Continuous Cell Lines from the European Biting Midge Culicoides nubeculosus (Meigen, 1830)

 ,
,  , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Generation of Primary C. nubeculosus Cultures
2.2. Establishment of C. nubeculosus Cell Lines
2.3. Characterisation of C. nubeculosus Cell Lines CNE/LULS44 and CNE/LULS47
2.4. Infection of C. nubeculosus Cell Line CNE/LULS44 with BTV
3. Results
3.1. Generation of C. nubeculosus Cell Lines
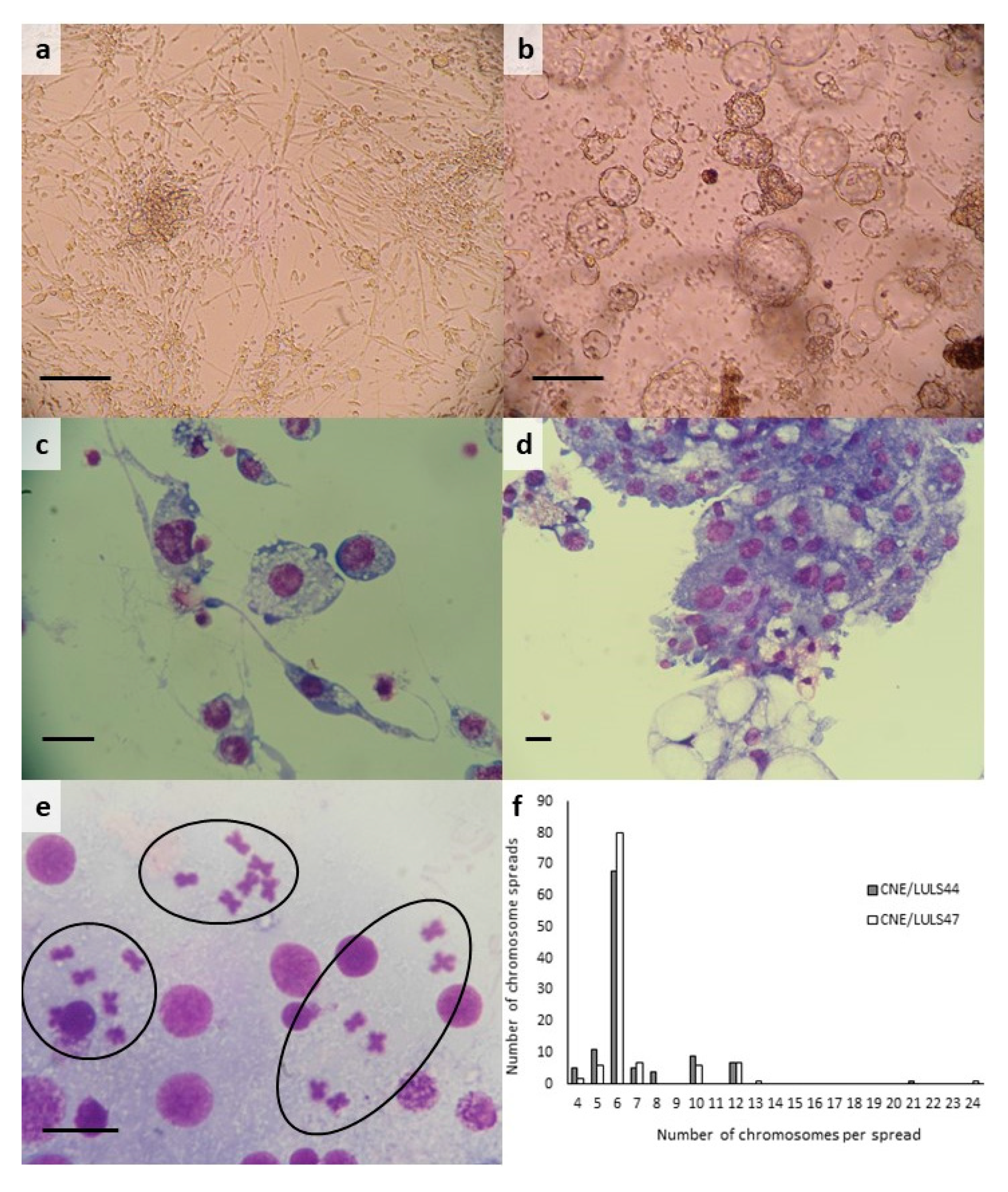
3.2. Morphology and Karyotype of C. nubeculosus Cell Lines
3.3. Replication of BTV Serotype 1 in CNE/LULS44 Cells
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carpenter, S.; Groschup, M.H.; Garros, C.; Felippe-Bauer, M.L.; Purse, B.V. Culicoides biting midges, arboviruses and public health in Europe. Antivir. Res. 2013, 100, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Purse, B.; Carpenter, S.; Venter, G.; Bellis, G.; Mullens, B. Bionomics of Temperate and Tropical Culicoides Midges: Knowledge Gaps and Consequences for Transmission of Culicoides-Borne Viruses. Annu. Rev. Entomol. 2015, 60, 373–392. [Google Scholar] [CrossRef] [PubMed]
- Linley, J.R. Biting Midges (Diptera: Ceratopogonidae) as Vectors of Nonviral Animal Pathogens. J. Med Entomol. 1985, 22, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, C.T. Epizootiology of Haemoproteus meleagridis (Protozoa: Haemosporina) in Florida: Potential Vectors and Prevalence in Naturally Infected Culicoides (Diptera: Ceratopogonidae). J. Med Entomol. 1988, 25, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Mellor, P.S. Studies on Onchocerca cervicalis Railliet and Henry 1910: I. Onchocerca cervicalis in British Horses. J. Helminthol. 1973, 47, 97–110. [Google Scholar] [CrossRef]
- Hoffmann, B.; Bauer, B.; Bauer, C.; Bätza, H.-J.; Beer, M.; Clausen, P.-H.; Geier, M.; Gethmann, J.; Kiel, E.; Liebisch, G.; et al. Monitoring of Putative Vectors of Bluetongue Virus Serotype 8, Germany. Emerg. Infect. Dis. 2009, 15, 1481–1484. [Google Scholar] [CrossRef]
- Jacquot, M.; Nomikou, K.; Palmarini, M.; Mertens, P.; Biek, R. Bluetongue virus spread in Europe is a consequence of climatic, landscape and vertebrate host factors as revealed by phylogeographic inference. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170919. [Google Scholar] [CrossRef]
- Elbers, A.R.W.; Meiswinkel, R.; Van Weezep, E.; Van Oldruitenborgh-Oosterbaan, M.M.S.; Kooi, E.A. Schmallenberg Virus in Culicoides spp. Biting Midges, the Netherlands, 2011. Emerg. Infect. Dis. 2013, 19, 106–109. [Google Scholar] [CrossRef]
- Balenghien, T.; Pages, N.; Goffredo, M.; Carpenter, S.; Augot, D.; Jacquier, E.; Talavera, S.; Monaco, F.; Depaquit, J.; Grillet, C.; et al. The emergence of Schmallenberg virus across Culicoides communities and ecosystems in Europe. Prev. Vet. Med. 2014, 116, 360–369. [Google Scholar] [CrossRef]
- Pages, N.; Talavera, S.; Verdún, M.; Pujol, N.; Valle, M.; Bensaid, A.; Pujols, J. Schmallenberg virus detection in Culicoides biting midges in Spain: First laboratory evidence for highly efficient infection of Culicoides of the Obsoletus complex and Culicoides imicola. Transbound. Emerg. Dis. 2017, 65, e1–e6. [Google Scholar] [CrossRef]
- Ségard, A.; Gardès, L.; Jacquier, E.; Grillet, C.; Mathieu, B.; Rakotoarivony, I.; Setier-Rio, M.-L.; Chavernac, D.; Cêtre-Sossah, C.; Balenghien, T.; et al. Schmallenberg virus in Culicoides Latreille (Diptera: Ceratopogonidae) populations in France during 2011–2012 outbreak. Transbound. Emerg. Dis. 2017, 65, e94–e103. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, S.; Mellor, P.S.; Fall, A.G.; Garros, C.; Venter, G.J. African Horse Sickness Virus: History, Transmission, and Current Status. Annu. Rev. Entomol. 2017, 62, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Mellor, P.; McCraig, J. The probable cause of “sweet itch” in England. Vet. Rec. 1974, 95, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Hendry, G.; Godwin, G. Biting midges in Scottish forestry: A costly irritant or a trivial nuisance? Scott. For. 1988, 42, 113–119. [Google Scholar]
- González, M.; Baldet, T.; Delécolle, J.C.; López, S.; Romón, P.; Goldarazena, A. Monitoring of Culicoides Latreille (Diptera: Ceratopogonidae) After Btv Outbreaks, in Sheep Farms and Natural Habitats from the Basque Country (Northern Spain). Proc. Entomol. Soc. Wash. 2013, 115, 48–69. [Google Scholar] [CrossRef]
- Mellor, P. A membrane feeding technique for the infection of Culicoides nubeculosus Mg. and Culicoides variipennis sonorensis Coq. with Onchocerca cervicalis Rail. and Henry. Trans. R. Soc. Trop. Med. Hyg. 1971, 65, 199–201. [Google Scholar] [CrossRef]
- Larska, M.; Grochowska, M.; Lechowski, L.; Żmudziński, J.F. Abundance and species composition of Culicoides spp. biting midges near cattle and horse in South-Eastern Poland. Acta Parasitol. 2017, 62, 739–747. [Google Scholar] [CrossRef]
- Dik, B.; Muz, D.; Muz, M.N.; Uslu, U. The geographical distribution and first molecular analysis of Culicoides Latreille (Diptera: Ceratopogonidae) species in the Southern and Southeastern Turkey during the 2012 outbreak of bovine ephemeral fever. Parasitol. Res. 2014, 113, 4225–4232. [Google Scholar] [CrossRef]
- Mellor, P.S.; Boorman, J. Simultaneous infection of Culicoides with bluetongue virus and filariae. Trans. R. Soc. Trop. Med. Hyg. 1980, 74, 117. [Google Scholar]
- Nayduch, D.; Cohnstaedt, L.W.; Saski, C.; Lawson, D.; Kersey, P.J.; Fife, M.; Carpenter, S. Studying Culicoides vectors of BTV in the post-genomic era: Resources, bottlenecks to progress and future directions. Virus Res. 2013, 182, 43–49. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Darby, A.C.; Baylis, M.; Makepeace, B.L. The Tick Cell Biobank: A global resource for in vitro research on ticks, other arthropods and the pathogens they transmit. Ticks Tick Borne Dis. 2018, 9, 1364–1371. [Google Scholar] [CrossRef] [PubMed]
- Wechsler, S.J.; McHolland, L.E.; Tabachnick, W.J. Cell lines from Culicoides variipennis (Diptera: Ceratopogonidae) support replication of bluetongue virus. J. Invertebr. Pathol. 1989, 54, 385–393. [Google Scholar] [CrossRef]
- Wechsler, S.; McHolland, L.; Wilson, W.C. A RNA virus in cells from Culicoides variipennis. J. Invertebr. Pathol. 1991, 57, 200–205. [Google Scholar] [CrossRef]
- McHolland, L.E.; Mecham, J.O. Characterization of cell lines developed from field populations of Culicoides sonorensis (Diptera: Ceratopogonidae). J. Med. Entomol. 2003, 40, 348–351. [Google Scholar] [CrossRef][Green Version]
- Mertens, P.P.; Burroughs, J.; Walton, A.; Wellby, M.; Fu, H.; O’Hara, R.; Brookes, S.M.; Mellor, P. Enhanced Infectivity of Modified Bluetongue Virus Particles for Two Insect Cell Lines and for Two Culicoides Vector Species. Virology 1996, 217, 582–593. [Google Scholar] [CrossRef][Green Version]
- Darpel, K.; Langner, K.F.A.; Nimtz, M.; Anthony, S.J.; Brownlie, J.; Takamatsu, H.-H.; Mellor, P.S.; Mertens, P.P. Saliva Proteins of Vector Culicoides Modify Structure and Infectivity of Bluetongue Virus Particles. PLoS ONE 2011, 6, e17545. [Google Scholar] [CrossRef]
- Eschbaumer, M.; Wernike, K.; Batten, C.; Savini, G.; Edwards, L.; Di Gennaro, A.; Teodori, L.; Oura, C.A.; Beer, M.; Hoffmann, B. Epizootic hemorrhagic disease virus serotype 7 in European cattle and sheep: Diagnostic considerations and effect of previous BTV exposure. Vet. Microbiol. 2012, 159, 298–306. [Google Scholar] [CrossRef]
- Pullinger, G.D.; Busquets, M.G.; Nomikou, K.; Boyce, M.; Attoui, H.; Mertens, P.P. Identification of the Genome Segments of Bluetongue Virus Serotype 26 (Isolate KUW2010/02) that Restrict Replication in a Culicoides sonorensis Cell Line (KC Cells). PLoS ONE 2016, 11, e0149709. [Google Scholar] [CrossRef]
- Schnettler, E.; Ratinier, M.; Watson, M.; Shaw, A.E.; McFarlane, M.; Varela, M.; Elliott, R.M.; Palmarini, M.; Kohl, A. RNA Interference Targets Arbovirus Replication in Culicoides cells. J. Virol. 2012, 87, 2441–2454. [Google Scholar] [CrossRef]
- Ghosh, A.; Jasperson, D.; Cohnstaedt, L.W.; Brelsfoard, C.L. Transfection of Culicoides sonorensis biting midge cell lines with Wolbachia pipientis. Parasites Vectors 2019, 12, 483. [Google Scholar] [CrossRef]
- Boorman, J. The maintenance of laboratory colonies of Culicoides variipennis (Coq.), C. nubeculosus (Mg.) and C. riethi Kieff. (Diptera, Ceratopogonidae). Bull. Entomol. Res. 1974, 64, 371–377. [Google Scholar] [CrossRef]
- Munderloh, U.G.; Kurtti, T.J. Formulation of medium for tick cell culture. Exp. Appl. Acarol. 1989, 7, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Maan, S.; Maan, N.S.; Nomikou, K.; Veronesi, E.; Bachanek-Bankowska, K.; Belaganahalli, M.; Attoui, H.; Mertens, P.P. Complete Genome Characterisation of a Novel 26th Bluetongue Virus Serotype from Kuwait. PLoS ONE 2011, 6, e26147. [Google Scholar] [CrossRef]
- Attoui, H.; Billoir, F.; Cantaloube, J.F.; Biagini, P.; De Micco, P.; De Lamballerie, X. Strategies for the sequence determination of viral dsRNA genomes. J. Virol. Methods 2000, 89, 147–158. [Google Scholar] [CrossRef]
- Hofmann, M.; Griot, C.; Chaignat, V.; Perler, L.; Thür, B. Blauzungunkrankheit erreicht die Schweiz. Schweiz. Arch. Tierheilkunde 2008, 150, 49–56. [Google Scholar] [CrossRef]
- Sato, M.; Maeda, H.; Yoshida, H.; Urade, M.; Saito, S.; Miyazaki, T.; Shibata, T.; Watanabe, M. Plaque formation of herpes virus hominis Type 2 and rubella virus in variants isolated from the colonies of BHK21/W1-2 cells formed in soft agar. Arch. Virol. 1977, 53, 269–273. [Google Scholar] [CrossRef]
- Jaafar, F.M.; Attoui, H.; Mertens, P.P.; De Micco, P.; De Lamballerie, X. Structural organization of an encephalitic human isolate of Banna virus (genus Seadornavirus, family Reoviridae). J. Gen. Virol. 2005, 86, 1147–1157. [Google Scholar] [CrossRef]
- Jaafar, F.M.; Belhouchet, M.; Vitour, D.; Adam, M.; Bréard, E.; Zientara, S.; Mertens, P.P.; Attoui, H. Immunisation with bacterial expressed VP2 and VP5 of bluetongue virus (BTV) protect α/β interferon-receptor knock-out (IFNAR−/−) mice from homologous lethal challenge. Vaccine 2014, 32, 4059–4067. [Google Scholar] [CrossRef]
- Nunamaker, R.A.; Brown, S.E.; Knudson, D. Metaphase Chromosomes of Culicoides variipennis (Diptera: Ceratopogonidae). J. Med Entomol. 1996, 33, 871–873. [Google Scholar] [CrossRef] [PubMed]
- Bell-Sakyi, L. Continuous Cell Lines from the Tick Hyalomma anatolicum anatolicum. J. Parasitol. 1991, 77, 1006. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, A.; (Luntz), A.J.M.; Mordue, W. Morphology of the antennae of two species of biting midge: Culicoides impunctatus (G oetghebuer) and Culicoides nubeculosus (Meigen) (Diptera, Ceratopogonidae). J. Morphol. 1992, 213, 85–103. [Google Scholar] [CrossRef] [PubMed]
- Filimonova, S. Morphological Study of Digestive Cycle in Bloodsucking Biting Midges of Genus Cu1icoides. J. Evol. Biochem. Physiol. 2005, 41, 221–232. [Google Scholar] [CrossRef]
- Bell-Sakyi, L.; Attoui, H. Article Commentary: Virus Discovery Using Tick Cell Lines. Evol. Bioinform. 2016, 12, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Bell-Sakyi, L.; Zweygarth, E.; Blouin, E.F.; Gould, E.A.; Jongejan, F. Tick cell lines: Tools for tick and tick-borne disease research. Trends Parasitol. 2007, 23, 450–457. [Google Scholar] [CrossRef]
- Rey, G.J.; Ferro, C.; Bello, F.J. Establishment and characterization of a new continuous cell line from Lutzomyia longipalpis (Diptera: Psychodidae) and its susceptibility to infections with arboviruses and Leishmania chagasi. Mem. Inst. Oswaldo Cruz 2000, 95, 103–110. [Google Scholar] [CrossRef]
- Mazzacano, C.A.; Munderloh, U.G.; Kurtti, T.J. Characterization of a new continuous cell line from the flood water mosquito, Aedes vexans. Cytotechnology 1991, 5, 147–154. [Google Scholar] [CrossRef]
- Batten, C.; Darpel, K.; Henstock, M.; Fay, P.; Veronesi, E.; Gubbins, S.; Graves, S.; Frost, L.; Oura, C. Evidence for Transmission of Bluetongue Virus Serotype 26 through Direct Contact. PLoS ONE 2014, 9, e96049. [Google Scholar] [CrossRef]
- Veronesi, E.; Antony, F.; Gubbins, S.; Golding, N.; Blackwell, A.; Mertens, P.P.; Brownlie, J.; Darpel, K.; Mellor, P.S.; Carpenter, S. Measurement of the Infection and Dissemination of Bluetongue Virus in Culicoides Biting Midges Using a Semi-Quantitative RT-PCR Assay and Isolation of Infectious Virus. PLoS ONE 2013, 8, e70800. [Google Scholar] [CrossRef]
- Jennings, D.; Mellor, P. The vector potential of British Culicoides species for bluetongue virus. Vet. Microbiol. 1988, 17, 1–10. [Google Scholar] [CrossRef]
- Fu, H.; Leake, C.J.; Mertens, P.P.C.; Mellor, P.S. The barriers to bluetongue virus infection, dissemination and transmission in the vector, Culicoides variipennis (Diptera: Ceratopogonidae). Arch. Virol. 1999, 144, 747–761. [Google Scholar] [CrossRef] [PubMed]
- Mellor, P.S.; Boorman, J.; Baylis, M. Culicoides Biting Midges: Their Role as Arbovirus Vectors. Annu. Rev. Entomol. 2000, 45, 307–340. [Google Scholar] [CrossRef]
- Mills, M.; Ruder, M.G.; Nayduch, D.; Michel, K.; Drolet, B.S. Dynamics of epizootic hemorrhagic disease virus infection within the vector, Culicoides sonorensis (Diptera: Ceratopogonidae). PLoS ONE 2017, 12, e0188865. [Google Scholar] [CrossRef]
- Boorman, J.; Gibbs, E.P.J. Multiplication of the virus of epizootic haemorrhagic disease of deer in Culicoides species (Diptera, Ceratopogonidae). Arch. Virol. 1973, 41, 259–266. [Google Scholar] [CrossRef]
- Mellor, P.S.; Boorman, J.; Jennings, M. The multiplication of African horse-sickness virus in two species of Culicoides (Diptera, Ceratopogonidae). Arch. Virol. 1975, 47, 351–356. [Google Scholar] [CrossRef]
- Wellby, M.P.; Baylis, M.; Rawlings, P.; Mellor, P.S. Effect of temperature on survival and rate of virogenesis of African horse sickness virus in Culicoides variipennis sonorensis (Diptera: Ceratopogonidae) and its significance in relation to the epidemiology of the disease. Bull. Entomol. Res. 1996, 86, 715–720. [Google Scholar] [CrossRef]
- Mellor, P.S.; Jennings, M. Replication of Eubenangee virus in Culicoides nubecuIosus (Mg.) and Culicoides variipennis (Coq.). Arch. Virol. 1980, 63, 203–208. [Google Scholar] [CrossRef]
- Jennings, M.; Mellor, P. Culicoides: Biological vectors of Akabane virus. Vet. Microbiol. 1989, 21, 125–131. [Google Scholar] [CrossRef]
- Mellor, P.S.; Boorman, J.; Loke, R. The multiplication of Main Drain virus in two species of CuIicoides (Diptera, Ceratopogonidae). Arch. Gesamte Virusforsch. 1974, 46, 105–110. [Google Scholar] [CrossRef]
- Möhlmann, T.W.R.; Oymans, J.; Schreur, P.J.W.; Koenraadt, C.J.M.; Kortekaas, J.; Vogels, C.B.F. Vector competence of biting midges and mosquitoes for Shuni virus. PLoS Negl. Trop. Dis. 2019, 13, e0006609. [Google Scholar] [CrossRef] [PubMed]
- Kurtti, T.J.; Munderloh, U.G.; Andreadis, T.G.; Magnarelli, L.A.; Mather, T.N. Tick Cell Culture Isolation of an Intracellular Prokaryote from the tick Ixodes scapularis. J. Invertebr. Pathol. 1996, 67, 318–321. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, S.; Kurtti, T.J.; Noda, H. In Vitro Cultivation and Antibiotic Susceptibility of a Cytophaga-Like Intracellular Symbiote Isolated from the tick Ixodes scapularis. Curr. Microbiol. 2006, 52, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Pilgrim, J.; Ander, M.; Garros, C.; Baylis, M.; Hurst, G.D.D.; Siozios, S. Torix group Rickettsia are widespread in Culicoides biting midges (Diptera: Ceratopogonidae), reach high frequency and carry unique genomic features. Environ. Microbiol. 2017, 19, 4238–4255. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, E.; Venter, G.; Labuschagne, K.; Mellor, P.; Carpenter, S. Life-history parameters of Culicoides (Avaritia) imicola Kieffer in the laboratory at different rearing temperatures. Vet. Parasitol. 2009, 163, 370–373. [Google Scholar] [CrossRef] [PubMed]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bell-Sakyi, L.; Mohd Jaafar, F.; Monsion, B.; Luu, L.; Denison, E.; Carpenter, S.; Attoui, H.; Mertens, P.P.C. Continuous Cell Lines from the European Biting Midge Culicoides nubeculosus (Meigen, 1830). Microorganisms 2020, 8, 825. https://doi.org/10.3390/microorganisms8060825
Bell-Sakyi L, Mohd Jaafar F, Monsion B, Luu L, Denison E, Carpenter S, Attoui H, Mertens PPC. Continuous Cell Lines from the European Biting Midge Culicoides nubeculosus (Meigen, 1830). Microorganisms. 2020; 8(6):825. https://doi.org/10.3390/microorganisms8060825
Chicago/Turabian StyleBell-Sakyi, Lesley, Fauziah Mohd Jaafar, Baptiste Monsion, Lisa Luu, Eric Denison, Simon Carpenter, Houssam Attoui, and Peter P. C. Mertens. 2020. "Continuous Cell Lines from the European Biting Midge Culicoides nubeculosus (Meigen, 1830)" Microorganisms 8, no. 6: 825. https://doi.org/10.3390/microorganisms8060825
APA StyleBell-Sakyi, L., Mohd Jaafar, F., Monsion, B., Luu, L., Denison, E., Carpenter, S., Attoui, H., & Mertens, P. P. C. (2020). Continuous Cell Lines from the European Biting Midge Culicoides nubeculosus (Meigen, 1830). Microorganisms, 8(6), 825. https://doi.org/10.3390/microorganisms8060825