan, M.; Sexton, C.; Padgett, K.A.; Schriefer, M.E. Chromosome and large linear plasmid sequences of a Borrelia miyamotoi strain isolated from Ixodes pacificus ticks from California. Genome Announc. 2017, 5, e00960-17. [Google Scholar] [CrossRef]
- Kingry, L.C.; Replogle, A.; Batra, D.; Rowe, L.A.; Sexton, C.; Dolan, M.; Connally, N.; Petersen, J.M.; Schriefer, M.E. Toward a complete north American Borrelia miyamotoi genome. Genome Announc. 2017, 5, e01557-16. [Google Scholar] [CrossRef] [PubMed]
- Kuleshov, K.V.; Hoornstra, D.; Sprong, H.; Platonov, A.E.; Hovius, J.W. Draft whole-genome sequences of two western European Borrelia miyamotoi isolates. Microbiol. Resour. Announc. 2019, 8, e01314-19. [Google Scholar] [CrossRef]
- Kuleshov, K.V.; Margos, G.; Fingerle, V.; Koetsveld, J.; Goptar, I.A.; Markelov, M.L.; Kolyasnikova, N.M.; Sarksyan, D.S.; Kirdyashkina, N.P.; Shipulin, G.A.; et al. Whole genome sequencing of Borrelia miyamotoi isolate Izh-4: Reference for a complex bacterial genome. BMC Genom. 2020, 21, 16. [Google Scholar] [CrossRef]
- Barbour, A.G. Multiple and diverse vsp and vlp sequences in Borrelia miyamotoi, a hard tick-borne zoonotic pathogen. PLoS ONE 2016, 11, e0146283. [Google Scholar] [CrossRef]
- Bergström, S.; Normark, J. Microbiological features distinguishing Lyme disease and relapsing fever spirochetes. Wien. Klin. Wochenschr. 2018, 130, 484–490. [Google Scholar] [CrossRef]
- Schwan, T.G.; Battisti, J.M.; Porcella, S.F.; Raffel, S.J.; Schrumpf, M.E.; Fischer, E.R.; Carroll, J.A.; Stewart, P.E.; Rosa, P.; Somerville, G.A. Glycerol-3-phosphate acquisition in spirochetes: Distribution and biological activity of glycerophosphodiester phosphodiesterase (GlpQ) among Borrelia species. J. Bacteriol. 2003, 185, 1346–1356. [Google Scholar] [CrossRef] [PubMed]
- Bacon, R.M.; Pilgard, M.A.; Johnson, B.J.B.; Raffel, S.J.; Schwan, T.G. Glycerophosphodiester phosphodiesterase gene (glpQ) of Borrelia lonestari identified as a target for differentiating Borrelia species associated with hard ticks (Acari:Ixodidae). J. Clin. Microbiol. 2004, 42, 2326–2328. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; Carroll, M.; Fedorova, N.; Brancato, J.; Dumouchel, C.; Akosa, F.; Narasimhan, S.; Fikrig, E.; Lane, R.S. Human Borrelia miyamotoi infection in California: Serodiagnosis is complicated by multiple endemic Borrelia species. PLoS ONE 2018, 13, e0191725. [Google Scholar] [CrossRef] [PubMed]
- Wagemakers, A.; Jahfari, S.; de Wever, B.; Spanjaard, L.; Starink, M.V.; de Vries, H.J.C.; Sprong, H.; Hovius, J.W. Borrelia miyamotoi in vectors and hosts in The Netherlands. Ticks Tick Borne Dis. 2017, 8, 370–374. [Google Scholar] [CrossRef] [PubMed]
- Hoornstra, D.; Koetsveld, J.; Sprong, H.; Platonov, A.E.; Hovius, J.W. Borrelia miyamotoi disease in an immunocompetent patient, Western Europe. Emerg. Infect. Dis. 2018, 24, 1770–1772. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Iwabu-Itoh, Y.; Bazartseren, B.; Naranbaatar, O.; Yondonjamts, E.; Furuno, K.; Lee, K.; Sato, K.; Kawabata, H.; Takada, N.; Andoh, M.; et al. Tick surveillance for Borrelia miyamotoi and phylogenetic analysis of isolates in Mongolia and Japan. Ticks Tick Borne Dis. 2017, 8, 850–857. [Google Scholar] [CrossRef]
- Jiang, B.G.; Jia, N.; Jiang, J.F.; Zheng, Y.C.; Chu, Y.L.; Jiang, R.R.; Wang, Y.W.; Liu, H.B.; Wei, R.; Zhang, W.H.; et al. Borrelia miyamotoi infections in humans and ticks, Northeastern China. Emerg. Infect. Dis. 2018, 24, 236–241. [Google Scholar] [CrossRef]
- Takano, A.; Toyomane, K.; Konnai, S.; Ohashi, K.; Nakao, M.; Ito, T.; Andoh, M.; Maeda, K.; Watarai, M.; Sato, K.; et al. Tick surveillance for relapsing fever spirochete Borrelia miyamotoi in Hokkaido, Japan. PLoS ONE 2014, 9, e104532. [Google Scholar] [CrossRef]
- Rar, V.; Livanova, N.; Tkachev, S.; Kaverina, G.; Tikunov, A.; Sabitova, Y.; Igolkina, Y.; Panov, V.; Livanov, S.; Fomenko, N.; et al. Detection and genetic characterization of a wide range of infectious agents in Ixodes pavlovskyi ticks in Western Siberia, Russia. Parasites Vectors 2017, 10, 258. [Google Scholar] [CrossRef]
- Scoles, G.; Papero, M.; Beati, L.; Fish, D. A relapsing fever group spirochete transmitted by Ixodes scapularis ticks. Vector Borne Zoonotic Dis. 2001, 1, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Barbour, A.G.; Bunikis, J.; Travinsky, B.; Hoen, A.G.; Diuk-Wasser, M.A.; Fish, D.; Tsao, J.I. Niche partitioning of Borrelia burgdorferi and Borrelia miyamotoi in the same tick vector and mammalian reservoir species. Am. J. Trop. Med. Hyg. 2009, 81, 1120–1131. [Google Scholar] [CrossRef] [PubMed]
- Crowder, C.D.; Carolan, H.E.; Rounds, M.A.; Honig, V.; Mothes, B.; Haag, H.; Nolte, O.; Luft, B.J.; Grubhoffer, L.; Ecker, D.J.; et al. Prevalence of Borrelia miyamotoi in Ixodes ticks in Europe and the United States. Emerg. Infect. Dis. 2014, 20, 1678–1682. [Google Scholar] [CrossRef] [PubMed]
- Lynn, G.E.; Graham, C.B.; Horiuchi, K.; Eisen, L.; Johnson, T.L.; Lane, R.S.; Eisen, R.J. Prevalence and geographic distribution of Borrelia miyamotoi in host-seeking Ixodes pacificus (Acari: Ixodidae) nymphs in Mendocino County, California Geoffrey. J. Med. Entomol. 2018, 55, 711–716. [Google Scholar] [CrossRef]
- Ogden, N.H.; Margos, G.; Aanensen, D.M.; Drebot, M.A.; Feil, E.J.; Hanincová, K.; Schwartz, I.; Tyler, S.; Lindsay, L.R. Investigation of genotypes of Borrelia burgdorferi in Ixodes scapularis ticks collected during surveillance in Canada. Appl. Environ. Microbiol. 2011, 77, 3244–3254. [Google Scholar] [CrossRef]
- Sormunen, J.J.; Penttinen, R.; Klemola, T.; Hänninen, J.; Vuorinen, I.; Laaksonen, M.; Sääksjärvi, I.E.; Ruohomäki, K.; Vesterinen, E.J. Tick-borne bacterial pathogens in southwestern Finland. Parasites Vectors 2016, 9, 168. [Google Scholar] [CrossRef]
- Kjelland, V.; Rollum, R.; Korslund, L.; Slettan, A.; Tveitnes, D. Borrelia miyamotoi is widespread in Ixodes ricinus ticks in southern Norway. Ticks Tick Borne Dis. 2015, 6, 516–521. [Google Scholar] [CrossRef]
- Kjelland, V.; Paulsen, K.M.; Rollum, R.; Jenkins, A.; Stuen, S.; Soleng, A.; Edgar, K.S.; Lindstedt, H.H.; Vaino, K.; Gibory, M.; et al. Tick-borne encephalitis virus, Borrelia burgdorferi sensu lato, Borrelia miyamotoi, Anaplasma phagocytophilum and Candidatus Neoehrlichia mikurensis in Ixodes ricinus ticks collected from recreational islands in southern Norway. Ticks Tick Borne Dis. 2018, 9, 1098–1102. [Google Scholar] [CrossRef]
- Fraenkel, C.J.; Garpmo, U.; Berglund, J. Determination of novel Borrelia genospecies in Swedish Ixodes ricinus ticks. J. Clin. Microbiol. 2002, 40, 3308–3312. [Google Scholar] [CrossRef]
- Szekeres, S.; Coipan, E.C.; Rigó, K.; Majoros, G.; Jahfari, S.; Sprong, H.; Földvári, G. Eco-epidemiology of Borrelia miyamotoi and Lyme borreliosis spirochetes in a popular hunting and recreational forest area in Hungary. Parasites Vectors 2015, 8, 309. [Google Scholar] [CrossRef]
- Wodecka, B.; Leońska, A.; Skotarczak, B. A comparative analysis of molecular markers for the detection and identification of Borrelia spirochaetes in Ixodes ricinus. J. Med. Microbiol. 2010, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Kiewra, D.; Stańczak, J.; Richter, M. Ixodes ricinus ticks (Acari, Ixodidae) as a vector of Borrelia burgdorferi sensu lato and Borrelia miyamotoi in Lower Silesia, Poland—Preliminary study. Ticks Tick Borne Dis. 2014, 5, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Kowalec, M.; Szewczyk, T.; Welc-Falȩciak, R.; Siński, E.; Karbowiak, G.; Bajer, A. Ticks and the city—Are there any differences between city parks and natural forests in terms of tick abundance and prevalence of spirochaetes? Parasites Vectors 2017, 10, 573. [Google Scholar] [CrossRef] [PubMed]
- Kubiak, K.; Dziekońska-Rynko, J.; Szymańska, H.; Kubiak, D.; Dmitryjuk, M.; Dzika, E. Questing Ixodes ricinus ticks (Acari, Ixodidae) as a vector of Borrelia burgdorferi sensu lato and Borrelia miyamotoi in an urban area of north-eastern Poland. Exp. Appl. Acarol. 2019, 78, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Kalmár, Z.; Sprong, H.; Mihalca, A.D.; Gherman, C.M.; Dumitrache, M.O.; Coipan, E.C.; Fonville, M.; Cozma, V. Borrelia miyamotoi and Candidatus Neoehrlichia mikurensis in Ixodes ricinus ticks, Romania. Emerg. Infect. Dis. 2016, 22, 550–551. [Google Scholar] [CrossRef] [PubMed]
- Hamšíková, Z.; Coipan, C.; Mahríková, L.; Minichová, L.; Sprong, H.; Kazimírová, M. Borrelia miyamotoi and co-infection with Borrelia afzelii in Ixodes ricinus ticks and rodents from Slovakia. Microb. Ecol. 2017, 73, 1000–1008. [Google Scholar] [CrossRef]
- Vaculová, T.; Derdáková, M.; Špitalská, E.; Václav, R.; Chvostáč, M.; Rusňáková Tarageľová, V. Simultaneous occurrence of Borrelia miyamotoi, Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum and Rickettsia helvetica in Ixodes ricinus ticks in urban foci in Bratislava, Slovakia. Acta Parasitol. 2019, 64, 19–30. [Google Scholar] [CrossRef]
- Oechslin, C.P.; Heutschi, D.; Lenz, N.; Tischhauser, W.; Péter, O.; Rais, O.; Beuret, C.M.; Leib, S.L.; Bankoul, S.; Ackermann-Gäumann, R. Prevalence of tick-borne pathogens in questing Ixodes ricinus ticks in urban and suburban areas of Switzerland. Parasites Vectors 2017, 10, 558. [Google Scholar] [CrossRef]
- Cochez, C.; Heyman, P.; Heylen, D.; Fonville, M.; Hengeveld, P.; Takken, W.; Simons, L.; Sprong, H. The Presence of Borrelia miyamotoi, a relapsing fever spirochaete, in questing Ixodes ricinus in Belgium and in The Netherlands. Zoonoses Public Health 2015, 62, 331–333. [Google Scholar] [CrossRef]
- Lernout, T.; De Regge, N.; Tersago, K.; Fonville, M.; Suin, V.; Sprong, H. Prevalence of pathogens in ticks collected from humans through citizen science in Belgium. Parasites Vectors 2019, 12, 550. [Google Scholar] [CrossRef]
- Lejal, E.; Marsot, M.; Chalvet-Monfray, K.; Cosson, J.F.; Moutailler, S.; Vayssier-Taussat, M.; Pollet, T. A three-years assessment of Ixodes ricinus-borne pathogens in a French peri-urban forest. Parasites Vectors 2019, 12, 551. [Google Scholar] [CrossRef]
- Nebbak, A.; Dahmana, H.; Almeras, L.; Raoult, D.; Boulanger, N.; Jaulhac, B.; Mediannikov, O.; Parola, P. Co-infection of bacteria and protozoan parasites in Ixodes ricinus nymphs collected in the Alsace region, France. Ticks Tick Borne Dis. 2019, 10, 101241. [Google Scholar] [CrossRef] [PubMed]
- Boyer, P.H.; Koetsveld, J.; Zilliox, L.; Sprong, H.; Talagrand-Reboul, É.; Hansmann, Y.; De Martino, S.J.; Boulanger, N.; Hovius, J.W.; Jaulhac, B. Assessment of Borrelia miyamotoi in febrile patients and ticks in Alsace, an endemic area for Lyme borreliosis in France. Parasites Vectors 2020, 13, 199. [Google Scholar] [CrossRef] [PubMed]
- Blazejak, K.; Raulf, M.K.; Janecek, E.; Jordan, D.; Fingerle, V.; Strube, C. Shifts in Borrelia burgdorferi (s.l.) geno-species infections in Ixodes ricinus over a 10-year surveillance period in the city of Hanover (Germany) and Borrelia miyamotoi-specific reverse line blot detection. Parasites Vectors 2018, 11, 304. [Google Scholar] [CrossRef] [PubMed]
- Page, S.; Daschkin, C.; Anniko, S.; Krey, V.; Nicolaus, C.; Maxeiner, H.G. First report of Borrelia miyamotoi in an Ixodes ricinus tick in Augsburg, Germany. Exp. Appl. Acarol. 2018, 74, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Geller, J.; Nazarova, L.; Katargina, O.; Järvekülg, L.; Fomenko, N.; Golovljova, I. Detection and Genetic Characterization of Relapsing fever spirochete Borrelia miyamotoi in Estonian ticks. PLoS ONE 2012, 7, e51914. [Google Scholar] [CrossRef] [PubMed]
- Ravagnan, S.; Tomassone, L.; Montarsi, F.; Krawczyk, A.I.; Mastrorilli, E.; Sprong, H.; Milani, A.; Rossi, L.; Capelli, G. First detection of Borrelia miyamotoi in Ixodes ricinus ticks from northern Italy. Parasites Vectors 2018, 11, 130. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.; Parreira, R.; Lopes, N.; Maia, C.; Carreira, T.; Sousa, C.; Faria, S.; Campino, L.; Vieira, M. Molecular identification of Borrelia miyamotoi in Ixodes ricinus from Portugal. Vector Borne Zoonotic Dis. 2015, 15, 515–517. [Google Scholar] [CrossRef] [PubMed]
- Potkonjak, A.; Kleinerman, G.; Gutiérrez, R.; Savić, S.; Vračar, V.; Nachum-Biala, Y.; Jurišić, A.; Rojas, A.; Petrović, A.; Ivanović, I.; et al. Occurrence of Borrelia burgdorferi sensu lato in Ixodes ricinus ticks with first identification of Borrelia miyamotoi in Vojvodina, Serbia. Vector Borne Zoonotic Dis. 2016, 16, 631–635. [Google Scholar] [CrossRef]
- Díaz, P.; Arnal, J.L.; Remesar, S.; Pérez-Creo, A.; Venzal, J.M.; Vázquez-López, M.E.; Prieto, A.; Fernández, G.; López, C.M.; Panadero, R.; et al. Molecular identification of Borrelia spirochetes in questing Ixodes ricinus from northwestern Spain. Parasites Vectors 2017, 10, 615. [Google Scholar] [CrossRef]
- Palomar, A.M.; Portillo, A.; Santibáñez, P.; Santibáñez, S.; Oteo, J.A. Borrelia miyamotoi: Should this pathogen be considered for the diagnosis of tick-borne infectious diseases in Spain? Enferm. Infecc. Microbiol. Clin. 2018, 36, 568–571. [Google Scholar] [CrossRef] [PubMed]
- Remesar, S.; Díaz, P.; Venzal, J.M.; Prieto, A.; Estrada-Peña, A.; López, C.M.; Panadero, R.; Fernández, G.; Díez-Baños, P.; Morrondo, P. Longitudinal study of infection with Borrelia spp. in questing ticks from north-western Spain. Vector Borne Zoonotic Dis. 2019, 19, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, K.; Şen, E.; Sato, K.; Kawabata, H.; Ohashi, N.; Masuzawa, T. Characterization of the emerging relapsing fever pathogen, Borrelia miyamotoi, from the Ixodes ricinus tick in the rural Trakya (Thrace) Region of Northwestern Turkey. Vector Borne Zoonotic Dis. 2016, 16, 797–799. [Google Scholar] [CrossRef] [PubMed]
- Capligina, V.; Seleznova, M.; Akopjana, S.; Freimane, L.; Lazovska, M.; Krumins, R.; Kivrane, A.; Namina, A.; Aleinikova, D.; Kimsis, J.; et al. Large-scale countrywide screening for tick-borne pathogens in field-collected ticks in Latvia during 2017–2019. Parasites Vectors 2020, 13, 351. [Google Scholar] [CrossRef]
- Lambert, J.S.; Cook, M.J.; Healy, J.E.; Murtagh, R.; Avramovic, G.; Lee, S.H. Metagenomic 16S rRNA gene sequencing survey of Borrelia species in Irish samples of Ixodes ricinus ticks. bioRxiv 2018. [Google Scholar] [CrossRef]
- Szekeres, S.; Lügner, J.; Fingerle, V.; Margos, G.; Földvári, G. Prevalence of Borrelia miyamotoi and Borrelia burgdorferi sensu lato in questing ticks from a recreational coniferous forest of East Saxony, Germany. Ticks Tick Borne Dis. 2017, 8, 922–927. [Google Scholar] [CrossRef]
- Rǎileanu, C.; Tauchmann, O.; Vasić, A.; Wöhnke, E.; Silaghi, C. Borrelia miyamotoi and Borrelia burgdorferi (sensu lato) identification and survey of tick-borne encephalitis virus in ticks from north-eastern Germany. Parasites Vectors 2020, 13, 106. [Google Scholar] [CrossRef]
- Michelet, L.; Delannoy, S.; Devillers, E.; Umhang, G.; Aspan, A.; Juremalm, M.; Chirico, J.; van der Wal, F.J.; Sprong, H.; Boye Pihl, T.P.; et al. High-throughput screening of tick-borne pathogens in Europe. Front. Cell. Infect. Microbiol. 2014, 4, 103. [Google Scholar] [CrossRef]
- Layzell, S.J.; Bailey, D.; Peacey, M.; Nuttall, P.A. Prevalence of Borrelia burgdorferi and Borrelia miyamotoi in questing Ixodes ricinus ticks from four sites in the UK. Ticks Tick Borne Dis. 2018, 9, 217–224. [Google Scholar] [CrossRef]
- Siński, E.; Welc-Falȩciak, R.; Zajkowska, J. Borrelia miyamotoi: A human tick-borne relapsing fever spirochete in Europe and its potential impact on public health. Adv. Med. Sci. 2016, 61, 255–260. [Google Scholar] [CrossRef]
- Richter, D.; Debski, A.; Hubalek, Z.; Matuschka, F.-R. Absence of Lyme disease spirochetes in larval Ixodes ricinus ticks. Vector Borne Zoonotic Dis. 2012, 12, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Richter, D.; Schlee, D.B.; Matuschka, F.R. Relapsing fever-like spirochetes infecting European vector tick of Lyme disease agent. Emerg. Infect. Dis. 2003, 9, 697–701. [Google Scholar] [CrossRef] [PubMed]
- Van Duijvendijk, G.; Coipan, C.; Wagemakers, A.; Fonville, M.; Ersöz, J.; Oei, A.; Földvári, G.; Hovius, J.; Takken, W.; Sprong, H. Larvae of Ixodes ricinus transmit Borrelia afzelii and B. miyamotoi to vertebrate hosts. Parasites Vectors 2016, 9, 97. [Google Scholar] [CrossRef] [PubMed]
- Burri, C.; Schumann, O.; Schumann, C.; Gern, L. Are Apodemus spp. mice and Myodes glareolus reservoirs for Borrelia miyamotoi, Candidatus Neoehrlichia mikurensis, Rickettsia helvetica, R. monacensis and Anaplasma phagocytophilum? Ticks Tick Borne Dis. 2014, 5, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Talagrand-Reboul, E.; Boyer, P.H.; Bergström, S.; Vial, L.; Boulanger, N. Relapsing fevers: Neglected tick-borne diseases. Front. Cell. Infect. Microbiol. 2018, 8, 98. [Google Scholar] [CrossRef] [PubMed]
- Majerová, K.; Hönig, V.; Houda, M.; Papežík, P.; Fonville, M.; Sprong, H.; Rudenko, N.; Golovchenko, M.; Černá Bolfíková, B.; Hulva, P.; et al. Hedgehogs, squirrels, and blackbirds as sentinel hosts for active surveillance of Borrelia miyamotoi and Borrelia burgdorferi complex in urban and rural environments. Microorganisms 2020, 8, 1908. [Google Scholar] [CrossRef]
- Wagemakers, A.; Staarink, P.J.; Sprong, H.; Hovius, J.W.R. Borrelia miyamotoi: A widespread tick-borne relapsing fever spirochete. Trends Parasitol. 2015, 31, 260–269. [Google Scholar] [CrossRef]
- Taylor, K.R.; Takano, A.; Konnai, S.; Shimozuru, M.; Kawabata, H.; Tsubota, T. Borrelia miyamotoi infections among wild rodents show age and month independence and correlation with Ixodes persulcatus larval attachment in Hokkaido, Japan. Vector Borne Zoonotic Dis. 2013, 13, 92–97. [Google Scholar] [CrossRef]
- Chowdri, H.R.; Gugliotta, J.L.; Berardi, V.P.; Goethert, H.K.; Molloy, P.J.; Sterling, S.L.; Telford, S.R. Borrelia miyamotoi infection presenting as human granulocytic anaplasmosis: A case report. Ann. Intern. Med. 2013, 159, 21–27. [Google Scholar] [CrossRef]
- Gugliotta, J.L.; Goethert, H.K.; Berardi, V.P.; Telford, S.R. Meningoencephalitis from Borrelia miyamotoi in an immunocompromised patient. N. Engl. J. Med. 2013, 368, 240–245. [Google Scholar] [CrossRef]
- Boden, K.; Lobenstein, S.; Hermann, B.; Margos, G.; Fingerle, V. Borrelia miyamotoi-associated neuroborreliosis in immunocompromised person. Emerg. Infect. Dis. 2016, 22, 1617–1620. [Google Scholar] [CrossRef] [PubMed]
- Henningsson, A.J.; Asgeirsson, H.; Hammas, B.; Karlsson, E.; Parke, Å.; Hoornstra, D.; Wilhelmsson, P.; Hovius, J.W. Two cases of Borrelia miyamotoi meningitis, Sweden, 2018. Emerg. Infect. Dis. 2019, 25, 1965–1968. [Google Scholar] [CrossRef] [PubMed]
- Tobudic, S.; Burgmann, H.; Stanek, G.; Winkler, S.; Schötta, A.; Obermüller, M.; Markowicz, M.; Lagler, H. Human Human Borrelia miyamotoi Infection, AustriaBorrelia miyamotoi Infection, Austria. Emerg. Infect. Dis. 2020, 26, 2201–2204. [Google Scholar] [CrossRef] [PubMed]
- Fiecek, B.; Lewandowska, G.; Roguska, U.; Rozej-Bielicka, W.; Tylewska-Wierzbanowska, S.; Chmielewski, T. Borrelia miyamotoi DNA in a Patient Suspected of Lyme Borreliosis. 2019, pp. 1–14. Available online: https://assets.researchsquare.com/files/rs-5981/v2/manuscript.pdf (accessed on 11 January 2021). [CrossRef]
- Franck, M.; Ghozzi, R.; Pajaud, J.; Lawson-Hogban, N.E.; Mas, M.; Lacout, A.; Perronne, C. Borrelia miyamotoi: 43 cases diagnosed in France by Real-Time PCR in patients with persistent polymorphic signs and symptoms. Front. Med. 2020, 7, 55. [Google Scholar] [CrossRef]
- Jahfari, S.; Herremans, T.; Platonov, A.E.; Kuiper, H.; Karan, L.S.; Vasilieva, O.; Koopmans, M.P.G.; Hovius, J.W.R.; Sprong, H. High seroprevalence of Borrelia miyamotoi antibodies in forestry workers and individuals suspected of human granulocytic anaplasmosis in The Netherlands. New Microbes New Infect. 2014, 2, 144–149. [Google Scholar] [CrossRef][Green Version]
- Reiter, M.; Stelzer, T.; Schötta, A.M.; Markowicz, M.; Leschnik, M.; Harsch, A.; Reiß, E.; Kneusel, R.E.; Stockinger, H.; Stanek, G. Glycerophosphodiester phosphodiesterase identified as non-reliable serological marker for Borrelia miyamotoi disease. Microorganisms 2020, 8, 1846. [Google Scholar] [CrossRef]
- Sudhindra, P.; Wang, G.; Schriefer, M.E.; McKenna, D.; Zhuge, J.; Krause, P.J.; Marques, A.R.; Wormser, G.P. Insights into Borrelia miyamotoi infection from an untreated case demonstrating relapsing fever, monocytosis and a positive C6 Lyme serology. Diagn. Microbiol. Infect. Dis. 2016, 86, 93–96. [Google Scholar] [CrossRef]
- Yamano, K.; Ito, T.; Kiyanagi, K.; Yamazaki, H.; Sugawara, M.; Saito, T.; Ohashi, N.; Zamoto-Niikura, A.; Sato, K.; Kawabata, H. Case report: Clinical features of a case of suspected Borrelia miyamotoi disease in Hokkaido, Japan. Am. J. Trop. Med. Hyg. 2017, 97, 84–87. [Google Scholar] [CrossRef][Green Version]
- Karan, L.; Makenov, M.; Kolyasnikova, N.; Stukolova, O.; Toporkova, M.; Olenkova, O. Dynamics of spirochetemia and early PCR detection of Borrelia miyamotoi. Emerg. Infect. Dis. 2018, 24, 860–867. [Google Scholar] [CrossRef]
- Wagemakers, A.; Koetsveld, J.; Narasimhan, S.; Wickel, M.; Deponte, K.; Bleijlevens, B.; Jahfari, S.; Sprong, H.; Karan, L.S.; Sarksyan, D.S.; et al. Variable major proteins as targets for specific antibodies against Borrelia miyamotoi. J. Immunol. 2016, 196, 4185–4195. [Google Scholar] [CrossRef]
- Wormser, G.P. Doxycycline for prevention of spirochetal infections: Status report. Clin. Infect. Dis. 2020, 71, 2014–2017. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).