Neospora caninum: Differential Proteome of Multinucleated Complexes Induced by the Bumped Kinase Inhibitor BKI-1294

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Culture Media, Biochemicals, and Drugs
2.2. Host Cell Cultivation and Parasite Maintenance
2.3. Microscopy
2.4. Proteomics
2.5. Measurement of RNA Expression Levels by Quantitative Reverse Transcriptase Real-Time PCR
2.6. Statistics
3. Results
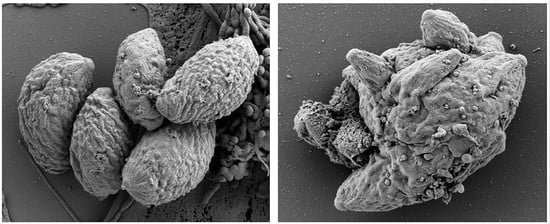
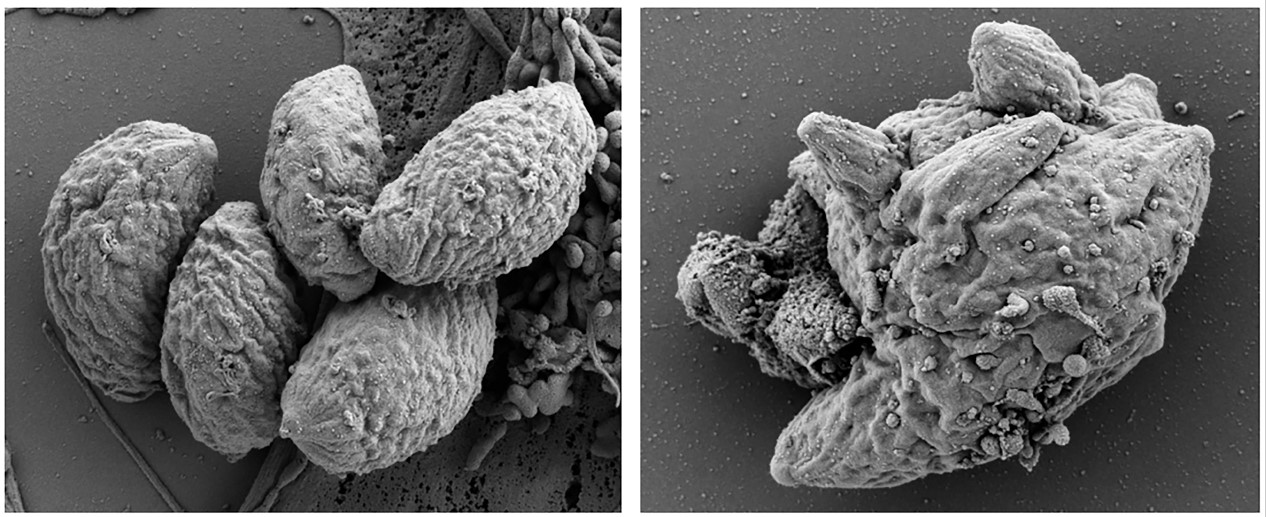
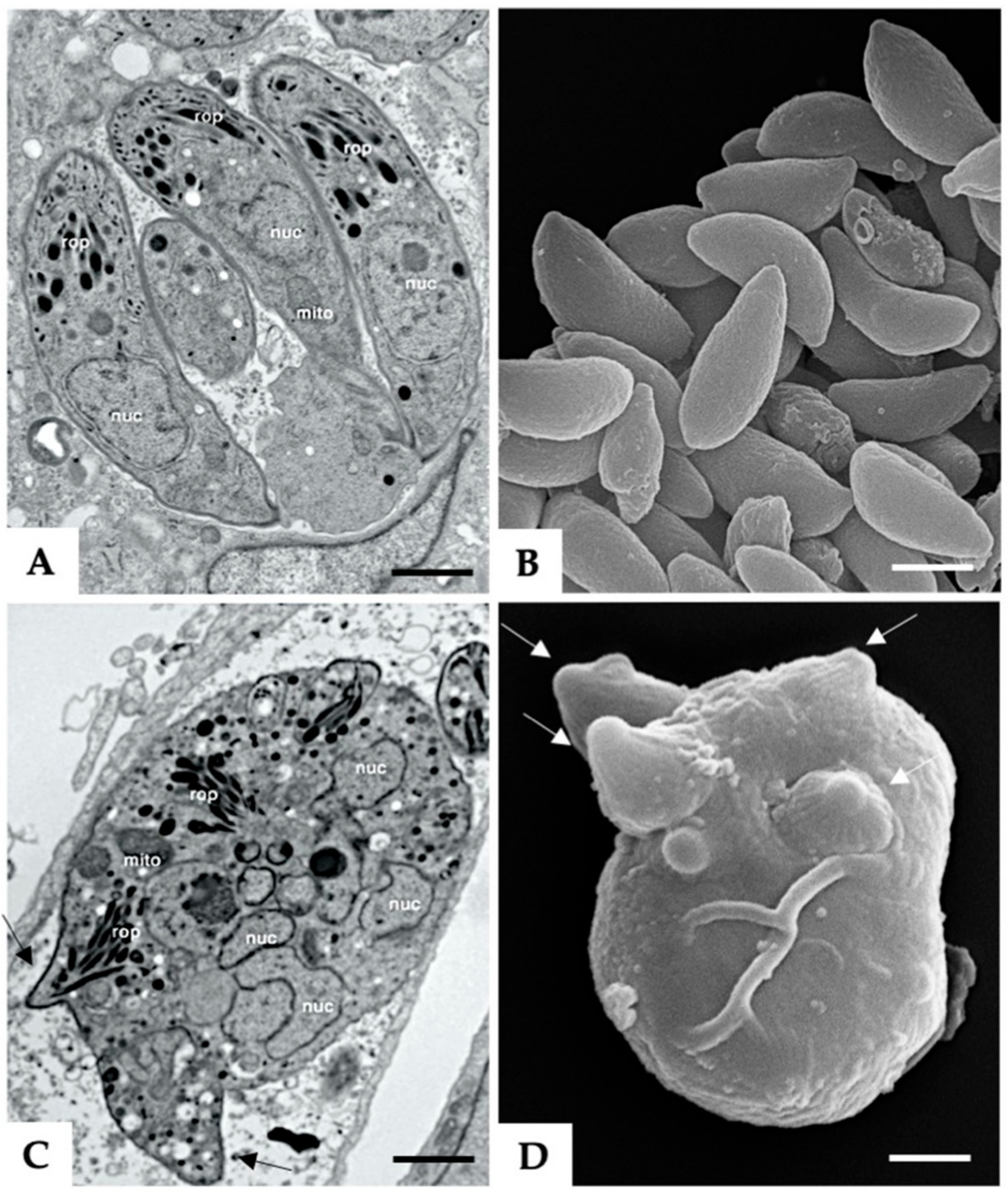
3.1. Formation of MNCs upon BKI-1294 Treatment In Vitro
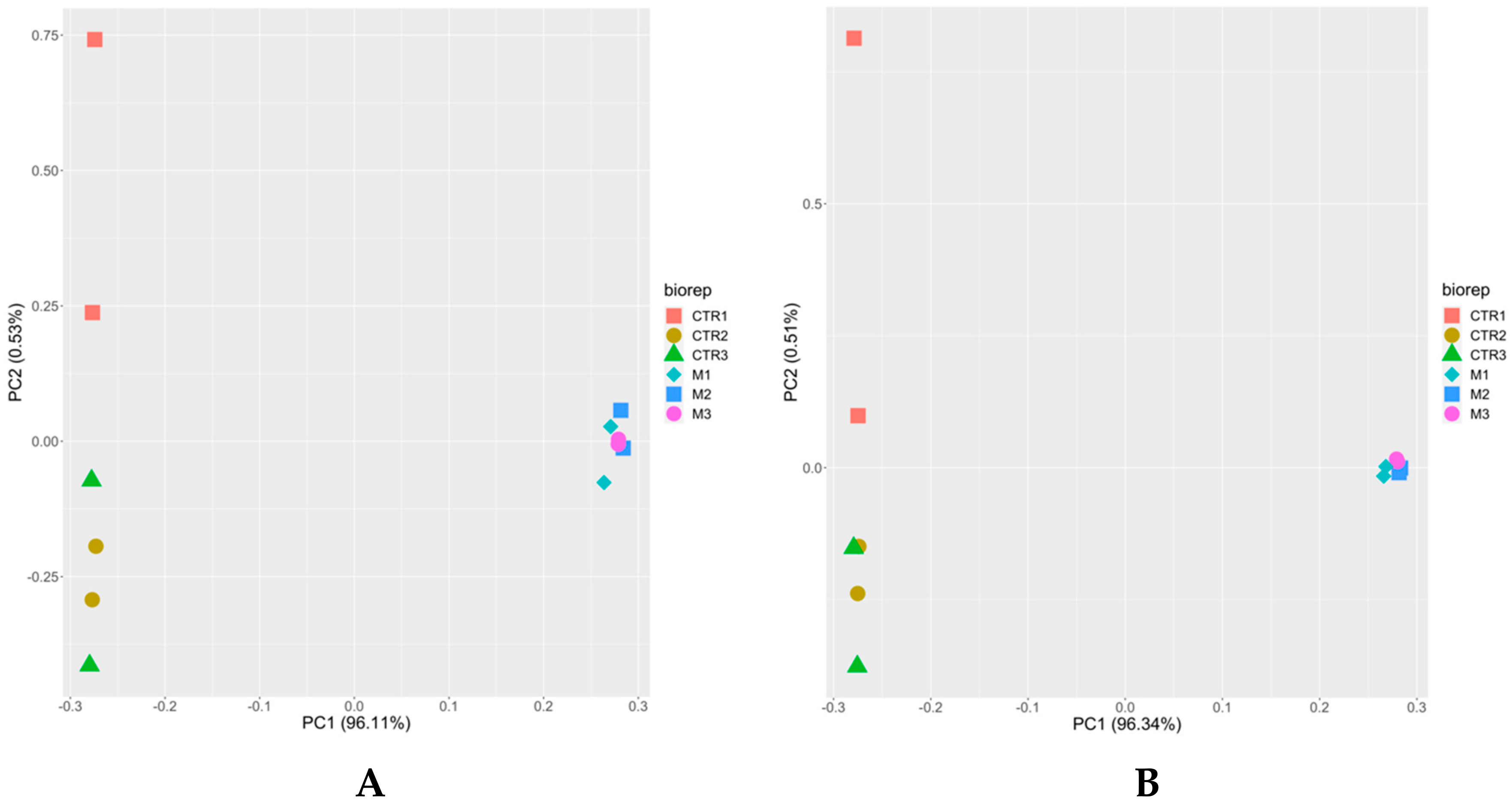
3.2. The Proteome of Multinucleated Complexes Is Different from the Proteome of Tachyzoites
3.3. Multinucleated Complexes Express a Distinct Set of Potential Surface Antigens
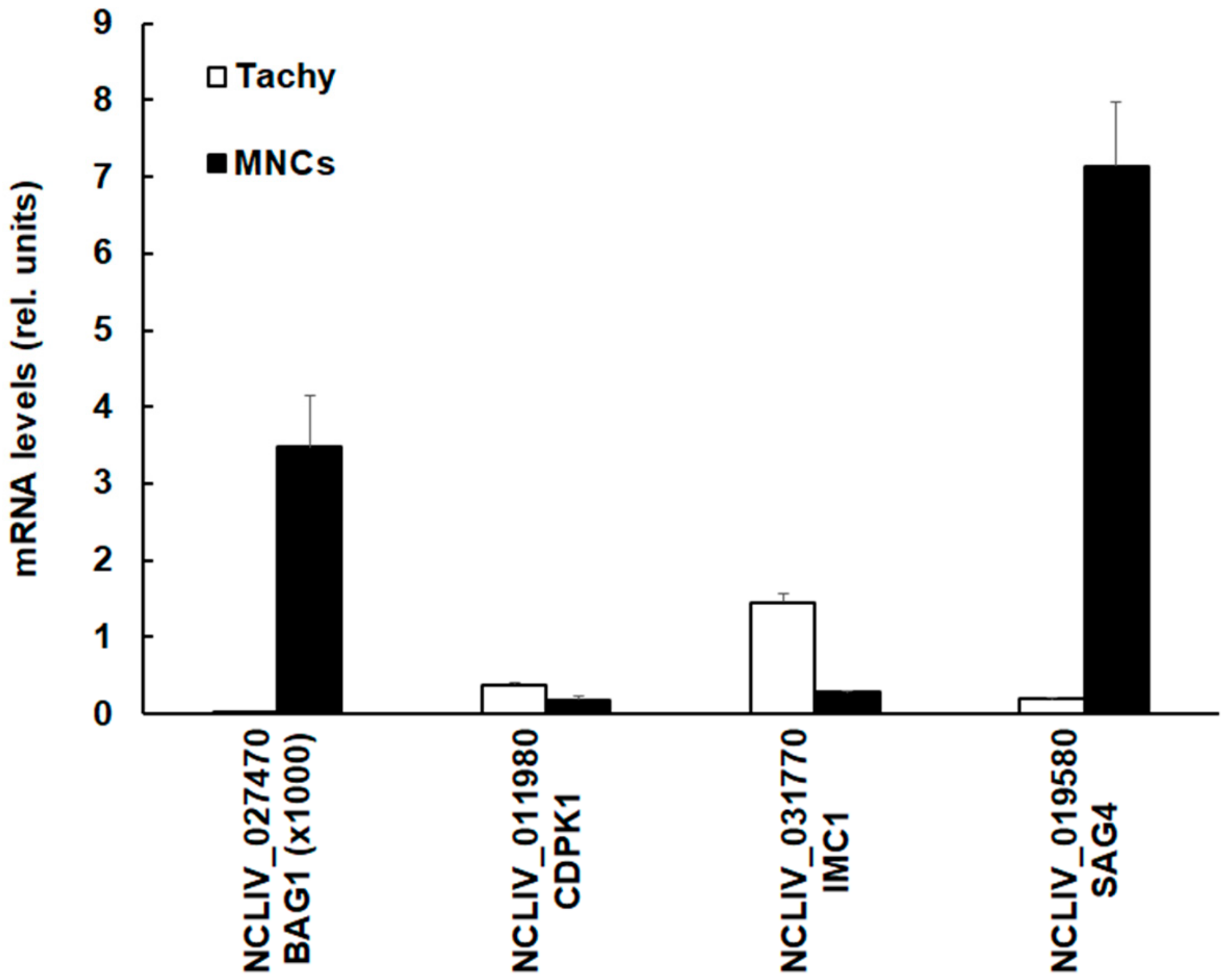
3.4. The SRS Domain Containing Protein NCLIV_019580 (NcSAG4) Has Higher mRNA Expression Levels in MNCs Compared to Tachyzoites
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goodswen, S.J.; Kennedy, P.J.; Ellis, J.T. A review of the infection, genetics, and evolution of Neospora caninum: From the past to the present. Infect. Genet. Evol. 2013, 13, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Coelho, A.M.; Cherubini, G.; De Stefani, A.; Negrin, A.; Gutierrez-Quintana, R.; Bersan, E.; Guevar, J. Serological prevalence of toxoplasmosis and neosporosis in dogs diagnosed with suspected meningoencephalitis in the UK. J. Small Anim. Pract. 2019, 60, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; Hemphill, A.; Calero-Bernal, R.; Schares, G. Neosporosis in Animals; Taylor & Francis: Boca Raton, FL, USA, 2017. [Google Scholar]
- Hemphill, A.; Aguado-Martinez, A.; Müller, J. Approaches for the vaccination and treatment of Neospora caninum infections in mice and ruminant models. Parasitology 2016, 143, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Reichel, M.P.; McAllister, M.M.; Pomroy, W.E.; Campero, C.; Ortega-Mora, L.M.; Ellis, J.T. Control options for Neospora caninum--is there anything new or are we going backwards? Parasitology 2014, 141, 1455–1470. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Hemphill, A. New approaches for the identification of drug targets in protozoan parasites. Int. Rev. Cell Mol. Biol. 2013, 301, 359–401. [Google Scholar] [CrossRef]
- Van Voorhis, W.C.; Doggett, J.S.; Parsons, M.; Hulverson, M.A.; Choi, R.; Arnold, S.L.M.; Riggs, M.W.; Hemphill, A.; Howe, D.K.; Mealey, R.H.; et al. Extended-spectrum antiprotozoal bumped kinase inhibitors: A review. Exp. Parasitol. 2017, 180, 71–83. [Google Scholar] [CrossRef]
- Choi, R.; Hulverson, M.A.; Huang, W.; Vidadala, R.S.R.; Whitman, G.R.; Barrett, L.K.; Schaefer, D.A.; Betzer, D.P.; Riggs, M.W.; Doggett, J.S.; et al. Bumped Kinase Inhibitors as therapy for apicomplexan parasitic diseases: Lessons learned. Int. J. Parasitol. 2020. [Google Scholar] [CrossRef]
- Ojo, K.K.; Larson, E.T.; Keyloun, K.R.; Castaneda, L.J.; Derocher, A.E.; Inampudi, K.K.; Kim, J.E.; Arakaki, T.L.; Murphy, R.C.; Zhang, L.; et al. Toxoplasma gondii calcium-dependent protein kinase 1 is a target for selective kinase inhibitors. Nat. Struct. Mol. Biol. 2010, 17, 602–607. [Google Scholar] [CrossRef]
- Johnson, S.M.; Murphy, R.C.; Geiger, J.A.; DeRocher, A.E.; Zhang, Z.; Ojo, K.K.; Larson, E.T.; Perera, B.G.; Dale, E.J.; He, P.; et al. Development of Toxoplasma gondii calcium-dependent protein kinase 1 (TgCDPK1) inhibitors with potent anti-toxoplasma activity. J. Med. Chem. 2012, 55, 2416–2426. [Google Scholar] [CrossRef]
- Lourido, S.; Tang, K.; Sibley, L.D. Distinct signalling pathways control Toxoplasma egress and host-cell invasion. EMBO J. 2012, 31, 4524–4534. [Google Scholar] [CrossRef]
- Winzer, P.; Müller, J.; Aguado-Martinez, A.; Rahman, M.; Balmer, V.; Manser, V.; Ortega-Mora, L.M.; Ojo, K.K.; Fan, E.; Maly, D.J.; et al. In vitro and In vivo Effects of the bumped kinase inhibitor 1294 in the related cyst-forming apicomplexans Toxoplasma gondii and Neospora caninum. Antimicrob. Agents Chemother. 2015, 59, 6361–6374. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Aguado-Martinez, A.; Ortega-Mora, L.M.; Moreno-Gonzalo, J.; Ferre, I.; Hulverson, M.A.; Choi, R.; McCloskey, M.C.; Barrett, L.K.; Maly, D.J.; et al. Development of a murine vertical transmission model for Toxoplasma gondii oocyst infection and studies on the efficacy of bumped kinase inhibitor (BKI)-1294 and the naphthoquinone buparvaquone against congenital toxoplasmosis. J. Antimicrob. Chemother. 2017, 72, 2334–2341. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Sanchez, R.; Ferre, I.; Re, M.; Ramos, J.J.; Regidor-Cerrillo, J.; Pizarro Diaz, M.; Gonzalez-Huecas, M.; Tabanera, E.; Benavides, J.; Hemphill, A.; et al. Treatment with bumped kinase inhibitor 1294 is safe and leads to significant protection against abortion and vertical transmission in sheep experimentally Iifected with Toxoplasma gondii during pregnancy. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Aguado-Martinez, A.; Balmer, V.; Maly, D.J.; Fan, E.; Ortega-Mora, L.M.; Ojo, K.K.; Van Voorhis, W.C.; Hemphill, A. Two novel calcium-dependent protein kinase 1 Inhibitors interfere with vertical transmission in mice infected with Neospora caninum tachyzoites. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Sanchez, R.; Ferre, I.; Re, M.; Vazquez, P.; Ferrer, L.M.; Blanco-Murcia, J.; Regidor-Cerrillo, J.; Pizarro Diaz, M.; Gonzalez-Huecas, M.; Tabanera, E.; et al. Safety and efficacy of the bumped kinase inhibitor BKI-1553 in pregnant sheep experimentally infected with Neospora caninum tachyzoites. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 112–124. [Google Scholar] [CrossRef]
- Ojo, K.K.; Reid, M.C.; Kallur Siddaramaiah, L.; Müller, J.; Winzer, P.; Zhang, Z.; Keyloun, K.R.; Vidadala, R.S.; Merritt, E.A.; Hol, W.G.; et al. Neospora caninum calcium-dependent protein kinase 1 is an effective drug target for neosporosis therapy. PLoS ONE 2014, 9, e92929. [Google Scholar] [CrossRef]
- Winzer, P.; Anghel, N.; Imhof, D.; Balmer, V.; Ortega-Mora, L.M.; Ojo, K.K.; Van Voorhis, W.C.; Müller, J.; Hemphill, A. Neospora caninum: Structure and fate of multinucleated complexes induced by the bumped kinase inhibitor BKI-1294. Pathogens 2020, 9, 382. [Google Scholar] [CrossRef]
- Marugan-Hernandez, V.; Alvarez-Garcia, G.; Risco-Castillo, V.; Regidor-Cerrillo, J.; Ortega-Mora, L.M. Identification of Neospora caninum proteins regulated during the differentiation process from tachyzoite to bradyzoite stage by DIGE. Proteomics 2010, 10, 1740–1750. [Google Scholar] [CrossRef]
- Horcajo, P.; Xia, D.; Randle, N.; Collantes-Fernandez, E.; Wastling, J.; Ortega-Mora, L.M.; Regidor-Cerrillo, J. Integrative transcriptome and proteome analyses define marked differences between Neospora caninum isolates throughout the tachyzoite lytic cycle. J. Proteom. 2018, 180, 108–119. [Google Scholar] [CrossRef]
- Hemphill, A.; Gottstein, B.; Kaufmann, H. Adhesion and invasion of bovine endothelial cells by Neospora caninum. Parasitology 1996, 112 (Pt 2), 183–197. [Google Scholar] [CrossRef]
- Müller, J.; Balmer, V.; Winzer, P.; Rahman, M.; Manser, V.; Haynes, R.K.; Hemphill, A. In vitro effects of new artemisinin derivatives in Neospora caninum-infected human fibroblasts. Int. J. Antimicrob. Agents 2015, 46, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Nash, T.E.; Luján, H.T.; Mowatt, M.R.; Conrad, J.T. Variant-specific surface protein switching in Giardia lamblia. Infect. Immun. 2001, 69, 1922–1923. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Reid, A.J.; Vermont, S.J.; Cotton, J.A.; Harris, D.; Hill-Cawthorne, G.A.; Konen-Waisman, S.; Latham, S.M.; Mourier, T.; Norton, R.; Quail, M.A.; et al. Comparative genomics of the apicomplexan parasites Toxoplasma gondii and Neospora caninum: Coccidia differing in host range and transmission strategy. PLoS Pathog. 2012, 8, e1002567. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Narabu, S.; Yanai, Y.; Hatano, Y.; Ito, A.; Imai, S.; Ike, K. Gene cloning and characterization of the protein encoded by the Neospora caninum bradyzoite-specific antigen gene BAG1. J. Parasitol. 2013, 99, 453–458. [Google Scholar] [CrossRef]
- Risco-Castillo, V.; Fernandez-Garcia, A.; Zaballos, A.; Aguado-Martinez, A.; Hemphill, A.; Rodriguez-Bertos, A.; Alvarez-Garcia, G.; Ortega-Mora, L.M. Molecular characterisation of BSR4, a novel bradyzoite-specific gene from Neospora caninum. Int. J. Parasitol. 2007, 37, 887–896. [Google Scholar] [CrossRef]
- Wasmuth, J.D.; Pszenny, V.; Haile, S.; Jansen, E.M.; Gast, A.T.; Sher, A.; Boyle, J.P.; Boulanger, M.J.; Parkinson, J.; Grigg, M.E. Integrated bioinformatic and targeted deletion analyses of the SRS gene superfamily identify SRS29C as a negative regulator of Toxoplasma virulence. mBio 2012, 3. [Google Scholar] [CrossRef]
- Bengoa-Luoni, S.A.; Corigliano, M.G.; Sanchez-Lopez, E.; Albarracin, R.M.; Legarralde, A.; Ganuza, A.; Clemente, M.; Sander, V.A. The potential of a DIVA-like recombinant vaccine composed by rNcSAG1 and rAtHsp81.2 against vertical transmission in a mouse model of congenital neosporosis. Acta Trop. 2019, 198, 105094. [Google Scholar] [CrossRef]
- Cannas, A.; Naguleswaran, A.; Muller, N.; Eperon, S.; Gottstein, B.; Hemphill, A. Vaccination of mice against experimental Neospora caninum infection using NcSAG1- and NcSRS2-based recombinant antigens and DNA vaccines. Parasitology 2003, 126, 303–312. [Google Scholar] [CrossRef]
- Liao, M.; Zhang, S.; Xuan, X.; Zhang, G.; Huang, X.; Igarashi, I.; Fujisaki, K. Development of rapid immunochromatographic test with recombinant NcSAG1 for detection of antibodies to Neospora caninum in cattle. Clin. Diagn. Lab. Immunol. 2005, 12, 885–887. [Google Scholar] [CrossRef]
- Moraveji, M.; Hosseini, A.; Moghaddar, N.; Namavari, M.M.; Eskandari, M.H. Development of latex agglutination test with recombinant NcSAG1 for the rapid detection of antibodies to Neospora caninum in cattle. Vet. Parasitol. 2012, 189, 211–217. [Google Scholar] [CrossRef]
- Zhou, M.; Cao, S.; Sevinc, F.; Sevinc, M.; Ceylan, O.; Liu, M.; Wang, G.; Moumouni, P.F.; Jirapattharasate, C.; Suzuki, H.; et al. Enzyme-linked immunosorbent assays using recombinant TgSAG2 and NcSAG1 to detect Toxoplasma gondii and Neospora caninum-specific antibodies in domestic animals in Turkey. J. Vet. Med Sci. Jpn. Soc. Vet. Sci. 2017, 78, 1877–1881. [Google Scholar] [CrossRef] [PubMed]
- Vonlaufen, N.; Müller, N.; Keller, N.; Naguleswaran, A.; Bohne, W.; McAllister, M.M.; Bjorkman, C.; Muller, E.; Caldelari, R.; Hemphill, A. Exogenous nitric oxide triggers Neospora caninum tachyzoite-to-bradyzoite stage conversion in murine epidermal keratinocyte cell cultures. Int. J. Parasitol. 2002, 32, 1253–1265. [Google Scholar] [CrossRef]
- Aguado-Martinez, A.; Ortega-Mora, L.M.; Alvarez-Garcia, G.; Rodriguez-Marco, S.; Risco-Castillo, V.; Marugan-Hernandez, V.; Fernandez-Garcia, A. Stage-specific expression of Nc SAG4 as a marker of chronic Neospora caninum infection in a mouse model. Parasitology 2009, 136, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Garcia, A.; Risco-Castillo, V.; Zaballos, A.; Alvarez-Garcia, G.; Ortega-Mora, L.M. Identification and molecular cloning of the Neospora caninum SAG4 gene specifically expressed at bradyzoite stage. Mol. Biochem. Parasitol. 2006, 146, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Marugan-Hernandez, V.; Ortega-Mora, L.M.; Aguado-Martinez, A.; Jimenez-Ruiz, E.; Alvarez-Garcia, G. Transgenic Neospora caninum strains constitutively expressing the bradyzoite NcSAG4 protein proved to be safe and conferred significant levels of protection against vertical transmission when used as live vaccines in mice. Vaccine 2011, 29, 7867–7874. [Google Scholar] [CrossRef]
- Fuchs, N.; Sonda, S.; Gottstein, B.; Hemphill, A. Differential expression of cell surface- and dense granule-associated Neospora caninum proteins in tachyzoites and bradyzoites. J. Parasitol. 1998, 84, 753–758. [Google Scholar] [CrossRef]
- Schares, G.; Zinecker, C.F.; Schmidt, J.; Azzouz, N.; Conraths, F.J.; Gerold, P.; Schwarz, R.T. Structural analysis of free and protein-bound glycosyl-phosphatidylinositols of Neospora caninum. Mol. Biochem. Parasitol. 2000, 105, 155–161. [Google Scholar] [CrossRef]
- Bezerra, M.A.; Pereira, L.M.; Bononi, A.; Biella, C.A.; Baroni, L.; Pollo-Oliveira, L.; Yatsuda, A.P. Constitutive expression and characterization of a surface SRS (NcSRS67) protein of Neospora caninum with no orthologue in Toxoplasma gondii. Parasitol. Int. 2017, 66, 173–180. [Google Scholar] [CrossRef]
- Bezerra, M.A.; Pereira, L.M.; Baroni, L.; Yatsuda, A.P. The soluble fraction of Neospora caninum treated with PI-PLC is dominated by NcSRS29B and NcSRS29C. Exp. Parasitol. 2019, 204, 107731. [Google Scholar] [CrossRef]
- Pollo-Oliveira, L.; Post, H.; Acencio, M.L.; Lemke, N.; van den Toorn, H.; Tragante, V.; Heck, A.J.; Altelaar, A.F.; Yatsuda, A.P. Unravelling the Neospora caninum secretome through the secreted fraction (ESA) and quantification of the discharged tachyzoite using high-resolution mass spectrometry-based proteomics. Parasites Vectors 2013, 6, 335. [Google Scholar] [CrossRef]
- Sugi, T.; Ma, Y.F.; Tomita, T.; Murakoshi, F.; Eaton, M.S.; Yakubu, R.; Han, B.; Tu, V.; Kato, K.; Kawazu, S.; et al. Toxoplasma gondii cyclic AMP-Dependent protein kinase subunit 3 Is Involved in the switch from tachyzoite to bradyzoite development. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Agop-Nersesian, C.; Naissant, B.; Ben Rached, F.; Rauch, M.; Kretzschmar, A.; Thiberge, S.; Menard, R.; Ferguson, D.J.; Meissner, M.; Langsley, G. Rab11A-controlled assembly of the inner membrane complex is required for completion of apicomplexan cytokinesis. PLoS Pathog. 2009, 5, e1000270. [Google Scholar] [CrossRef] [PubMed]
- Deitsch, K.W.; Dzikowski, R. Variant gene expression and antigenic variation by malaria parasites. Annu. Rev. Microbiol. 2017, 71, 625–641. [Google Scholar] [CrossRef] [PubMed]
- McCulloch, R.; Cobbold, C.A.; Figueiredo, L.; Jackson, A.; Morrison, L.J.; Mugnier, M.R.; Papavasiliou, N.; Schnaufer, A.; Matthews, K. Emerging challenges in understanding trypanosome antigenic variation. Emerg. Top. Life Sci. 2017, 1, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Borst, P. Antigenic variation and allelic exclusion. Cell 2002, 109, 5–8. [Google Scholar] [CrossRef]
- Gargantini, P.R.; Serradell, M.D.C.; Rios, D.N.; Tenaglia, A.H.; Lujan, H.D. Antigenic variation in the intestinal parasite Giardia lamblia. Curr. Opin. Microbiol. 2016, 32, 52–58. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Rippey, N.S.; Toivio-Kinnucan, M.A.; Blagburn, B.L. Ultrastructural effects of diclazuril against Toxoplasma gondii and investigation of a diclazuril-resistant mutant. J. Parasitol. 1995, 81, 459–466. [Google Scholar] [CrossRef]
- Stock, M.; Elazab, S.; Hsu, W. Review of triazine anti-protozoal drugs used in veterinary medicine. J. Vet. Pharmacol. Ther. 2017, 41, 184–194. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Annotation | Accession No | Primer | Sequence |
|---|---|---|---|
| Nc28SrRNA | L49389.1 | Nc28S_F | TCTCTCTCACCAGGTTTAGG |
| Nc28S_R | CCGTGTTTCAAGACGGGTC | ||
| Bradyzoite antigen, putative (BAG1) | NCLIV_027470 | NcBAG1_F | CTCGACTTCATGGATGAGG |
| NcBAG1_R | CTTCTATGGTAACGTCATCC | ||
| Calcium dependent protein kinase 1 | NCLIV_011980 | NcCDPKI_F | AGACGCTGCTATCGCGGG |
| NcCDPKI_R | TTAGTTTCCGCAAAGCTTCAG | ||
| Inner membrane protein 1 | NCLIV_031770 | IMC1_F | GGATGGGACGGCGGTG |
| IMC1_R | CGCCACCGGCTGGATG | ||
| SRS-domain containing protein | NCLIV_019580 | Nc019580_F | CGCGTGCTCGGCGTGG |
| Nc019580_R | GTGCTACCAGTTTGCATCC |
| Parameter | Number |
|---|---|
| Unique peptides | 207041 |
| Non-redundant proteins | 2558 |
| Higher in tachyzoites | 1325 |
| Higher in MNCs | 12 |
| Similar levels in both | 1221 |
| Accession No | Annotation | Protein Features and Properties |
|---|---|---|
| 001830 | Hypothetical protein | SP; homologies to IF2 and collagen. |
| 010030 | Bradyzoite surface protein BSR4 | SP; 2 SRS domains |
| 010050 | SRS domain-containing protein | 2 SRS domains; homologies to 010030 |
| 019520 | MGC83258 protein, related | 1.7 MDa protein; homologies to ubiquitin ligase and mucin |
| 019580 | SRS domain-containing protein | SP, SRS domain, 2 TM domains, 1 lipoprotein domain, homology to T. gondii, bradyzoite surface antigen. Annotated as SAG4. |
| 022800 | Conserved hypothetical protein | Homologies to serine-rich adhesion and collagen |
| 035375 | SRS domain-containing protein | SP, 2 SRS domains |
| 044320 | Zinc finger protein 467, related | Homologies to serine-rich adhesins |
| 044430 | Hypothetical protein | Short sequences with homologies to zinc finger proteins and ATP-ase domain of Hsp90 |
| 061850 | SRS domain-containing protein | SP, TM domain, 4 SRS domains. |
| 069001 | SRS domain-containing protein | SP, 2 SRS domains. |
| 069110 | Hypothetical protein | SP |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winzer, P.; Müller, J.; Imhof, D.; Ritler, D.; Uldry, A.-C.; Braga-Lagache, S.; Heller, M.; Ojo, K.K.; Van Voorhis, W.C.; Ortega-Mora, L.-M.; et al. Neospora caninum: Differential Proteome of Multinucleated Complexes Induced by the Bumped Kinase Inhibitor BKI-1294. Microorganisms 2020, 8, 801. https://doi.org/10.3390/microorganisms8060801
Winzer P, Müller J, Imhof D, Ritler D, Uldry A-C, Braga-Lagache S, Heller M, Ojo KK, Van Voorhis WC, Ortega-Mora L-M, et al. Neospora caninum: Differential Proteome of Multinucleated Complexes Induced by the Bumped Kinase Inhibitor BKI-1294. Microorganisms. 2020; 8(6):801. https://doi.org/10.3390/microorganisms8060801
Chicago/Turabian StyleWinzer, Pablo, Joachim Müller, Dennis Imhof, Dominic Ritler, Anne-Christine Uldry, Sophie Braga-Lagache, Manfred Heller, Kayode K. Ojo, Wesley C. Van Voorhis, Luis-Miguel Ortega-Mora, and et al. 2020. "Neospora caninum: Differential Proteome of Multinucleated Complexes Induced by the Bumped Kinase Inhibitor BKI-1294" Microorganisms 8, no. 6: 801. https://doi.org/10.3390/microorganisms8060801
APA StyleWinzer, P., Müller, J., Imhof, D., Ritler, D., Uldry, A.-C., Braga-Lagache, S., Heller, M., Ojo, K. K., Van Voorhis, W. C., Ortega-Mora, L.-M., & Hemphill, A. (2020). Neospora caninum: Differential Proteome of Multinucleated Complexes Induced by the Bumped Kinase Inhibitor BKI-1294. Microorganisms, 8(6), 801. https://doi.org/10.3390/microorganisms8060801