Exploring the Pathogenic Potential of Vibrio vulnificus Isolated from Seafood Harvested along the Mangaluru Coast, India


Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of V. vulnificus
2.2. Biochemical Characterization
2.3. Genotyping and Detection of Virulence Genes of V. vulnificus
2.4. Phylogeny Tree Construction
2.5. Bacteria Strains Used for Virulence Assay
2.6. Serum Resistance
2.7. Adherence and Invasion Assays
2.8. Cytotoxicity Assay
2.9. RNA Extraction and DNase Treatment
2.10. qPCR Assay
2.11. Evaluation of Cell Structural Damage by Immunofluorescence Assay
2.12. Statistical Analysis
3. Results
3.1. Isolation and Phenotypic Characterization of V. vulnificus from Seafood
3.2. Genotyping of V. vulnificus Isolates
3.3. Serum Resistance/Sensitivity Assay
3.4. Adherence Ability of Environmental Strains of V. vulnificus
3.5. Cytotoxic Effect of V. vulnificus on HeLa Cells
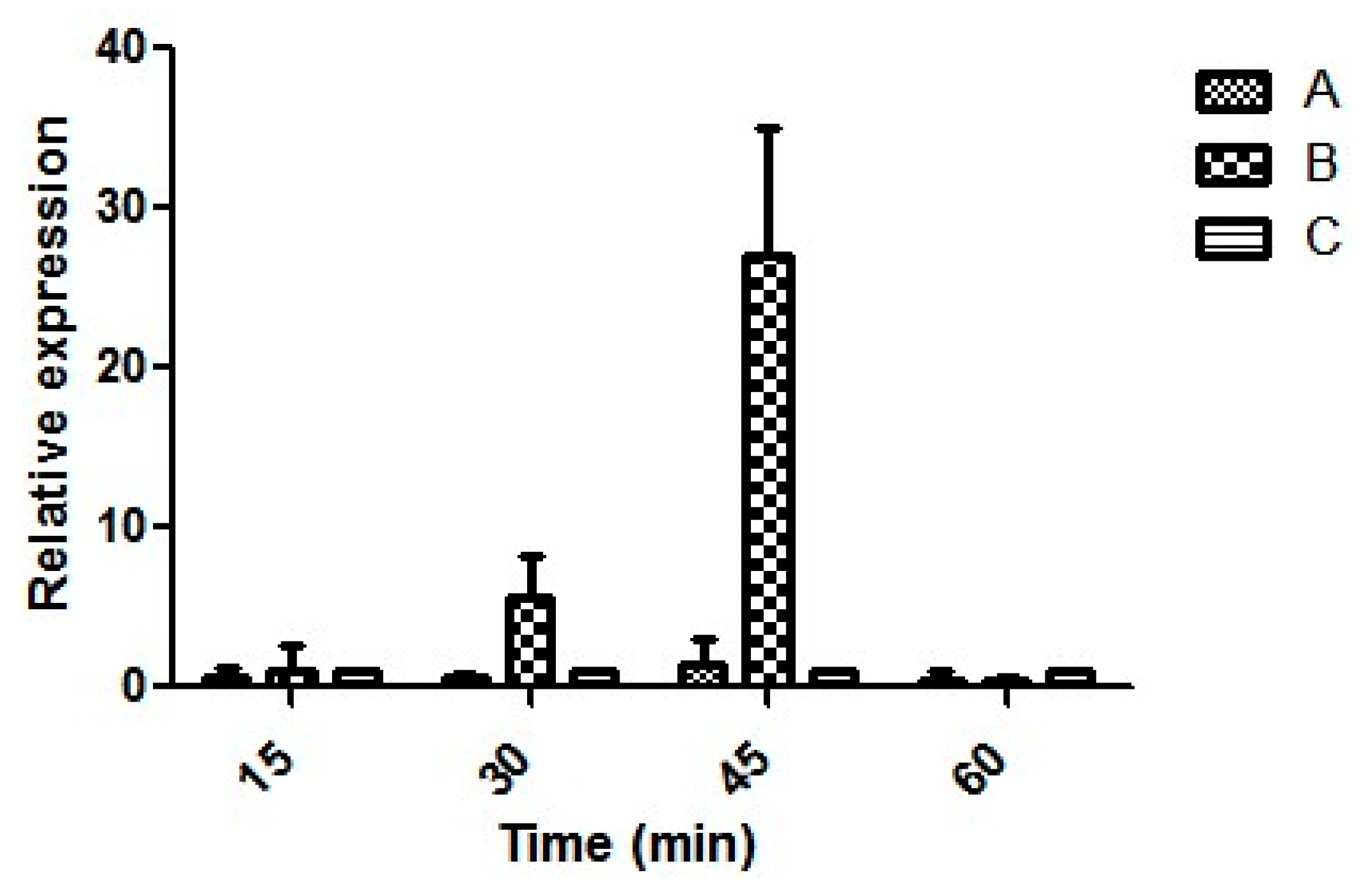
3.6. Expression of Virulence Genes
3.7. Effect of V. vulnificus Infection on HeLa Cell Morphology and Actin Perturbation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heng, S.-P.; Letchumanan, V.; Deng, C.-Y.; Ab Mutalib, N.-S.; Khan, T.M.; Chuah, L.-H.; Chan, K.-G.; Goh, B.-H.; Pusparajah, P.; Lee, L.-H. Vibrio vulnificus: An Environmental and Clinical Burden. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Oliver, J.D. Vibrio vulnificus: New insights into a deadly opportunistic pathogen. Environ. Microbiol. 2018, 20, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.K.; Oliver, J.D. Vibrio vulnificus: Disease and Pathogenesis. Infect. Immun. 2009, 77, 1723–1733. [Google Scholar] [CrossRef] [PubMed]
- Strom, M.S.; Paranjpye, R.N. Epidemiology and pathogenesis of Vibrio vulnificus. Microbes Infect. 2000, 2, 177–188. [Google Scholar] [CrossRef]
- Parvathi, A.; Kumar, H.S.; Karunasagar, I.; Karunasagar, I. Detection and enumeration of Vibrio vulnificus in oysters from two estuaries along the southwest coast of India, using molecular methods. Appl. Environ. Microbiol. 2004, 70, 6909–6913. [Google Scholar] [CrossRef]
- Parvathi, A.; Kumar, H.S.; Karunasagar, I.; Karunasagar, I. Study of the occurrence of Vibrio vulnificus in oysters in India by polymerase chain reaction (PCR) and heterogeneity among V. vulnificus by randomly amplified polymorphic DNA PCR and gyrB sequence analysis. Environ. Microbiol. 2005, 7, 995–1002. [Google Scholar] [CrossRef]
- Kumar, H.S.; Parvathi, A.; Karunasagar, I.; Karunasagar, I. A gyrB-based PCR for the detection of Vibrio vulnificus and its application for direct detection of this pathogen in oyster enrichment broths. Int. J. Food Microbiol. 2006, 111, 216–220. [Google Scholar] [CrossRef]
- Jones, J.L.; Lüdeke, C.H.M.; Bowers, J.C.; DeRosia-Banick, K.; Carey, D.H.; Hastback, W. Abundance of Vibrio cholerae, V. vulnificus, and V. parahaemolyticus in Oysters (Crassostrea virginica) and Clams (Mercenaria mercenaria) from Long Island Sound. Appl. Environ. Microbiol. 2014, 80, 7667–7672. [Google Scholar] [CrossRef]
- De, A.; Mathur, M. Vibrio vulnificus diarrhea in a child with respiratory infection. J. Glob. Infect. Dis. 2011, 3, 310. [Google Scholar] [CrossRef]
- Madiyal, M.; Eshwara, V.; Halim, I.; Stanley, W.; Prabhu, M.; Mukhopadhyay, C. A rare glimpse into the morbid world of necrotising fasciitis: Flesh-eating bacteria Vibrio vulnificus. Indian J. Med. Microbiol. 2016, 34, 384. [Google Scholar] [CrossRef]
- Kimman, T.G.; Smit, E.; Klein, M.R. Evidence-based biosafety: A review of the principles and effectiveness of microbiological containment measures. Clin. Microbiol. Rev. 2008, 21, 403–425. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, W.B.; Paranjype, R.N.; DePaola, A.; Strom, M.S. Sequence Polymorphism of the 16S rRNA Gene of Vibrio vulnificus Is a Possible Indicator of Strain Virulence. J. Clin. Microbiol. 2003, 41, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Rosche, T.M.; Yano, Y.; Oliver, J.D. A rapid and simple PCR analysis indicates there are two subgroups of Vibrio vulnificus which correlate with clinical or environmental isolation. Microbiol. Immunol. 2005, 49, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Chatzidaki-Livanis, M.; Jones, M.K.; Wright, A.C. Genetic Variation in the Vibrio vulnificus Group 1 Capsular Polysaccharide Operon. J. Bacteriol. 2006, 188, 1987–1998. [Google Scholar] [CrossRef]
- Thiaville, P.C.; Bourdage, K.L.; Wright, A.C.; Farrell-Evans, M.; Garvan, C.W.; Gulig, P.A. Genotype Is Correlated with but Does Not Predict Virulence of Vibrio vulnificus Biotype 1 in Subcutaneously Inoculated, Iron Dextran-Treated Mice. Infect. Immun. 2011, 79, 1194–1207. [Google Scholar] [CrossRef]
- Tisont, D.L.; Kellyt, M.T. Virulence of Vibrio vulnificus Strains from Marine Environments. Appl. Environ. Microbiol. 1986, 51, 1004–1006. [Google Scholar] [CrossRef]
- Moreno, M.L.G. Landgraf Virulence factors and pathogenicity of Vibrio vulnificus strains isolated from seafood. J. Appl. Microbiol. 1998, 84, 747–751. [Google Scholar] [CrossRef]
- DePaola, A.; Nordstrom, J.L.; Dalsgaard, A.; Forslund, A.; Oliver, J.; Bates, T.; Bourdage, K.L.; Gulig, P.A. Analysis of Vibrio vulnificus from Market Oysters and Septicemia Cases for Virulence Markers. Appl. Environ. Microbiol. 2003, 69, 4006–4011. [Google Scholar] [CrossRef]
- Wong, H.-C.; Liu, S.-H.; Chen, M.-Y. Virulence and stress susceptibility of clinical and environmental strains of Vibrio vulnificus isolated from samples from Taiwan and the United States. J. Food Prot. 2005, 68, 2533–2540. [Google Scholar] [CrossRef]
- Natividad-Bonifacio, I.; Fernández, F.J.; Quiñones-Ramírez, E.I.; Curiel-Quesada, E.; Vázquez-Salinas, C. Presence of virulence markers in environmental Vibrio vulnificus strains. J. Appl. Microbiol. 2013, 114, 1539–1546. [Google Scholar] [CrossRef]
- Gavin, H.E.; Beubier, N.T.; Satchell, K.J.F. The Effector Domain Region of the Vibrio vulnificus MARTX Toxin Confers Biphasic Epithelial Barrier Disruption and Is Essential for Systemic Spread from the Intestine. PLoS Pathog. 2017, 13, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.E.; Calia, F.M.; Musher, D.M.; Goree, A. Resistance of Vibrio vulnificus to Serum Bactericidal and Opsonizing Factors: Relation to Virulence in Suckling Mice and Humans. J. Infect. Dis. 1984, 150, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Panicker, G.; Bej, A.K. Rapid detection of Vibrio vulnificus in Gulf water and shellfish using real-time PCR. Appl. Environ. Microbiol. 2004, 70, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.C.; Simpson, L.M.; Oliver, J.D. Role of Iron in the Pathogenesis of Vibrio vulnificus Infections. Infect. Immun. 1981, 34, 503–507. [Google Scholar] [CrossRef]
- Han, F.; Pu, S.; Hou, A.; Ge, B. Characterization of Clinical and Environmental Types of Vibrio vulnificus Isolates from Louisiana Oysters. Foodborne Pathog. Dis. 2009, 6, 1251–1258. [Google Scholar] [CrossRef]
- Panicker, G.; Vickery, M.C.; Bej, A.K. Multiplex PCR detection of clinical and environmental strains of Vibrio vulnificus in shellfish. Can. J. Microbiol. 2004, 50, 911–922. [Google Scholar] [CrossRef]
- Bogard, R.W.; Oliver, J.D. Role of Iron in Human Serum Resistance of the Clinical and Environmental Vibrio vulnificus Genotypes. Appl. Environ. Microbiol. 2007, 73, 7501–7505. [Google Scholar] [CrossRef]
- Yokochi, N.; Tanaka, S.; Matsumoto, K.; Oishi, H.; Tashiro, Y.; Yoshikane, Y.; Nakashima, M.; Kanda, K.; Kobayashi, G. Distribution of Virulence Markers among Vibrio vulnificus Isolates of Clinical and Environmental Origin and Regional Characteristics in Japan. PLoS ONE 2013, 8, 14–16. [Google Scholar] [CrossRef]
- Wright, A.C.; Simpson, L.M.; Oliver, J.D.; Morris, J.G. Phenotypic evaluation of acapsular transposon mutants of Vibrio vulnificus. Infect. Immun. 1990, 58, 1769–1773. [Google Scholar] [CrossRef]
- Hwang, I.; Kim, B.; Lee, K.S.; Kim, I.; Son, J.S.; Kim, K. Identification of Virulence Factors in Vibrio vulnificus by Comparative Transcriptomic Analyses between Clinical and Environmental Isolates Using cDNA Microarray. J. Microbiol. Biotechnol. 2011, 21, 1228–1235. [Google Scholar] [CrossRef]
- Paranjpye, R.N.; Lara, J.C.; Pepe, J.C.; Pepe, C.M.; Strom, M.S. The Type IV Leader Peptidase/N -Methyltransferase of Vibrio vulnificus Controls Factors Required for Adherence to HEp-2 Cells and Virulence in Iron-Overloaded Mice. Infect. Immun. 1998, 66, 5659–5668. [Google Scholar] [CrossRef] [PubMed]
- Paranjpye, R.N.; Strom, M.S. A Vibrio vulnificus Type IV Pilin Contributes to Biofilm Formation, Adherence to Epithelial Cells, and Virulence. Infect. Immun. 2005, 73, 1411–1422. [Google Scholar] [CrossRef] [PubMed]
- Garrett, S.B.; Garrison-Schilling, K.L.; Cooke, J.T.; Pettis, G.S. Capsular polysaccharide production and serum survival of Vibrio vulnificus are dependent on antitermination control by RfaH. FEBS Lett. 2016, 590, 4564–4572. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.; Ayrapetyan, M.; Ryan, H.; Oliver, J. Serum Survival of Vibrio vulnificus: Role of Genotype, Capsule, Complement, Clinical Origin, and in Situ Incubation. Pathogens 2014, 3, 822–832. [Google Scholar] [CrossRef] [PubMed]
- Saraswathi, K.; Barve, S.M.; Deodhar, L.P. Septicaemia due to Vibrio vulnificus. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 714. [Google Scholar] [CrossRef]
- D’Souza, C.; Kumar, B.K.; Kapinakadu, S.; Shetty, R.; Karunasagar, I.; Karunasagar, I. PCR-based evidence showing the presence of Vibrio vulnificus in wound infection cases in Mangaluru, India. Int. J. Infect. Dis. 2018, 68, 74–76. [Google Scholar] [CrossRef]
- Bhat, P.; Bhaskar, M.; Sistla, S.; Kadhiravan, T. Fatal case of necrotising fasciitis due to Vibrio vulnificus in a patient with alcoholic liver disease and diabetes mellitus. BMJ Case Rep. 2019, 12. [Google Scholar] [CrossRef]
- Thampuran, N.; Surendran, P.K. Occurrence and distribution of Vibrio vulnificus in tropical fish and shellfish from Cochin (India). Lett. Appl. Microbiol. 1998, 26, 110–112. [Google Scholar] [CrossRef]
- Karunasagar, I.; Rohit, A. Epidemiology and Molecular Pathogenesis of Vibrio vulnificus. Foodborne Waterborne Bact. Pathog. Epidemiol. Evol. Mol. Biol. 2012, 197–209. [Google Scholar] [CrossRef]
- Kaysner, C.A.; DePaola, J. Bacteriological Analytical Manual Chapter 9: Vibrio; Food US and Drug Administration: Silver Spring, MD, USA, 2004; Volume 40, pp. 1–15. [Google Scholar]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Ausubel, F.M.; Brent, R.; Kingston, R.E.; Moore, D.D.; Seidman, J.G.; Smith, J.A.; Struhl, K. Current Protocols in Molecular Biology, 3rd ed.; John Wiley & Sons Inc. Media: Hoboken, NJ, USA, 1989. [Google Scholar] [CrossRef]
- Acharya, S.B. Quantification of Human Pathogenic Vibrios in Seafood Using Molecular Techniques Karnataka Veterinary, Animal and Fisheries Sciences University. 2013. Available online: http://hdl.handle.net/10603/7012 (accessed on 21 February 2013).
- Lee, S.E.; Kim, S.Y.; Kim, S.J.; Kim, H.S.; Shin, J.H.; Choi, S.H.; Chung, S.S.; Rhee, J.H. Direct Identification of Vibrio vulnificus in Clinical Specimens by Nested PCR. J. Clin. Microbiol. 1998, 36, 2887–2892. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Biosca, E.G.; Oliver, J.D.; Amaro, C. Phenotypic characterization of Vibrio vulnificus biotype 2, a lipopolysaccharide-based homogeneous O serogroup within Vibrio vulnificus. Appl. Environ. Microbiol. 1996, 62, 918–927. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, C.; Alva, P.P.; Padyana, A.K.; Karunasagar, I.; Karunasagar, I.; Kumar, B.K.; Ballamoole, K.K. Unveiling the acid stress response of clinical genotype Vibrio vulnificus isolated from the marine environments of Mangaluru coast, India. Can. J. Microbiol. 2019, 65, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Reynaud, Y.; Pitchford, S.; Decker, S.D.; Wikfors, G.H.; Brown, C.L. Molecular Typing of Environmental and Clinical Strains of Vibrio vulnificus Isolated in the Northeastern USA. PLoS One. 2013, 8, e83357. [Google Scholar] [CrossRef]
- Sangeetha, M.S.; Shekar, M.; Venugopal, M.N. Occurrence of clinical genotype Vibrio vulnificus in clam samples in Mangalore, Southwest coast of India. J. Food Sci. Technol. 2017, 54, 786–791. [Google Scholar] [CrossRef]
- Kim, Y.R.; Lee, S.E.; Kook, H.; Yeom, J.A.; Na, H.S.; Kim, S.Y.; Chung, S.S.; Choy, H.E.; Rhee, J.H. Vibrio vulnificus RTX toxin kills host cells only after contact of the bacteria with host cells. Cell. Microbiol. 2008, 10, 848–862. [Google Scholar] [CrossRef]
- Al-assafi, M.M.K.; Mutalib, S.A.; Ghani, A. A Review of Important Virulence Factors of Vibrio vulnificus. Curr. Res. J. Biol. Sci. 2016. [Google Scholar] [CrossRef]
- Kim, B.S.; Gavin, H.E.; Satchell, K.J.F. Distinct Roles of the Repeat-Containing Regions and Effector Domains of the Vibrio vulnificus Multifunctional-Autoprocessing Repeats-in-Toxin (MARTX) Toxin. MBio. 2015, 6, 1–11. [Google Scholar] [CrossRef]
- Chen, C.-L.; Chien, S.-C.; Leu, T.-H.; Harn, H.I.-C.; Tang, M.; Hor, L.-I. Vibrio vulnificus MARTX cytotoxin causes inactivation of phagocytosis-related signaling molecules in macrophages. J. Biomed. Sci. 2017, 24, 58. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, S.; Paranjpye, R.N.; Dykhuizen, D.E.; Sokurenko, E.V.; Strom, M.S. Comparative Evolutionary Analysis of the Major Structural Subunit of Vibrio vulnificus Type IV Pili. Mol. Boil. Evol. 2009, 26, 2185–2196. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, S.I.; Ogawa, M.; Mizuguchi, Y. Relation of capsular materials and colony opacity to virulence of Vibrio vulnificus. Infect. Immun. 1985, 47, 446–451. [Google Scholar] [CrossRef] [PubMed]


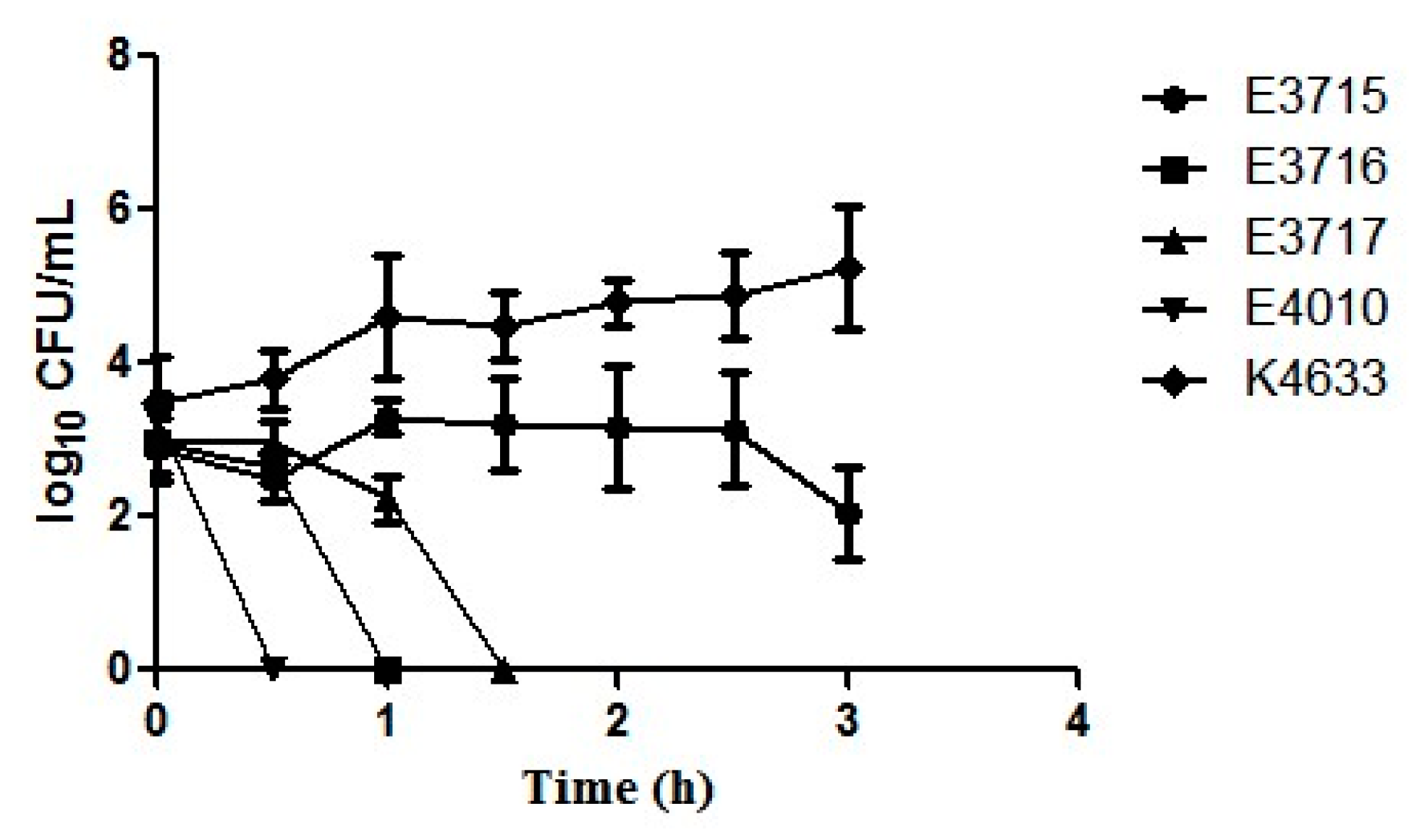
 ), E3716 (
), E3716 ( ), E3717 (
), E3717 ( ), E4010 (
), E4010 ( ), and a clinical strain K4633 (
), and a clinical strain K4633 ( ) exposed to serum for 3 h. Error bars represent the standard error of the mean of three replicates each.
), E3716 (), E3717 (), E4010 (), and a clinical strain K4633 () exposed to serum for 3 h. Error bars represent the standard error of the mean of three replicates each.
) exposed to serum for 3 h. Error bars represent the standard error of the mean of three replicates each.
), E3716 (), E3717 (), E4010 (), and a clinical strain K4633 () exposed to serum for 3 h. Error bars represent the standard error of the mean of three replicates each.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Percentage of Adherence | ||
|---|---|---|---|
| 0.5 h | 1 h | 2 h | |
| E3715 | 8 ± 1.5 | 11.3 ± 3.05 | >100 |
| E3716 | 10.1 ± 2.1 | 12 ± 2.5 | >100 |
| E3717 | 6.5 ± 1.6 | 17 ± 1.4 | 50–100 |
| E4010 | 17 ± 2.8 | 16.5 ± 3.5 | >100 |
| K4633 | 17.3 ± 3.05 | 23.6 ± 4.8 | >100 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Souza, C.; Prithvisagar, K.S.; Deekshit, V.K.; Karunasagar, I.; Karunasagar, I.; Kumar, B.K. Exploring the Pathogenic Potential of Vibrio vulnificus Isolated from Seafood Harvested along the Mangaluru Coast, India. Microorganisms 2020, 8, 999. https://doi.org/10.3390/microorganisms8070999
D’Souza C, Prithvisagar KS, Deekshit VK, Karunasagar I, Karunasagar I, Kumar BK. Exploring the Pathogenic Potential of Vibrio vulnificus Isolated from Seafood Harvested along the Mangaluru Coast, India. Microorganisms. 2020; 8(7):999. https://doi.org/10.3390/microorganisms8070999
Chicago/Turabian StyleD’Souza, Caroline, Kattapuni Suresh Prithvisagar, Vijay Kumar Deekshit, Indrani Karunasagar, Iddya Karunasagar, and Ballamoole Krishna Kumar. 2020. "Exploring the Pathogenic Potential of Vibrio vulnificus Isolated from Seafood Harvested along the Mangaluru Coast, India" Microorganisms 8, no. 7: 999. https://doi.org/10.3390/microorganisms8070999
APA StyleD’Souza, C., Prithvisagar, K. S., Deekshit, V. K., Karunasagar, I., Karunasagar, I., & Kumar, B. K. (2020). Exploring the Pathogenic Potential of Vibrio vulnificus Isolated from Seafood Harvested along the Mangaluru Coast, India. Microorganisms, 8(7), 999. https://doi.org/10.3390/microorganisms8070999