Population Genetics of Phlebotomus papatasi from Endemic and Nonendemic Areas for Zoonotic Cutaneous Leishmaniasis in Morocco, as Revealed by Cytochrome Oxidase Gene Subunit I Sequencing

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Origins and Sampling of Sand Flies
2.2. Morphological Identification
2.3. DNA Extraction, Cytochrome Oxidase Subunit I (COI) Amplification, and DNA Sequencing
2.4. Data Analysis
3. Results
3.1. Sequence Analysis
3.2. Haplotype Diversity and Distribution
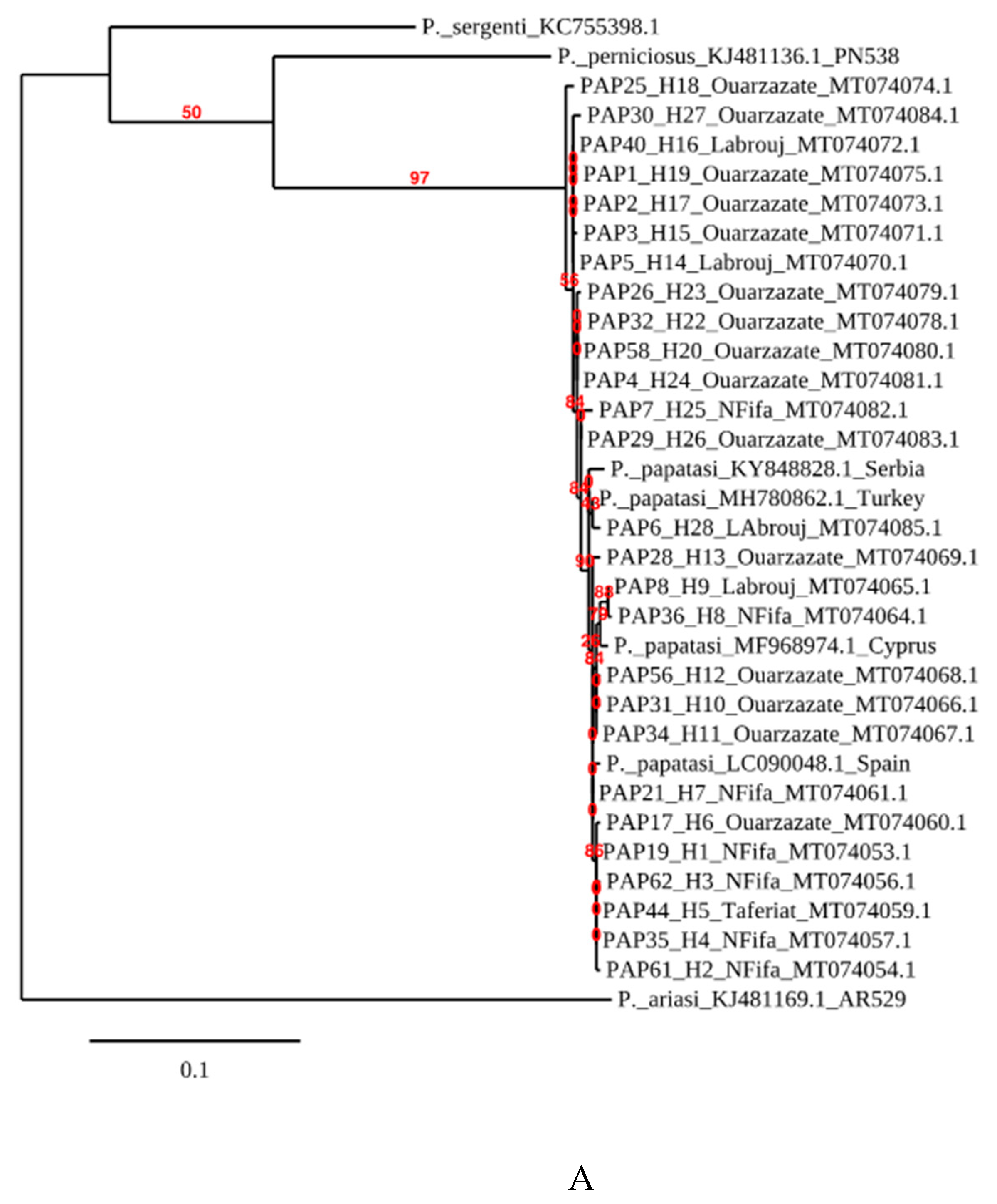
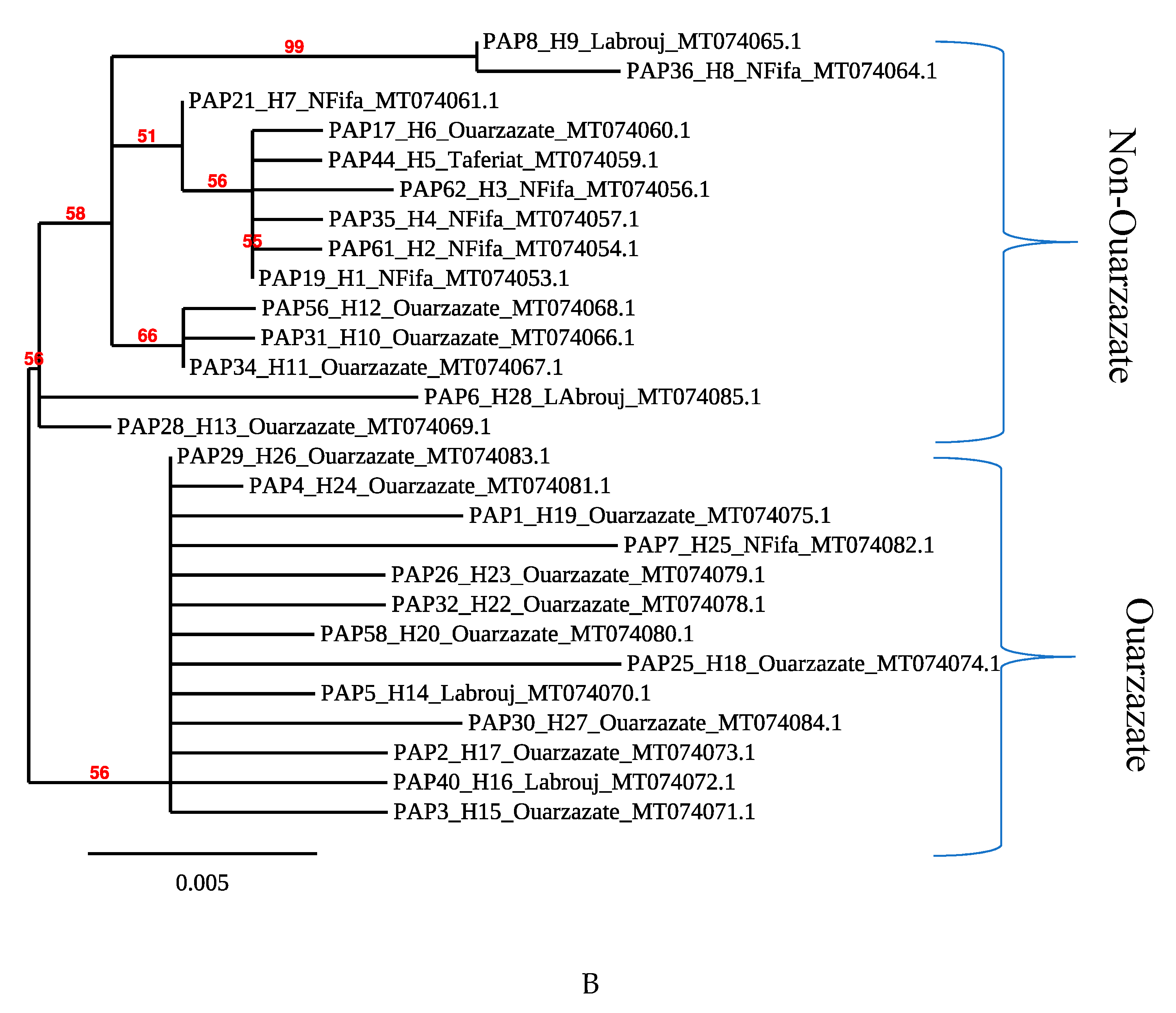
3.3. Phylogenetic Analysis
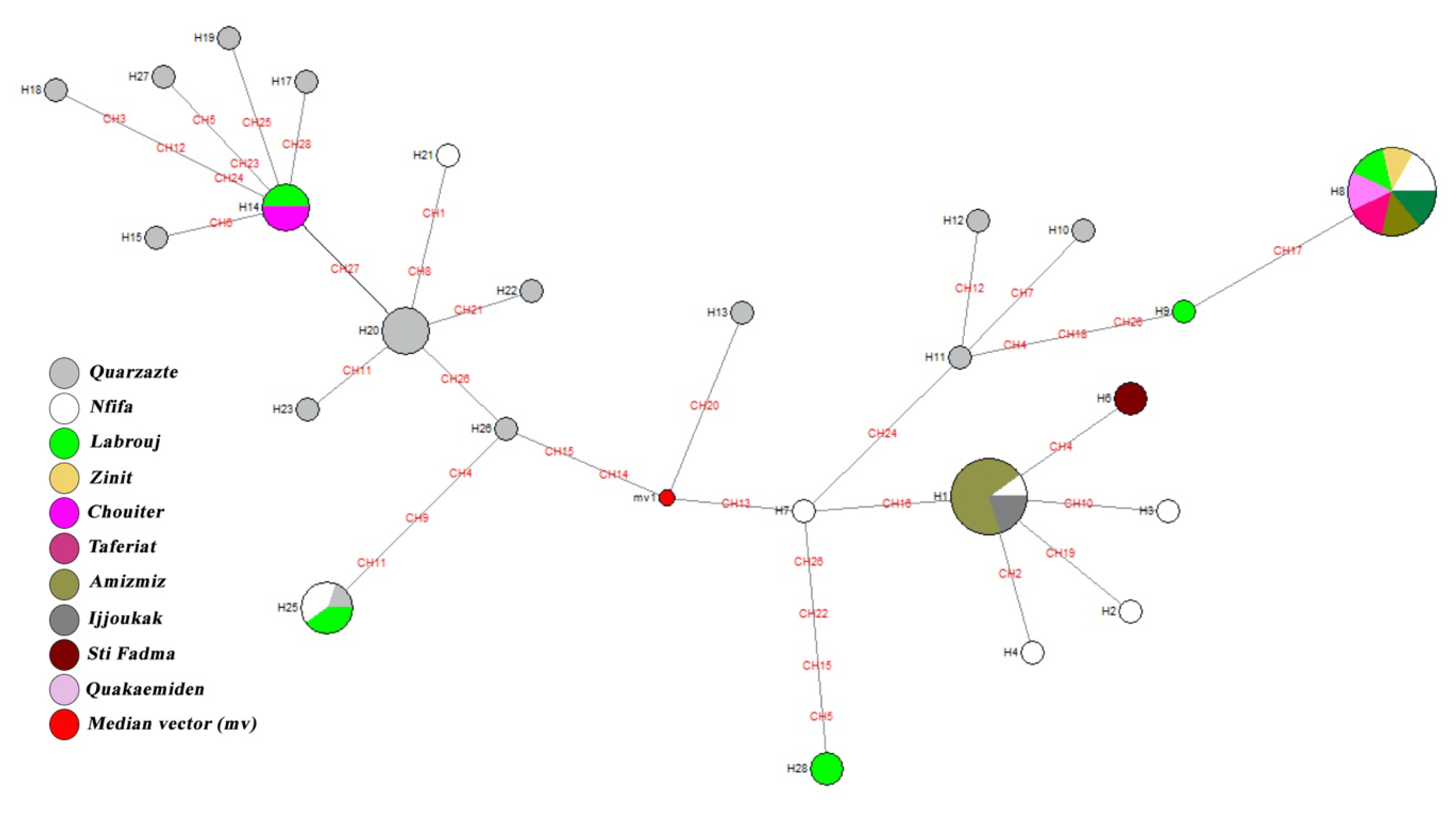
3.4. Median-Joining Network Analysis
3.5. Population Differentiation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sereno, D. Leishmania (Mundinia) spp.: From description to emergence as new human and animal Leishmania pathogens. New Microbes New Infect. 2019, 11540. [Google Scholar] [CrossRef]
- Sereno, D.; Akhoundi, M.; Sayehmri, K.; Mirzaei, A.; Holzmuller, P.; Lejon, V.; Waleckx, E. Noninvasive Biological Samples to Detect and Diagnose Infections due to Trypanosomatidae Parasites: A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2020, 21, 1684. [Google Scholar] [CrossRef] [PubMed]
- Akhoundi, M.; Kuhls, K.; Cannet, A.; Votýpka, J.; Marty, P.; Delaunay, P.; Sereno, D. A Historical Overview of the Classification, Evolution, and Dispersion of Leishmania Parasites and Sandflies. PLoS Negl. Trop. Dis. 2016, 10, e0004349. [Google Scholar] [CrossRef]
- Killick-Kendrick, R. Phlebotomine vectors of leishmaniasis: A review. Med. Vet. Entomol. 1990, 4, 1–24. [Google Scholar] [CrossRef]
- Killick-Kendrick, R. The biology and control of phlebotomine sand flies. Clin. Dermatol. 1999, 17, 279–289. [Google Scholar] [CrossRef]
- Maazoun, R.; Pratlong, F.; Lanotte, G.; Rioux, J.A. Le complexe Leishmania major. A propos de l’analyse numérique de 35 souches identifiées par la méthode enzymatique. In Leishmania: Taxonomie et Phylogenèse, Applications Écoépidémiologiques; Rioux, J.A., Ed.; CNRS/INSERM: Paris, France, 1984; IMEEE: Montpellier, France, 1986; pp. 139–142. [Google Scholar]
- Rioux, J.A.; Guilvard, E.; Dereure, J.; Lanotte, G.; Deniau, M.; Pratlong, F. Infestation naturelle de Phlebotomus papatasi (Scopoli, 1786) par Leishmania major MON-25. A propos de 28 souches isolées dans un foyer du Sud Marocain. In Leishmania: Taxonomie et Phylogenèse, Applications Eco-Epidémiologiques; Rioux, J.A., Ed.; CNRS/INSERM: Paris, France, 1984; IMEEE: Montpellier, France, 1986; pp. 471–480. [Google Scholar]
- Rioux, J.A.; Petter, F.; Akalay, O.; Lanotte, G.; Ouazani, A.; Séguignes, M.; Mohcine, A. Meriones Shawi (Duvernoy, 1842) [Rodentia, Gerbillidae], réservoir de Leishmania major Yakimoff et Shokhor, 1914 dans le Sud Marocain. Comptes Rendus Acad. Sci. Paris Sér. III 1982, 294, 515–517. [Google Scholar]
- MHM. Santé en Chiffres 2015; Direction de la Planification et des Ressources Financière, Division de la Planification et des Etudes, Service des Études et de L’information Sanitaire: Rabat, Morocco, 2016. [Google Scholar]
- Guernaoui, S. Les Leishmanioses dans les Zones Arides et Semi-Arides du Sud-Ouest Marocain. Ecologie, Épidémiologie, Modélisation et Aide à la Décision Pour la Lutte Anti-Vectorielle; Université Cadi Ayyad: Marrakech, Morocco, 2006. [Google Scholar]
- Kholoud, K.; Denis, S.; Lahouari, B.; El Hidan, M.; Souad, B. Management of Leishmaniases in the Era of Climate Change in Morocco. Int. J. Environ. Res. Public Health 2018, 15, 1542. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.J. A taxonomic review of the genus Phlebotomus (Diptera: Psychodidae). Bull. Br. Mus. Nat. Hist. 1982, 45, 121–209. [Google Scholar]
- Wasserberg, G.; Abramsky, Z.; Kotler, B.P.; Ostfeld, R.S.; Yarom, I.; Warburg, A. Anthropogenic disturbances enhance occurrence of cutaneous leishmaniasis in Israel deserts: Patterns and mechanisms. Ecol. Appl. 2003, 13, 868–881. [Google Scholar] [CrossRef]
- Kamhawi, S.; Abdel-Hafez, S.K.; Molyneux, D.H. Urbanization—How does it affect the behaviour of sandflies? Parassitologia 1991, 33, 299–306. [Google Scholar]
- Guernaoui, S.; Ramaoui, K.; Rahola, N.; Barnabe, C.; Sereno, D.; Boumezzough, A. Malformations of the genitalia in male Phlebotomus papatasi (Scopoli) (Diptera: Psychodidae). J. Vector Ecol. 2010, 35, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Guernaoui, S.; Boumezzough, A. Habitat preferences of sand flies (Diptera: Psychodidae) in the southwestern Morocco. J. Med. Entomol. 2009, 46, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Lysenko, A.J.; Bejaev, A.E. Some problems of primary importance concerning the epidemiology of leishmaniasis in the Mediterranean-Midlle Asian Region. In Proceedings of the Ecologie des Leishmanioses, Paris, France.
- Esseghir, S.; Ready, P.D.; Killick-Kendrick, R.; Ben-Ismail, R. Mitochondrial haplotypes and phylogeography of Phlebotomus vectors of Leishmania major. Insect. Mol. Biol. 1997, 6, 211–225. [Google Scholar] [CrossRef] [PubMed]
- Flanley, C.M.; Ramalho-Ortigao, M.; Coutinho-Abreu, I.V.; Mukbel, R.; Hanafi, H.A.; El-Hossary, S.S.; Fawaz, E.E.-D.Y.; Hoel, D.F.; Bray, A.W.; Stayback, G.; et al. Population genetics analysis of Phlebotomus papatasi sand flies from Egypt and Jordan based on mitochondrial cytochrome b haplotypes. Parasites Vectors 2018, 11, 214. [Google Scholar] [CrossRef] [PubMed]
- Parvizi, P.; Benlarbi, M.; Ready, P.D. Mitochondrial and Wolbachia markers for the sandfly Phlebotomus papatasi: Little population differentiation between peridomestic sites and gerbil burrows in Isfahan province, Iran. Med. Vet. Entomol. 2003, 17, 351–362. [Google Scholar] [CrossRef]
- Parvizi, P.; Ready, P.D. Molecular investigation of the population differentiation of Phlebotomus papatasi, important vector of Leishmania major, in different habitats and regions of Iran. Iran Biomed. J. 2006, 10, 69–77. [Google Scholar]
- Hamarsheh, O.; Presber, W.; Abdeen, Z.; Sawalha, S.; Al-Lahem, A.; Schönian, G. Genetic structure of Mediterranean populations of the sand fly Phlebotomus papatasi by mitochondrial cytochrome b haplotype analysis. Med. Vet. Entomol. 2007, 21, 270–277. [Google Scholar] [CrossRef]
- Raja, B.; Jaouadi, K.; Haouas, N.; Mezhoud, H.; Bdira, S.; Amor, S. Mitochondrial cytochrome b variation in populations of the cutaneous leishmaniasis vector Phlebotomus papatasi across eastern Tunisia. Int. J. Biodivers. Conserv. 2012, 4, 189–196. [Google Scholar]
- Depaquit, J.; Lienard, E.; Verzeaux-Griffon, A.; Ferte, H.; Bounamous, A.; Gantier, J.C.; Hanafi, H.A.; Jacobson, R.L.; Maroli, M.; Moin-Vaziri, V.; et al. Molecular homogeneity in diverse geographical populations of Phlebotomus papatasi (Diptera, Psychodidae) inferred from ND4 mtDNA and ITS2 rDNA Epidemiological consequences. Infect. Genet. Evol. 2008, 8, 159–170. [Google Scholar] [CrossRef]
- Hamarsheh, O.; Presber, W.; Yaghoobi-Ershadi, M.R.; Amro, A.; Al-Jawabreh, A.; Sawalha, S.; Al-Lahem, A.; Das, M.L.; Guernaoui, S.; Seridi, N.; et al. Population structure and geographical subdivision of the Leishmania major vector Phlebotomus papatasi as revealed by microsatellite variation. Med. Vet. Entomol. 2009, 23, 69–77. [Google Scholar] [CrossRef]
- Hamarsheh, O.; Karakus, M.; Azmi, K.; Jaouadi, K.; Yaghoobi-Ershadi, M.R.; Kruger, A.; Amro, A.; Kenawy, M.A.; Dokhan, M.R.; Abdeen, Z.; et al. Development of polymorphic EST microsatellite markers for the sand fly, Phlebotomus papatasi (Diptera: Psychodidae). Parasites Vectors 2018, 11, 160. [Google Scholar] [CrossRef] [PubMed]
- Khalid, N.M.; Aboud, M.A.; Alrabba, F.M.; Elnaiem, D.E.; Tripet, F. Evidence for genetic differentiation at the microgeographic scale in Phlebotomus papatasi populations from Sudan. Parasites Vectors 2012, 5, 249. [Google Scholar] [CrossRef] [PubMed]
- Bailly-Choumara, H.; Abonnenc, E.; Pastre, J. Contribution à l’étude des phlébotomes du Maroc (Diptera: Psychodidae). Données faunistiques et écologiques. Cahiers ORSTOM 1971, 9, 431–460. [Google Scholar]
- Rioux, J.A.; Rispail, P.; Lanotte, G.; Lepart, J. Relations Phlébotomes-bioclimats en écologie des leishmanioses Corollaires épidémiologiques. L’exemple du Maroc. Bull. Soc. Bot. Fr. 1984, 2–4, 549–557. [Google Scholar] [CrossRef]
- Rioux, J.A.; Akalay, O.; Perieres, J.; Dereure, J.; Mahjour, J.; Le Houerou, H.N.; Leger, N.; Desjeux, P.; Gallego, M.; Saddiki, A.; et al. L’évaluation écoépidémiologique du « risque leishmanien » au Sahara atlantique marocain. Intérêt heuristique de la relation « Phlébotomes-bioclimats ». Ecol. Mediterr. 1997, 23, 73–92. [Google Scholar] [CrossRef]
- Guernaoui, S.; Boumezzough, A.; Laamrani, A. Altitudinal structuring of sand flies (Diptera: Psychodidae) in the High-Atlas mountains (Morocco) and its relation to the risk of leishmaniasis transmission. Acta Trop. 2006, 97, 346–351. [Google Scholar] [CrossRef]
- Abonnenc, E. Les phlébotomes de la région éthiopienne (Diptera: Psychodidae). Mémoire ORSTOM 1972, 55, 289. [Google Scholar]
- Ratnasingham, S.; Hebert, P.D. Bold: The Barcode of Life Data System (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1998, 16, 10881–10890. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Tamura, K.; Nei, M. MEGA3: Integrated software for Molecular Evolutionary Genetics Analysis and sequence alignment. Brief Bioinform. 2004, 5, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Watterson, G.A. On the number of segregating sites in genetical models without recombination. Theor. Popul. Biol. 1975, 7, 256–276. [Google Scholar] [CrossRef]
- Tajima, F. Evolutionary relationship of DNA sequences in finite populations. Genetics 1983, 105, 437–460. [Google Scholar] [PubMed]
- Weir, B.S.; Cockerham, C.C. Group inbreeding with two linked loci. Genetics 1969, 63, 711–742. [Google Scholar] [PubMed]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software for population genetics data analysis. Evol. Bioinform. Online 2005, 1, 47–50. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Rohl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. Intraspecific gene genealogies: Trees grafting into networks. Trends Ecol. Evol. 2001, 16, 37–45. [Google Scholar] [CrossRef]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Erisoz Kasap, O.; Linton, Y.M.; Karakus, M.; Ozbel, Y.; Alten, B. Revision of the species composition and distribution of Turkish sand flies using DNA barcodes. Parasites Vectors 2019, 12, 410. [Google Scholar] [CrossRef]
- Kress, W.J.; Garcia-Robledo, C.; Uriarte, M.; Erickson, D.L. DNA barcodes for ecology, evolution, and conservation. Trends Ecol. Evol. 2015, 30, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Arrivillaga, J.C.; Norris, D.E.; Feliciangeli, M.D.; Lanzaro, G.C. Phylogeography of the neotropical sand fly Lutzomyia longipalpis inferred from mitochondrial DNA sequences. Infect. Genet. Evol. 2002, 2, 83–95. [Google Scholar] [CrossRef]
- Scarpassa, V.M.; Alencar, R.B. Lutzomyia umbratilis, the main vector of Leishmania guyanensis, represents a novel species complex? PLoS ONE 2012, 7, e37341. [Google Scholar] [CrossRef]
- Lehrter, V.; Banuls, A.L.; Leger, N.; Rioux, J.A.; Depaquit, J. Phlebotomus (Paraphlebotomus) chabaudi and Phlebotomus riouxi: Closely related species or synonyms? Parasite 2017, 24, 47. [Google Scholar] [CrossRef][Green Version]
- Sjostrand, A.E.; Sjodin, P.; Jakobsson, M. Private haplotypes can reveal local adaptation. BMC Genet. 2014, 15, 61. [Google Scholar] [CrossRef] [PubMed]
- Prudhomme, J.; Gunay, F.; Rahola, N.; Ouanaimi, F.; Guernaoui, S.; Boumezzough, A.; Bañuls, A.L.; Sereno, D.; Alten, B. Wing size and shape variation of Phlebotomus papatasi (Diptera: Psychodidae) populations from the south and north slopes of the Atlas Mountains in Morocco. J. Vector Ecol. 2012, 37. [Google Scholar] [CrossRef]
- Guernaoui, S.; Garcia, D.; Boumezzough, A.; Fontenille, D.; Sereno, D. Preliminary study on the galectin molecular diversity in Moroccoan Phlebotomus papatasi sandfly populations. J. Vector Borne Dis. 2011, 48, 67–71. [Google Scholar]
- Kamhawi, S.; Ramalho-Ortigao, M.; Pham, V.M.; Kumar, S.; Lawyer, P.G.; Turco, S.J.; Barillas-Mury, C.; Sacks, D.L.; Valenzuela, J.G. A role for insect galectins in parasite survival. Cell 2004, 119, 329–341. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stations | Latitude | Longitude | Altitude (m) | Number of Specimens | Trapping Method |
|---|---|---|---|---|---|
| Ouarzazate | 31°23′19 N | 5°59′15 W | 1648 | 20 | CDC trap |
| Nfifa | 31°32′40 N | 8°45′58 W | 322 | 10 | CDC trap |
| Labrouj | 32°30′0 N | 7°12′0 W | 388 | 6 | CDC trap |
| Chouiter | 31°34′0 N | 7°49′0 W | 529 | 2 | Sticky trap |
| Taferiat | 31°32′59 N | 7°36′29 W | 755 | 5 | Sticky trap |
| Amizmiz | 31°13′0 N | 8°15′0 W | 1004 | 8 | Sticky trap |
| Zinit | 31°4′60 N | 8°41′60 W | 1148 | 2 | CDC trap |
| Ijjoukak | 31°0′0 N | 8°10′0 W | 1233 | 2 | CDC trap |
| Sti Fadma | 31°13′0 N | 7°42′0 W | 1772 | 2 | CDC trap |
| Oukaemiden | 31°12′21 N | 7°51′51 W | 2613 | 2 | CDC trap |
| Haplotypes | Specimen Code | Nb | Variant Character Position |
|---|---|---|---|
| H1 | PAP78,70,43,41, 19,13,11,10,9 | 9 | GGTCGCA CCTTGGGCGA TTAGGAGTAG CTATAT |
| H2 | PAP61 | 1 | ....... .......... .C........ ...... |
| H3 | PAP62 | 1 | ....... ..A....... .......... ....G. |
| H4 | PAP35 | 1 | .A..... .......... .......... ...... |
| H5 | PAP44 | 1 | ....... .......... .......... ...C.. |
| H6 | PAP79,17 | 2 | ...T... .......... .......... ...... |
| H7 | PAP21 | 1 | ....... ........A. .......... ...... |
| H8 | PAP80,77,76,75,71,55,47, 46,45,36,22,18,12,9 | 14 | ...T... ........AG C.....A.G. ...... |
| H9 | PAP8 | 1 | ...T... ........A. C.....A.G. ...... |
| H10 | PAP31 | 1 | ......G ........A. ......A... ...... |
| H11 | PAP34 | 1 | ....... ........A. ......A... ...... |
| H12 | PAP56 | 1 | ....... ....A...A. ......A... ...... |
| H13 | PAP28 | 1 | ....... .....A..A. ..G....... ...... |
| H14 | PAP5,49 | 2 | ....... .....AATA. ........GA ...... |
| H15 | PAP3 | 1 | .....T. .....AATA. ........GA ...... |
| H16 | PAP72,40 | 2 | ....... .....AATA. ........GA .....C |
| H17 | PAP2 | 1 | ....... .....AATA. ........GA T..... |
| H18 | PAP25 | 1 | ..C.... ....AAATA. ......A.GA .C.... |
| H19 | PAP1 | 1 | ....... .....AATA. .......GGA ..G... |
| H20 | PAP58,33,24 | 3 | ....... .....AATA. ........G. ..G... |
| H21 | PAP37 | 1 | C...... T....AATA. ........G. ..G... |
| H22 | PAP32 | 1 | ....... .....AATA. ...A....G. ..G... |
| H23 | PAP26 | 1 | ....... ...C.AATA. ........G. ..G... |
| H24 | PAP4 | 1 | ....... .....AATA. ........G. ...... |
| H25 | PAP67,60,57,38,7 | 5 | ...T... .T.A.AATA. .......... ..G... |
| H26 | PAP29 | 1 | ....... .....AATA. .......... ...... |
| H27 | PAP30 | 1 | ....A.. .....AATA. .....G..GA ...... |
| H28 | PAP73,6 | 2 | ....A.. .......TA. ....A...T. ...... |
| Stations | COI-Haplotypes | Unique Haplotypes |
|---|---|---|
| Ouarzazate | H10, H11, H12, H13, H15, H17, H18, H19, H20, H22, H23, H24, H25, H26, H27 | 13 |
| Nfifa | H1, H2, H3, H4, H7, H8, H21, H25 | 5 |
| Labrouj | H8, H9, H14, H16, H25, H28 | 1 |
| Chouiter | H8, H14 | 0 |
| Taferiat | H5, H8 | 1 |
| Amizmiz | H1, H8 | 0 |
| Zinit | H8 | 0 |
| Ijjoukak | H1 | 0 |
| Sti Fadma | H6 | 0 |
| Oukaemiden | H8 | 0 |
| Source of Variation | * d.f | Sum of Squares | Variance Components | Variation (%) |
|---|---|---|---|---|
| Among populations | 1 | 31.716 | 1.08341 Va | 31.03 |
| Within populations | 57 | 137.284 | 2.40850 Vb | 68.97 |
| Total | 58 | 169.000 | 3.49190 | / |
| Fixation index (FST) | 0.31026 | / | / | / |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guernaoui, S.; Hamarsheh, O.; Garcia, D.; Fontenille, D.; Sereno, D. Population Genetics of Phlebotomus papatasi from Endemic and Nonendemic Areas for Zoonotic Cutaneous Leishmaniasis in Morocco, as Revealed by Cytochrome Oxidase Gene Subunit I Sequencing. Microorganisms 2020, 8, 1010. https://doi.org/10.3390/microorganisms8071010
Guernaoui S, Hamarsheh O, Garcia D, Fontenille D, Sereno D. Population Genetics of Phlebotomus papatasi from Endemic and Nonendemic Areas for Zoonotic Cutaneous Leishmaniasis in Morocco, as Revealed by Cytochrome Oxidase Gene Subunit I Sequencing. Microorganisms. 2020; 8(7):1010. https://doi.org/10.3390/microorganisms8071010
Chicago/Turabian StyleGuernaoui, Souad, Omar Hamarsheh, Deborah Garcia, Didier Fontenille, and Denis Sereno. 2020. "Population Genetics of Phlebotomus papatasi from Endemic and Nonendemic Areas for Zoonotic Cutaneous Leishmaniasis in Morocco, as Revealed by Cytochrome Oxidase Gene Subunit I Sequencing" Microorganisms 8, no. 7: 1010. https://doi.org/10.3390/microorganisms8071010
APA StyleGuernaoui, S., Hamarsheh, O., Garcia, D., Fontenille, D., & Sereno, D. (2020). Population Genetics of Phlebotomus papatasi from Endemic and Nonendemic Areas for Zoonotic Cutaneous Leishmaniasis in Morocco, as Revealed by Cytochrome Oxidase Gene Subunit I Sequencing. Microorganisms, 8(7), 1010. https://doi.org/10.3390/microorganisms8071010