Phenotypic Response of Wolbachia pipientis in a Cell-Free Medium


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Trypan Blue Staining and Hemocytometer Aa23 Cell Counts
2.3. Confirmation of Wolbachia Infection Status Using PCR
2.4. Fluorescent In-Situ Hybridization
2.5. Isolation of Extracellular Wolbachia
2.6. Epifluorescence Microscopy
2.7. Phenotypic Microarray Assays
2.8. Quantitative Real-Time PCR Analysis
2.9. Wolbachia Cell Counts
2.10. Statistical Analyses
3. Results
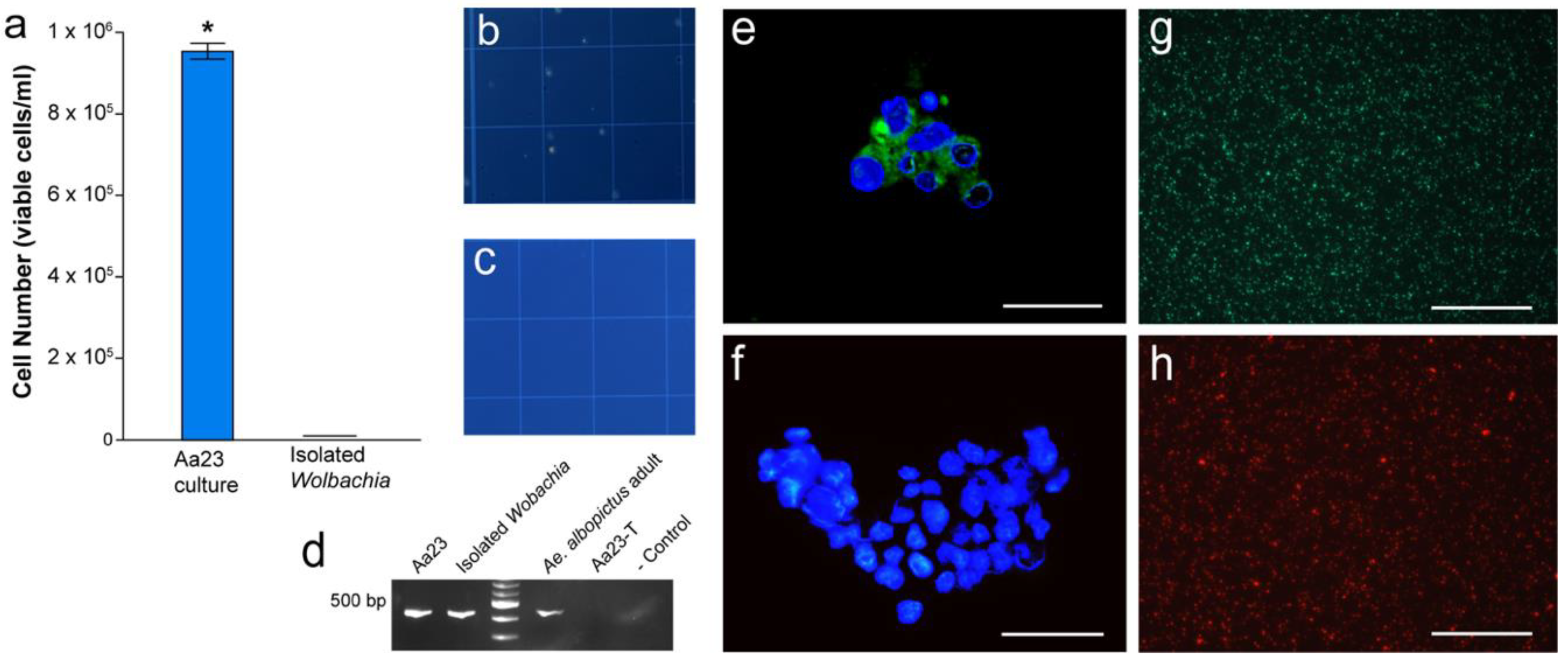
3.1. Confirmation of Wolbachia Infections and Isolation of Wolbachia
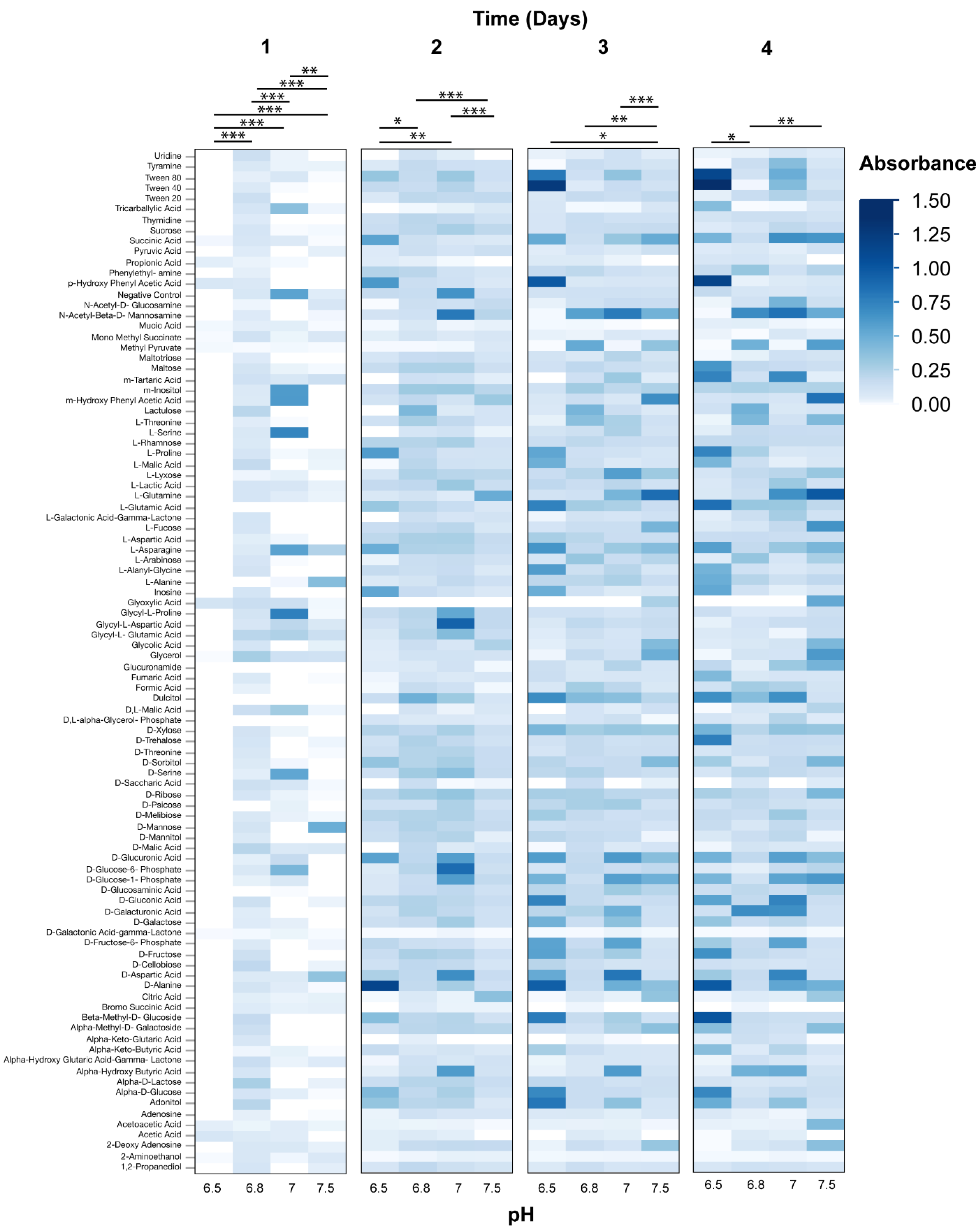
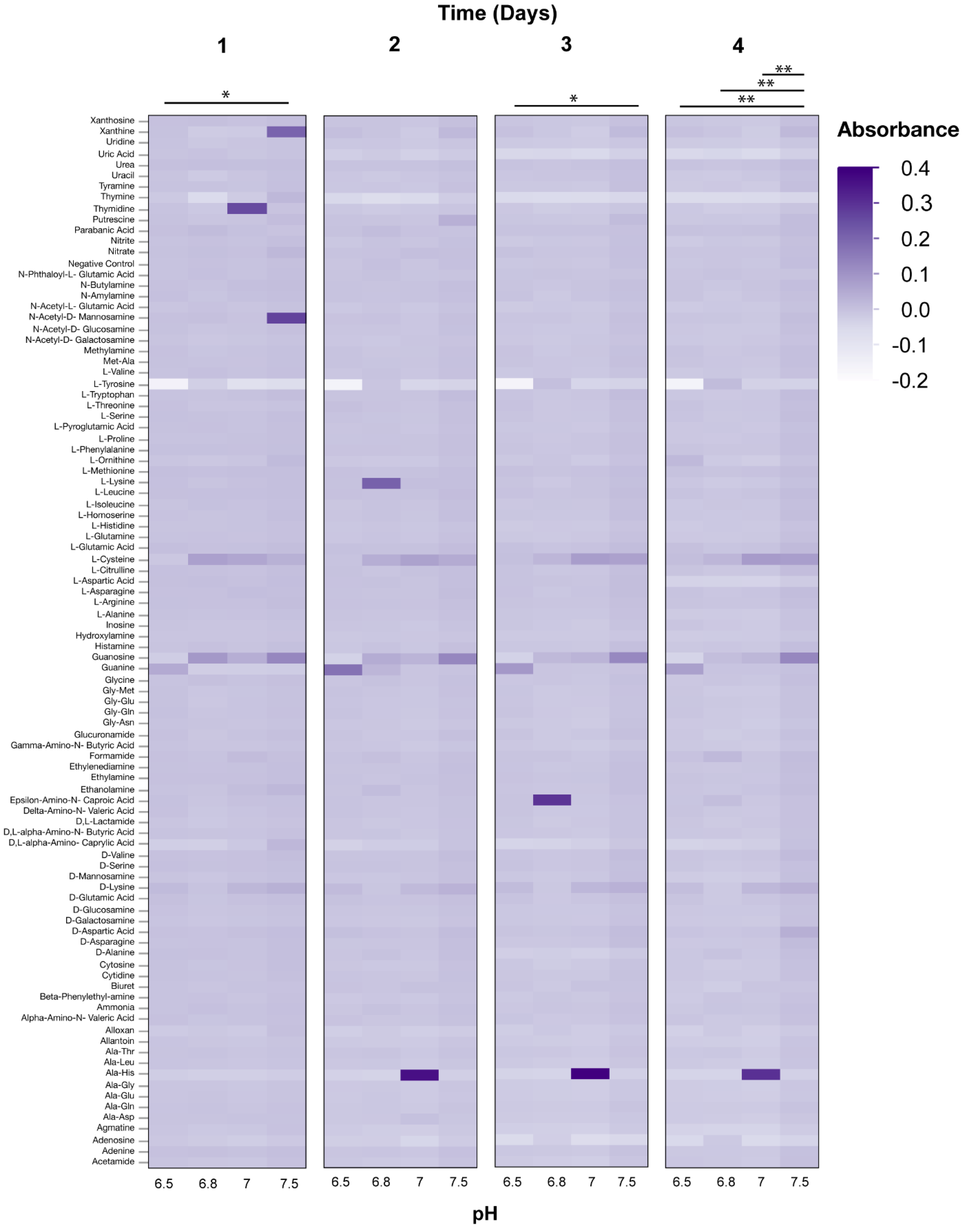
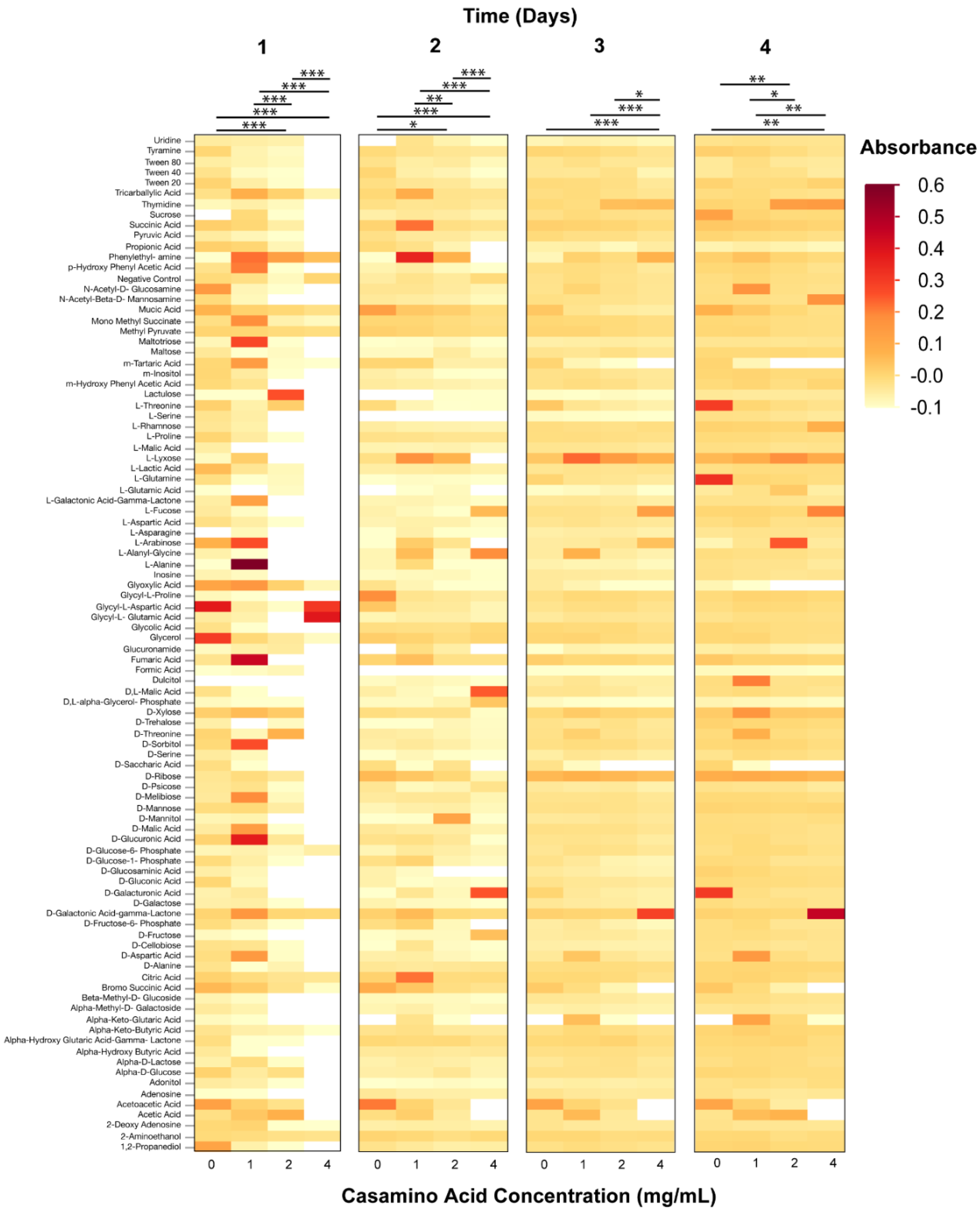
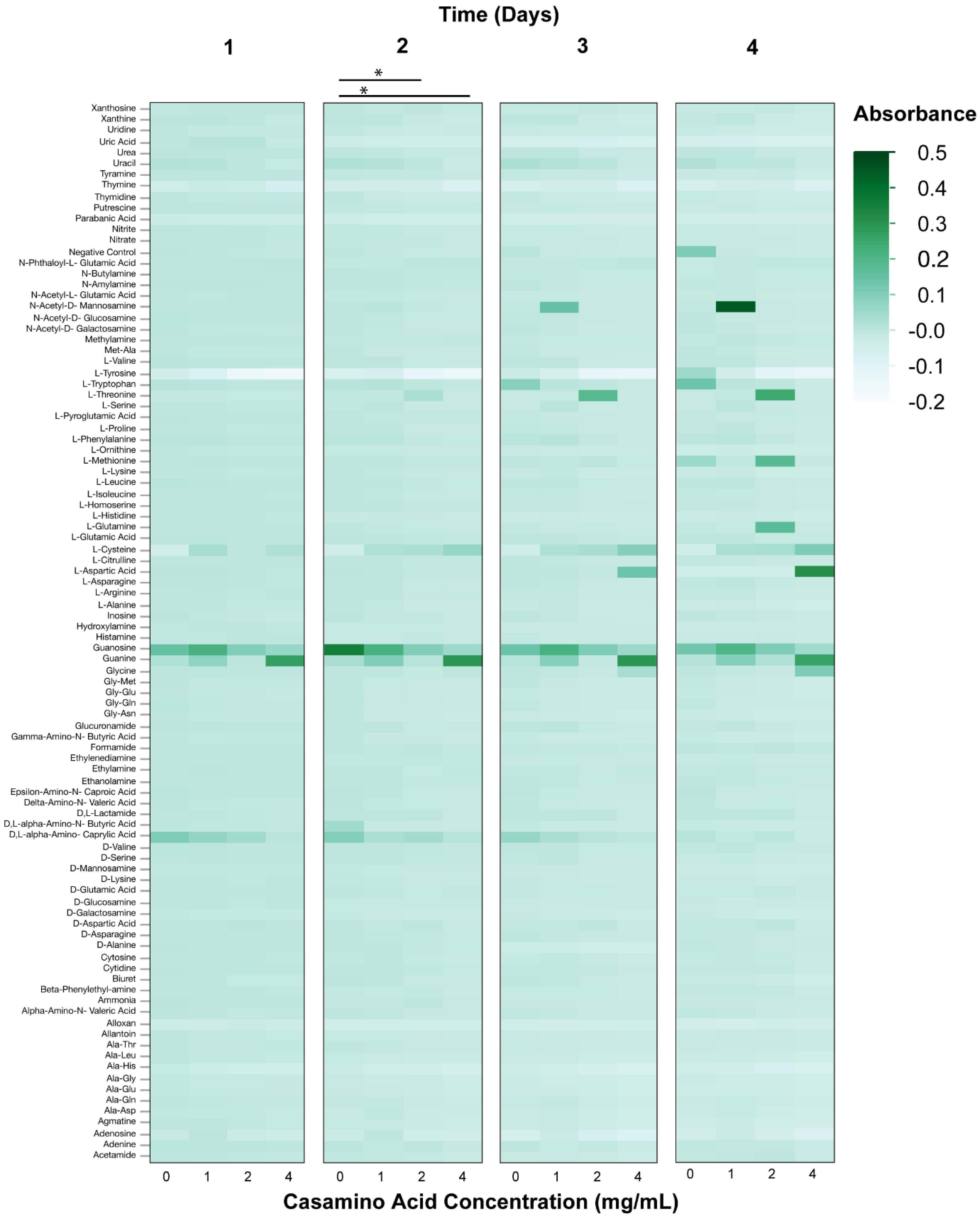
3.2. Phenotypic Microarray Assays
3.3. Quantitative Real-Time PCR
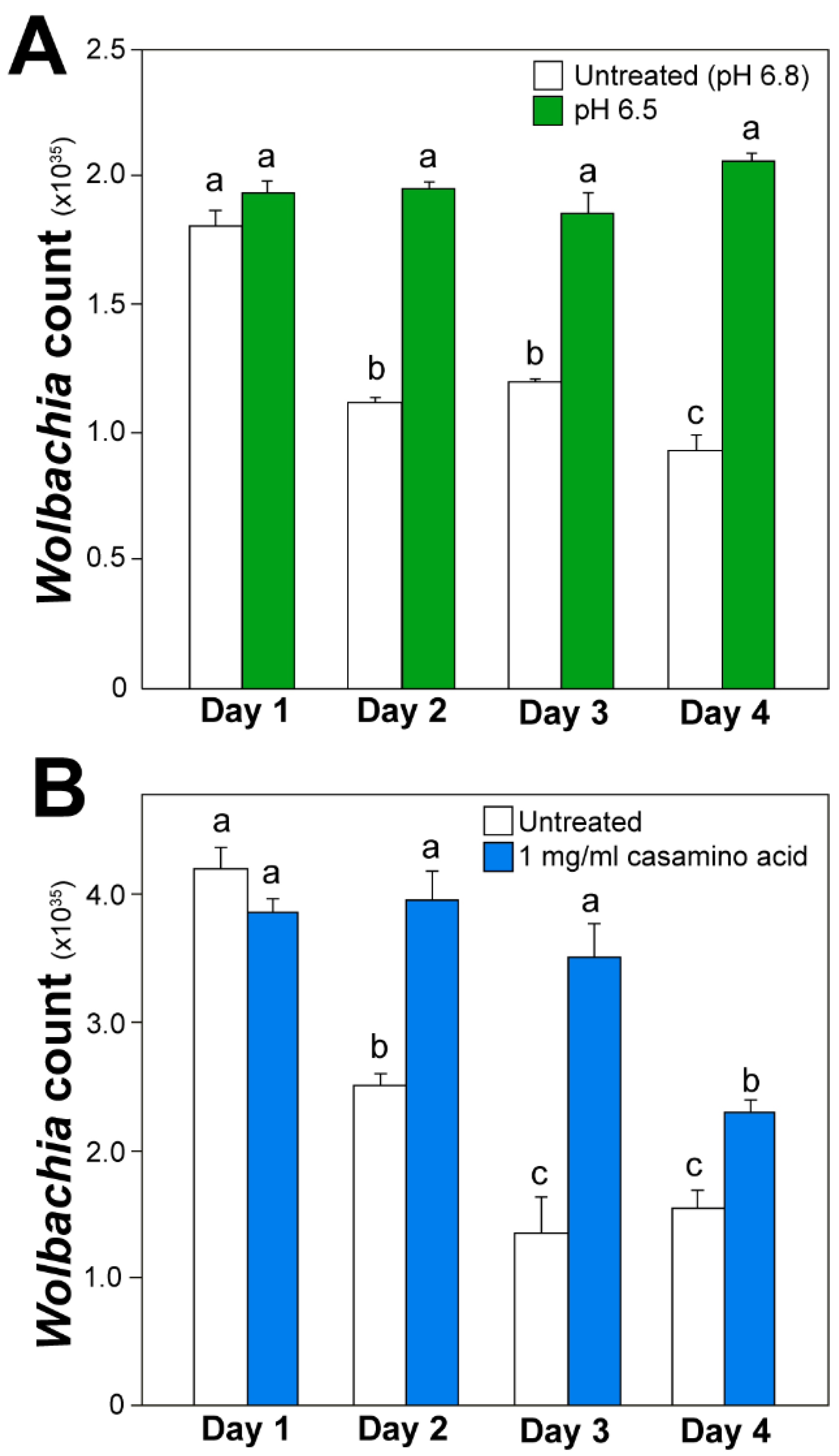
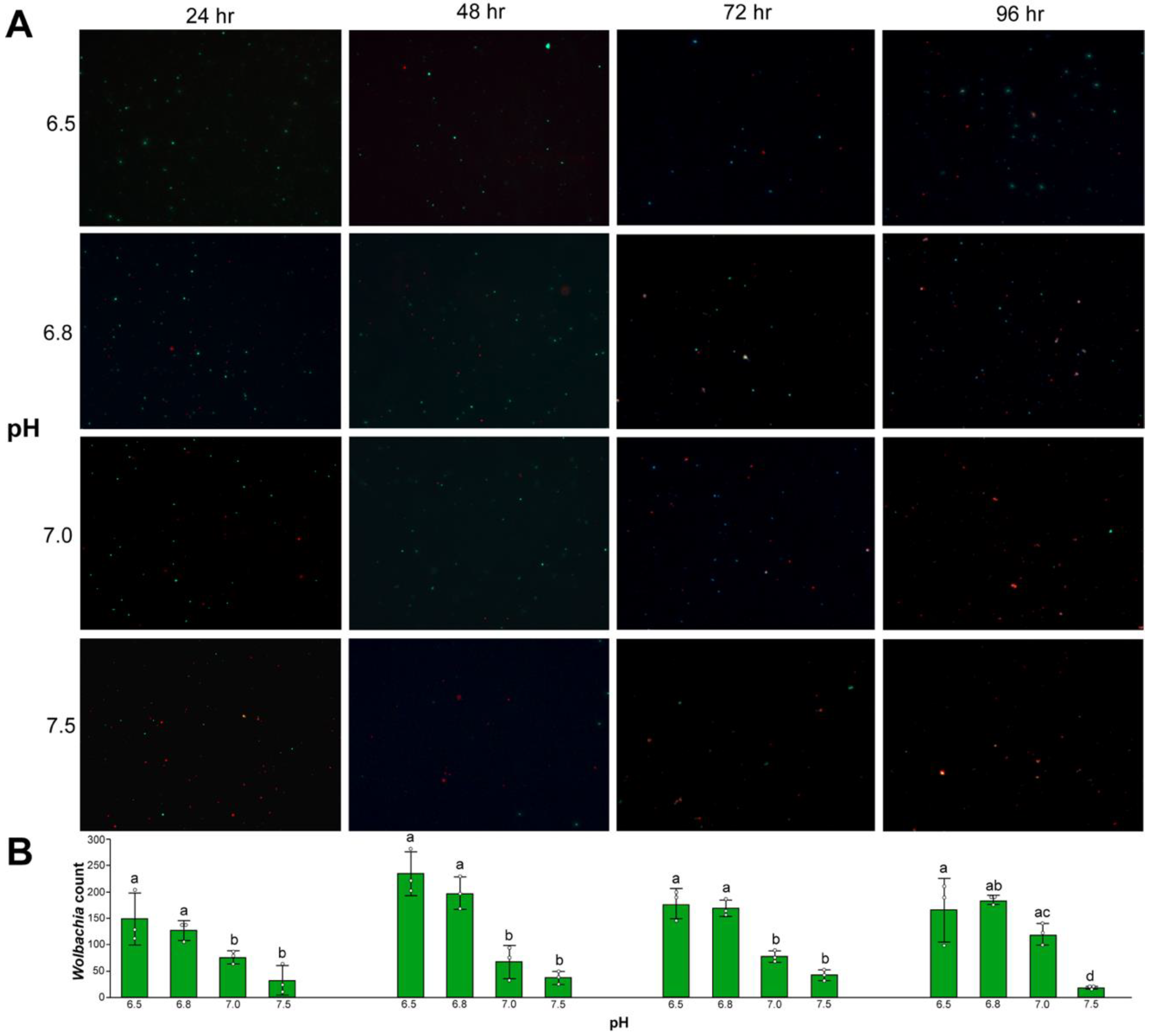
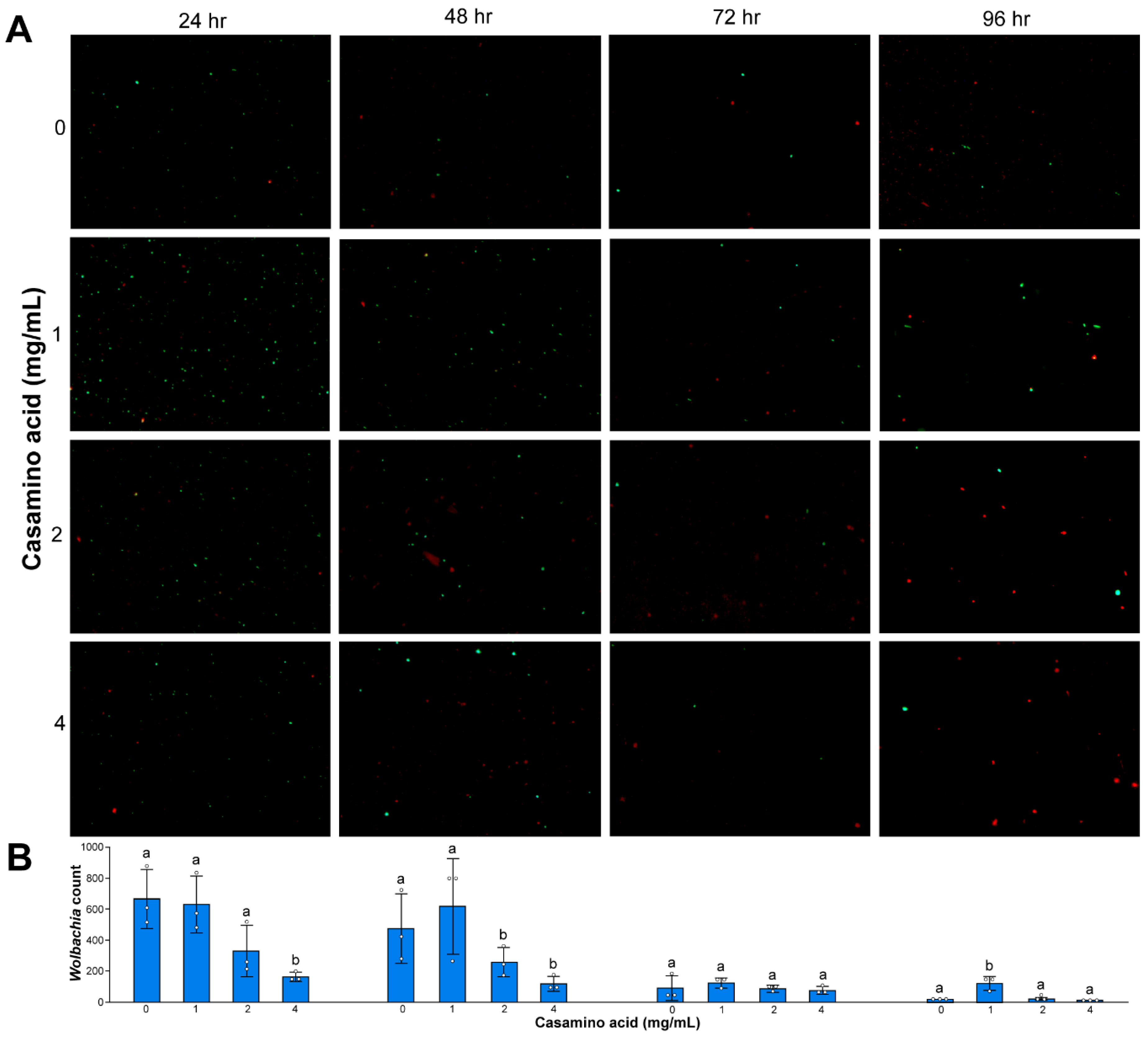
3.4. Wolbachia Cell Counts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Werren, J.H.; Windsor, D.M. Wolbachia infection frequencies in insects: Evidence of a global equilibrium? Proc. Biol. Sci. 2000, 267, 1277–1285. [Google Scholar] [CrossRef]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia?—A statistical analysis of current data. Fems. Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef]
- Hurk, v.d.A.F.; Hall-Mendelin, S.; Pyke, A.T.; Frentiu, F.D.; McElroy, K.; Day, A.; Higgs, S.; O’Neill, S.L. Impact of Wolbachia on infection with chikungunya and yellow fever viruses in the mosquito vector Aedes aegypti. PLoS Negl. Trop. Dis. 2012, 6, e1892. [Google Scholar]
- Caragata, E.P.; Dutra, H.L.C.; Moreira, L.A. Inhibition of Zika virus by Wolbachia in Aedes aegypti. Microb. Cell 2016, 3, 293–295. [Google Scholar] [CrossRef]
- Tan, C.H.; Wong, P.J.; Li, M.I.; Yang, H.T.; Ng, L.C.; O’Neill, S.L. wMel limits zika and chikungunya virus infection in a Singapore Wolbachia-introgressed Ae. aegypti strain, wMel-Sg. PLoS. Negl. Trop. Dis. 2017, 11, e0005496. [Google Scholar] [CrossRef]
- Flores, H.A.; O’Neill, S.L. Controlling vector-borne diseases by releasing modified mosquitoes. Nat. Rev. Microbiol. 2018, 16, 508–518. [Google Scholar] [CrossRef]
- Mains, J.W.; Brelsfoard, C.L.; Rose, R.I.; Dobson, S.L. Femal adult Aedes albopictus suppression by Wolbachia-infected male mosquitoes. Sci. Rep. 2016, 6, 33846. [Google Scholar] [CrossRef]
- Aultman, K.S.; Beaty, B.J.; Walker, E.D. Genetically manipulated vectors of human disease: A practical overview. Trends Para. 2001, 17, 507–509. [Google Scholar] [CrossRef]
- Aksoy, S.; Weiss, B.; Attardo, G. Paratransgenesis applied for control of tsetse transmitted sleeping sickness. In Transgenesis and the Management of Vector-Borne Disease; Springer: New York, NY, USA, 2008; Volume 627, pp. 35–48. [Google Scholar]
- Bourtzis, K. Wolbachia-based technologies for insect pest population control. Adv. Exp. Med. Biol. 2008, 627, 104–113. [Google Scholar] [PubMed]
- Rasgon, J.L.; Gamston, C.E.; Xiaoxia, R. Survival of Wolbachia pipientis in cell-free medium. Appl. Env. Micro. 2006, 72, 6934–6937. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hughes, G.L.; Koga, R.; Xue, P.; Fukatsu, T.; Rasgon, J.L. Wolbachia infections are virulent and inhibit the human malaria parasite Plasmodium falciparum in Anopheles gambiae. PLoS Pathog. 2011, 7, e1002043. [Google Scholar] [CrossRef] [PubMed]
- Dobson, S.L.; Bourtzis, K.; Braig, H.R.; Jones, B.F.; Zhou, W.; Rousset, F.; O’Neill, S.L. Wolbachia infections are distributed throughout insect somatic and germ line tissues. Insect Biochem. Mol. Biol. 1999, 29, 153–160. [Google Scholar] [CrossRef]
- Frydman, H.M.; Li, J.M.; Robson, D.N.; Wieschaus, E. Somatic stem cell niche tropism in Wolbachia. Nature 2006, 441, 509–512. [Google Scholar] [CrossRef]
- Gamston, C.; Rasgon, J. Maintaining Wolbachia in cell-free medium. J. Vis. Exp. 2007, 233. [Google Scholar] [CrossRef]
- Wu, M.; Sun, L.V.; Vamathevan, J.; Riegler, M.; Deboy, R.; Brownlie, J.C.; McGraw, E.A.; Martin, W.; Esser, C.; Ahmadinejad, N.; et al. Phylogenomics of the reproductive parasite Wolbachia pipientis wMel: A streamlined genome overrun by mobile genetic elements. PLoS Bio. 2004, 2, E69. [Google Scholar] [CrossRef]
- Brownlie, J.C.; O’Neill, S.L. Wolbachia genomes: Insights into an intracellular lifestyle. Curr. Biol. 2005, 15, R507–R509. [Google Scholar] [CrossRef]
- Klasson, L.; Westberg, J.; Sapountzis, P.; Naslund, K.; Lutnaes, Y.; Darby, A.C.; Veneti, Z.; Chen, L.; Braig, H.R.; Garrett, R.; et al. The mosaic genome structure of the Wolbachia wRi strain infecting Drosophila simulans. Proc. Natl. Acad. Sci. USA 2009, 106, 5725–5730. [Google Scholar] [CrossRef]
- Maren Ellegaard, K.; Klasson, L.; Naslund, K.; Bourtzis, K.; Anderson, S. Comparative genomics of Wolbachia and the bacterial species concept. PLoS Genet. 2013, 9, e1003381. [Google Scholar] [CrossRef]
- White, P.M.; Serbus, L.R.; Debec, A.; Codina, A.; Bray, W.; Guichet, A.; Lokey, R.S.; Sullivan, W. Reliance of Wolbachia on High Rates of Host Proteolysis Revealed by a Genome-Wide RNAi Screen of Drosophila Cells. Genetics 2017, 205, 1473–1488. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.; Ganatra, M.; Kamal, I.; Ware, J.; Makarova, K.; Ivanova, N.; Bhattacharyya, A.; Kapatral, V.; Kumar, S.; Posfai, J.; et al. The Wolbachia genome of Brugia malayi: Endosymbiont evolution within a human pathogenic nematode. PLoS Biol. 2005, 3, e121. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, T.P.; Verhoeve, V.I.; Guillotte, M.L.; Lehman, S.S.; Rennoll, S.A.; Beier-Sexton, M.; Rahman, M.S.; Azad, A.F.; Gillespie, J.J. Wholly Rickettsia! Reconstructed Metabolic Profile of the Quintessential Bacterial Parasite of Eukaryotic Cells. MBio 2017, 8, e00859-17. [Google Scholar] [CrossRef] [PubMed]
- Marmion, B.P.; Storm, P.A.; Ayres, J.G.; Semendric, L.; Mathews, L.; Winslow, W.; Turra, M.; Harris, R.J. Long-term persistence of Coxiella burnetii after acute primary Q fever. QJM 2005, 98, 7–20. [Google Scholar] [CrossRef]
- Vlak, J.M.; de Gooijer, C.D.; Tramper, J.; Miltenburger, H.G. Insect Cell Cultures: Fundamental and Applied Aspects; Springer: Dordrecht, The Netherlands, 1996. [Google Scholar]
- Medina, M.; Lopez-Rivas, A.; Zuidema, D.; Belsham, G.J.; Domingo, E.; Vlak, J.M. Strong buffering capacity of insect cells. Implications for the baculovirus expression system. Cytotechnology 1995, 17, 21–26. [Google Scholar] [CrossRef]
- Beare, P.A.; Jeffrey, B.M.; Long, C.M.; Martens, C.M.; Heinzen, R.A. Genetic mechanisms of Coxiella burnetii lipopolysaccharide phase variation. PLoS Pathog. 2018, 14, e1006922. [Google Scholar] [CrossRef]
- Beare, P.A.; Larson, C.L.; Gilk, S.D.; Heinzen, R.A. Two systems for targeted gene deletion in Coxiella burnetii. Appl. Env. Microbiol. 2012, 78, 4580–4589. [Google Scholar] [CrossRef]
- Clay, K.A.; Hartley, M.G.; Russell, P.; Norville, I.H. Use of axenic media to determine antibiotic efficacy against Coxiella burnetii. Int. J. Antimicrob. Agents 2018, 51, 806–808. [Google Scholar] [CrossRef]
- Omsland, A.; Cockrell, D.C.; Fischer, E.R.; Heinzen, R.A. Sustained axenic metabolic activity by the obligate intracellular bacterium Coxiella burnetii. J. Bacteriol. 2008, 190, 3203–3212. [Google Scholar] [CrossRef][Green Version]
- Sanchez, S.E.; Vallejo-Esquerra, E.; Omsland, A. Use of Axenic Culture Tools to Study Coxiella burnetii. Curr. Protoc. Microbiol. 2018, 50, e52. [Google Scholar] [CrossRef]
- Sandoz, K.M.; Popham, D.L.; Beare, P.A.; Sturdevant, D.E.; Hansen, B.; Nair, V.; Heinzen, R.A. Transcriptional Profiling of Coxiella burnetii Reveals Extensive Cell Wall Remodeling in the Small Cell Variant Developmental Form. PLoS ONE 2016, 11, e0149957. [Google Scholar] [CrossRef] [PubMed]
- Pontes, M.H.; Dale, C. Culture and manipulation of insect facultative symbionts. Trends Microbiol. 2006, 14, 406–412. [Google Scholar] [CrossRef]
- Omsland, A.; Cockrell, D.C.; Howe, D.; Fischer, E.R.; Virtaneva, K.; Sturdevant, D.E.; Porcella, S.F.; Heinzen, R.A. Host cell-free growth of the Q fever bacterium Coxiella burnetii. Proc. Natl. Acad. Sci. USA 2009, 106, 4430–4434. [Google Scholar] [CrossRef]
- Omsland, A.; Hackstadt, T.; Heinzen, R.A. Bringing culture to the uncultured: Coxiella burnetii and lessons for obligate intracellular bacterial pathogens. PLoS Pathog. 2013, 9, e1003540. [Google Scholar] [CrossRef] [PubMed]
- Brandt, J.W.; Chevignon, G.; Oliver, K.M.; Strand, M.R. Culture of an aphid heritable symbiont demonstrates its direct role in defence against parasitoids. Proc. Biol. Sci. 2017, 284, 20171925. [Google Scholar] [CrossRef] [PubMed]
- Matthew, C.Z.; Darby, A.C.; Young, S.A.; Hume, L.H.; Welburn, S.C. The rapid isolation and growth dynamics of the tsetse symbiont Sodalis glossinidius. Fems. Microbiol. Lett. 2005, 248, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Masson, F.; Calderon Copete, S.; Schupfer, F.; Garcia-Arraez, G.; Lemaitre, B. In Vitro Culture of the Insect Endosymbiont Spiroplasma poulsonii Highlights Bacterial Genes Involved in Host-Symbiont Interaction. MBio 2018, 9, e00024-18. [Google Scholar] [CrossRef]
- O’Neill, S.L.; Pettigrew, M.M.; Sinkins, S.P.; Braig, H.R.; Andreadis, T.G.; Tesh, R.B. In vitro cultivation of Wolbachia pipientis in an Aedes albopictus cell line. Insect Mol. Biol. 1997, 6, 33–39. [Google Scholar] [CrossRef]
- McGraw, E.A.; Merritt, D.J.; Droller, J.N.; O’Neill, S.L. Wolbachia-mediated sperm modification is dependent on the host genotype in Drosophila. Proc. Biol. Sci. 2001, 268, 2565–2570. [Google Scholar] [CrossRef]
- Minard, G.; Tran, F.H.; Raharimalala, F.N.; Hellard, E.; Ravelonandro, P.; Mavingui, P.; Valiente Moro, C. Prevalence, genomic and metabolic profiles of Acinetobacter and Asaia associated with field-caught Aedes albopictus from Madagascar. Fems. Microbiol. Ecol. 2013, 83, 63–73. [Google Scholar] [CrossRef]
- Mühling, M.; Woolven-Allen, J.; Murrell, J.C.; Joint, I. Improved group-specific PCR primers for denaturing gradient gel electrophoresis analysis of the genetic diversity of complex microbial communities. Isme. J. 2008, 2, 379. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Voronin, D.; Tran-Van, V.; Potier, P.; Mavingui, P. Transinfection and growth discrepancy of Drosophila Wolbachia strain wMel in cell lines of the mosquito Aedes albopictus. J. Appl. Microbiol. 2010, 108, 2133–2141. [Google Scholar] [PubMed]
- Mackie, A.M.; Hassan, K.A.; Paulsen, I.T.; Tetu, S.G. Biolog Phenotype Microarrays for phenotypic characterization of microbial cells. Methods Mol. Biol. 2014, 1096, 123–130. [Google Scholar] [PubMed]
- Mee, P.T.; Weeks, A.R.; Walker, P.J.; Hoffmann, A.A.; Duchemin, J.-B. Detection of Low-Level Cardinium and Wolbachia Infections in Culicoides. Appl. Env. Microbiol. 2015, 81, 6177–6188. [Google Scholar] [CrossRef] [PubMed]
- Turelli, M.; Hoffman, A.A. Microbe-induced cytoplasmic incompatibility as a mechanism for introducing transgenes into arthropod populations. Insect Mol. Biol. 1999, 8, 243–255. [Google Scholar] [CrossRef]
- Coutinho-Abreu, I.V.; Zhu, K.Y.; Ramalho-Ortigao, M. Transgenesis and paratransgenesis to control insect-borne diseases: Current status and future challenges. Parasitol. Int. 2010, 59, 1–8. [Google Scholar] [CrossRef]
- Kriesner, P.; Hoffmann, A.A.; Lee, S.F.; Turelli, M.; Weeks, A.R. Rapid sequential spread of two Wolbachia variants in Drosophila simulans. PLoS Pathog. 2013, 9, e1003607. [Google Scholar] [CrossRef]
- Berg, I.A.; Kockelkorn, D.; Buckel, W.; Fuchs, G. A 3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway in Archaea. Science 2007, 318, 1782–1786. [Google Scholar] [CrossRef]
- Cho, E.A.; Lee, D.W.; Cha, Y.H.; Lee, S.J.; Jung, H.C.; Pan, J.G.; Pyun, Y.R. Characterization of a novel D-lyxose isomerase from Cohnella laevoribosii RI-39 sp. nov. J. Bacteriol. 2007, 189, 1655–1663. [Google Scholar] [CrossRef][Green Version]
- Darby, A.C.; Armstrong, S.D.; Bah, G.S.; Kaur, G.; Hughes, M.A.; Kay, S.M.; Koldkjaer, P.; Rainbow, L.; Radford, A.D.; Blaxter, M.L.; et al. Analysis of gene expression from the Wolbachia genome of a filarial nematode supports both metabolic and defensive roles within the symbiosis. Genome Res. 2012, 22, 2467–2477. [Google Scholar] [CrossRef]
- Grote, A.; Voronin, D.; Ding, T.; Twaddle, A.; Unnasch, T.R.; Lustigman, S.; Ghedin, E. Defining Brugia malayi and Wolbachia symbiosis by stage-specific dual RNA-seq. PLoS Negl. Trop. Dis. 2017, 11, e0005357. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.A.; Ye, Y.H.; Turner, K.; Eyles, D.W.; McGraw, E.A.; O’Neill, S.L. The wMelPop strain of Wolbachia interferes with dopamine levels in Aedes aegypti. Parasit. Vectors 2011, 4, 28. [Google Scholar] [CrossRef] [PubMed]
- Rohrscheib, C.E.; Bondy, E.; Josh, P.; Riegler, M.; Eyles, D.; van Swinderen, B.; Weible, M.W.; Brownlie, J.C. Wolbachia Influences the Production of Octopamine and Affects Drosophila Male Aggression. Appl. Env. Microbiol. 2015, 81, 4573–4580. [Google Scholar]
- Valentine, R.C.; Bojanowski, R.; Gaudy, E.; Wolfe, R.S. Mechanism of the allantoin fermentation. J. Biol. Chem. 1962, 237, 2271–2277. [Google Scholar]
- Beckmann, J.F.; Ronau, J.A.; Hochstrasser, M. A Wolbachia deubiquitylating enzyme induces cytoplasmic incompatibility. Nat. Microbiol. 2017, 2, 17007. [Google Scholar] [CrossRef]
- Zhang, G.; Hussain, M.; Asgari, S. Regulation of arginine methyltransferase 3 by a Wolbachia-induced microRNA in Aedes aegypti and its effect on Wolbachia and dengue virus replication. Insect Biochem. Mol. Biol. 2014, 53, 81–88. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krafsur, A.M.; Ghosh, A.; Brelsfoard, C.L. Phenotypic Response of Wolbachia pipientis in a Cell-Free Medium. Microorganisms 2020, 8, 1060. https://doi.org/10.3390/microorganisms8071060
Krafsur AM, Ghosh A, Brelsfoard CL. Phenotypic Response of Wolbachia pipientis in a Cell-Free Medium. Microorganisms. 2020; 8(7):1060. https://doi.org/10.3390/microorganisms8071060
Chicago/Turabian StyleKrafsur, Alyssa M., Arnab Ghosh, and Corey L. Brelsfoard. 2020. "Phenotypic Response of Wolbachia pipientis in a Cell-Free Medium" Microorganisms 8, no. 7: 1060. https://doi.org/10.3390/microorganisms8071060
APA StyleKrafsur, A. M., Ghosh, A., & Brelsfoard, C. L. (2020). Phenotypic Response of Wolbachia pipientis in a Cell-Free Medium. Microorganisms, 8(7), 1060. https://doi.org/10.3390/microorganisms8071060