EMS-Induced Mutagenesis of Clostridium carboxidivorans for Increased Atmospheric CO2 Reduction Efficiency and Solvent Production


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain, Media, and Culture Conditions
2.2. Ethyl Methanesulfonate (EMS) Mutagenesis of the P7 Population
2.3. End Product Analysis
2.4. CO2 Reduction and Statistical Analysis
3. Results
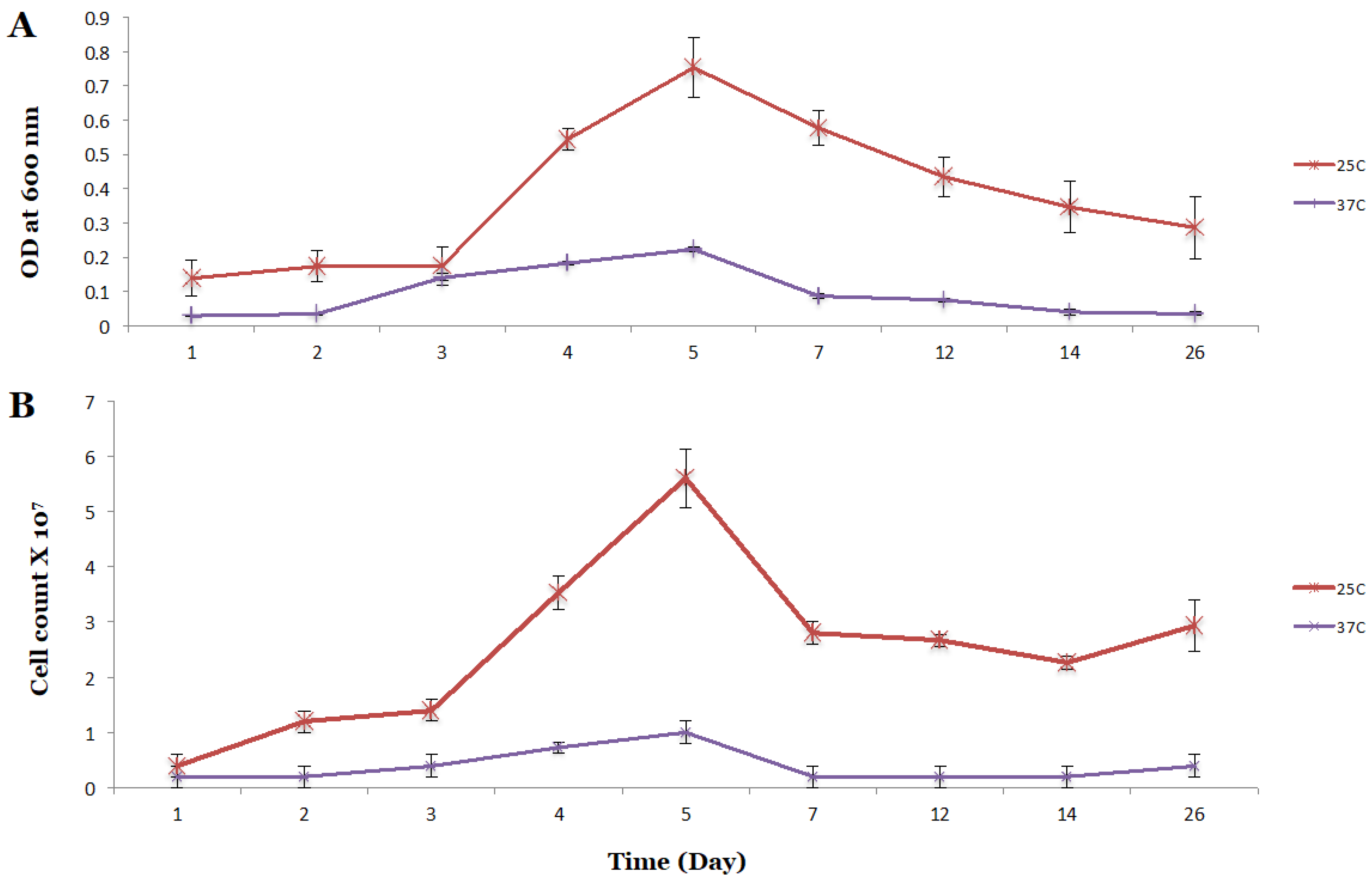
3.1. Cell Growth and Product Formation of the Wild Type P7 Strain
3.2. Development of an EMS Mutagenized Population of C. carboxidivorans P7
3.3. P7-EMS Mutagenized Mutants Showed Significant Increased Levels of Alcohol Production
3.4. P7-EMS Mutants Showed Increased Levels of Atmospheric CO2 Reduction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Availability of data and materials
Ethics and consent to participate
References
- Schlager, S.; Fuchsbauer, A.; Haberbauer, M.; Neugebauer, H.; Sariciftci, N.S. Carbon dioxide conversion to synthetic fuels using biocatalytic electrodes. J. Mater. Chem. A 2017, 5, 2429–2443. [Google Scholar] [CrossRef]
- Leung, D.Y.C.; Caramanna, G.; Maroto-Valer, M.M. An overview of current status of carbon dioxide capture and storage technologies. Renew. Sustain. Energy Rev. 2014, 39, 426–443. [Google Scholar] [CrossRef] [Green Version]
- Stephens, J.C. Growing interest in carbon capture and storage (ccs) for climate change mitigation. Sustain. Sci. Pract. Policy 2006, 2, 4–13. [Google Scholar] [CrossRef]
- Liou, J.S.; Balkwill, D.L.; Drake, G.R.; Tanner, R.S. Clostridium carboxidivorans sp. Nov., a solvent-producing clostridium isolated from an agricultural settling lagoon, and reclassification of the acetogen clostridium scatologenes strain sl1 as Clostridium drakei sp. Nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 2085–2091. [Google Scholar] [CrossRef]
- Ukpong, M.N.; Atiyeh, H.K.; De Lorme, M.J.; Liu, K.; Zhu, X.; Tanner, R.S.; Wilkins, M.R.; Stevenson, B.S. Physiological response of clostridium carboxidivorans during conversion of synthesis gas to solvents in a gas-fed bioreactor. Biotechnol. Bioeng. 2012, 109, 2720–2728. [Google Scholar] [CrossRef]
- Alissandratos, A.; Kim, H.K.; Matthews, H.; Hennessy, J.E.; Philbrook, A.; Easton, C.J. Clostridium carboxidivorans strain p7t recombinant formate dehydrogenase catalyzes reduction of co2 to formate. Appl. Environ. Microbiol. 2013, 79, 741–744. [Google Scholar] [CrossRef] [Green Version]
- Bruant, G.; Levesque, M.J.; Peter, C.; Guiot, S.R.; Masson, L. Genomic analysis of carbon monoxide utilization and butanol production by clostridium carboxidivorans strain p7. PLoS ONE 2010, 5, e13033. [Google Scholar] [CrossRef] [Green Version]
- Paul, D.; Austin, F.W.; Arick, T.; Bridges, S.M.; Burgess, S.C.; Dandass, Y.S.; Lawrence, M.L. Genome sequence of the solvent-producing bacterium clostridium carboxidivorans strain p7t. J. Bacteriol. 2010, 192, 5554–5555. [Google Scholar] [CrossRef] [Green Version]
- Ramio-Pujol, S.; Ganigue, R.; Baneras, L.; Colprim, J. Incubation at 25 degrees c prevents acid crash and enhances alcohol production in clostridium carboxidivorans p7. Bioresour. Technol. 2015, 192, 296–303. [Google Scholar] [CrossRef]
- Li, N.; Yang, J.; Chai, C.; Yang, S.; Jiang, W.; Gu, Y. Complete genome sequence of clostridium carboxidivorans p7(t), a syngas-fermenting bacterium capable of producing long-chain alcohols. J. Biotechnol. 2015, 211, 44–45. [Google Scholar] [CrossRef]
- Pyne, M.E.; Moo-Young, M.; Chung, D.A.; Chou, C.P. Development of an electrotransformation protocol for genetic manipulation of clostridium pasteurianum. Biotechnol. Biofuels 2013, 6, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, C.; Jang, Y.-S.; Moon, H.G.; Lee, J.; Lee, S.Y. Metabolic engineering of clostridia for the production of chemicals. Biofuels Bioprod. Biorefin. 2015, 9, 211–225. [Google Scholar] [CrossRef]
- Wang, J.; Yang, X.; Chen, C.-C.; Yang, S.-T. Engineering clostridia for butanol production from biorenewable resources: From cells to process integration. Curr. Opin. Chem. Eng. 2014, 6, 43–54. [Google Scholar] [CrossRef]
- Huang, H.; Chai, C.; Li, N.; Rowe, P.; Minton, N.P.; Yang, S.; Jiang, W.; Gu, Y. Crispr/cas9-based efficient genome editing in clostridium ljungdahlii, an autotrophic gas-fermenting bacterium. ACS Synth. Biol. 2016. [Google Scholar] [CrossRef]
- Lakhssassi, N.; Zhou, Z.; Liu, S.; Colantonio, V.; AbuGhazaleh, A.; Meksem, K. Characterization of the fad2 gene family in soybean reveals the limitations of gel-based tilling in genes with high copy number. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Lakhssassi, N.; Colantonio, V.; Flowers, N.D.; Zhou, Z.; Henry, J.S.; Liu, S.; Meksem, K. Stearoyl-acyl carrier protein desaturase mutations uncover an impact of stearic acid in leaf and nodule structure. Plant Physiol. 2017. [Google Scholar] [CrossRef] [Green Version]
- Howell-Saxton, E.; Zamenhof, S.; Zamenhof, P.J. Response of escherichia coli to ethyl methanesulfonate: Influence of growth phase and repair ability on survival and mutagenesis. Mutat. Res. 1973, 20, 327–337. [Google Scholar] [CrossRef]
- Lakhssassi, N.; Patil, G.; Piya, S.; Zhou, Z.; Baharlouei, A.; Kassem, M.A.; Lightfoot, D.A.; Hewezi, T.; Barakat, A.; Nguyen, H.T.; et al. Genome reorganization of the gmshmt gene family in soybean showed a lack of functional redundancy in resistance to soybean cyst nematode. Sci. Rep. 2019, 9, 1506. [Google Scholar] [CrossRef] [Green Version]
- Kandoth, P.K.L.; Prenger, S.; Ludwig, E.; Andrew Lakhssassi, N.; Heinz, R.; Zhou, Z.; Howland, A.; Gunther, J.; Eidson, S.; Dhroso, A.; et al. Systematic mutagenesis of serine hydroxymethyltransferase reveals essential role in nematode resistance. Plant Physiol. 2017, in press. [Google Scholar] [CrossRef] [Green Version]
- Ramio-Pujol, S.; Ganigue, R.; Baneras, L.; Colprim, J. Impact of formate on the growth and productivity of clostridium ljungdahlii petc and clostridium carboxidivorans p7 grown on syngas. Int. Microbiol. 2014, 17, 195–204. [Google Scholar]
- Nozzi, N.E.; Desai, S.H.; Case, A.E.; Atsumi, S. Metabolic engineering for higher alcohol production. Metab. Eng. 2014, 25, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Ramió-Pujol, S.; Ganigué, R.; Bañeras, L.; Colprim, J. How can alcohol production be improved in carboxydotrophic clostridia? Process Biochem. 2015, 50, 1047–1055. [Google Scholar] [CrossRef]
- Ma, C.; Kojima, K.; Xu, N.; Mobley, J.; Zhou, L.; Yang, S.T.; Liu, X.M. Comparative proteomics analysis of high n-butanol producing metabolically engineered clostridium tyrobutyricum. J. Biotechnol. 2015, 193, 108–119. [Google Scholar] [CrossRef]
- Phillips, J.R.; Atiyeh, H.K.; Tanner, R.S.; Torres, J.R.; Saxena, J.; Wilkins, M.R.; Huhnke, R.L. Butanol and hexanol production in clostridium carboxidivorans syngas fermentation: Medium development and culture techniques. Bioresour. Technol. 2015, 190, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, T.D.C. Sarah Medium for use in antibiotic susceptibility testing of anaerobic bacteria. Antimicrob. Agents Chemother. 1976, 10, 926–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, J.R.; Huhnke, R.L.; Atiyeh, H.K. Syngas fermentation: A microbial conversion process of gaseous substrates to various products. Fermentation 2017, 3, 28. [Google Scholar] [CrossRef] [Green Version]
- Grzesiuk, E.; Janion, C. Some aspects of ems-induced mutagenesis in Escherichia coli. Mutat. Res. 1993, 297, 313–321. [Google Scholar] [CrossRef]
- Pienkowska, M.; Glickman, B.W.; Ferreira, A.; Anderson, M.; Zielenska, M. Large-scale mutational analysis of ems-induced mutation in the laci gene of escherichia coli. Mutat. Res. 1993, 288, 123–131. [Google Scholar] [CrossRef]
- Verma, D.; Singla, A.; Lal, B.; Sarma, P.M. Conversion of biomass-generated syngas into next-generation liquid transport fuels through microbial intervention: Potential and current status. Curr. Sci. 2016, 110, 329–336. [Google Scholar] [CrossRef] [Green Version]
- Daniell, J.; Köpke, M.; Simpson, S. Commercial biomass syngas fermentation. Energies 2012, 5, 5372–5417. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Formula | 1754-A (Original PETC Medium) | 1754-B (Modified PETC Medium) |
|---|---|---|---|
| Volume (0.5 L) | Volume (0.5 L) | ||
| Resazurin | |||
| Resazurin | C12H6NO4Na | 200 µL | 200 µL |
| Minerals | |||
| Ammonium chloride | NH4Cl | 500 mg | 500 mg |
| Magnesium sulfate | MgSO4·7H2O | 100 mg | 100 mg |
| Potassium chloride | KCl | 50 mg | 50 mg |
| Potassium phosphate | KH2PO4 | 50 mg | 50 mg |
| Sodium chloride | NaCl | 400 mg | 400 mg |
| Calcium chloride | CaCl2·2H2O | 20 mg | Not added |
| Buffers | |||
| HEPES | C8H18N2O4S | Undefined | 1.8 g |
| Carbon source | |||
| Yeast Extract | 500 mg | Not added | |
| Fructose | C6H12O6 | 2.5 g | Not added |
| Sodium Bicarbonate | NaHCO3 | 1 g | 1 g |
| Minerals and Vitamins | |||
| ATCC Minerals | 10 mL | 1 mL | |
| ATCC Vitamins | 10 mL | 1 mL | |
| Cysteine/Sulfide | |||
| L-Cysteine | C3H7NO2S | 2 g | 2 g |
| Sodium sulfide | Na2S·9H2O | 2 g | 2 g |
| End Product | P7 Strains | Average (g/L) | End Product Content Increase | Maximum Production (g/L) | End Product Converted on (g/L Abs) | CO2 Reduction (g/Abs) | CO2 Reduction Increase |
|---|---|---|---|---|---|---|---|
| Ethanol | III-J | 0.77 ± 0.20 * | 3.50X | 0.87 | 4.58 | 0.218 | 8.72 X |
| III-R | 1.03 ± 0.40 * | 4.68X | 1.64 | 4.21 | 0.200 | 8 X | |
| III-P | 0.91 ± 0.58 * | 4.13X | 1.79 | 3.38 | 0.161 | 6.44 X | |
| P7-WT | 0.21 ± 0.01 | _ | 0.22 | 0.40 | 0.025 | _ | |
| Butyrate | III-J | 0.54 ± 0.10** | 3.85X | 0.7 | 3.33 | 0.166 | 8.73 X |
| III-R | 0.45 ± 0.07** | 3.21X | 0.49 | 2.88 | 0.143 | 7.52 X | |
| III-P | 0.27 ± 0.01 * | 1.92X | 0.28 | 1.12 | 0.055 | 2.89 X | |
| P7-WT | 0.13 ± 0.01 | _ | 0.14 | 0.50 | 0.019 | _ | |
| Butanol | III-J | 0.23 ± 0.02 | x | 0.25 | 1.38 | 0.082 | x |
| P7-WT | ND | _ | ND | NA | NA | _ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lakhssassi, N.; Baharlouei, A.; Meksem, J.; Hamilton-Brehm, S.D.; Lightfoot, D.A.; Meksem, K.; Liang, Y. EMS-Induced Mutagenesis of Clostridium carboxidivorans for Increased Atmospheric CO2 Reduction Efficiency and Solvent Production. Microorganisms 2020, 8, 1239. https://doi.org/10.3390/microorganisms8081239
Lakhssassi N, Baharlouei A, Meksem J, Hamilton-Brehm SD, Lightfoot DA, Meksem K, Liang Y. EMS-Induced Mutagenesis of Clostridium carboxidivorans for Increased Atmospheric CO2 Reduction Efficiency and Solvent Production. Microorganisms. 2020; 8(8):1239. https://doi.org/10.3390/microorganisms8081239
Chicago/Turabian StyleLakhssassi, Naoufal, Azam Baharlouei, Jonas Meksem, Scott D. Hamilton-Brehm, David A. Lightfoot, Khalid Meksem, and Yanna Liang. 2020. "EMS-Induced Mutagenesis of Clostridium carboxidivorans for Increased Atmospheric CO2 Reduction Efficiency and Solvent Production" Microorganisms 8, no. 8: 1239. https://doi.org/10.3390/microorganisms8081239