Experimental Infection of Dogs with Toscana Virus and Sandfly Fever Sicilian Virus to Determine Their Potential as Possible Vertebrate Hosts


 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
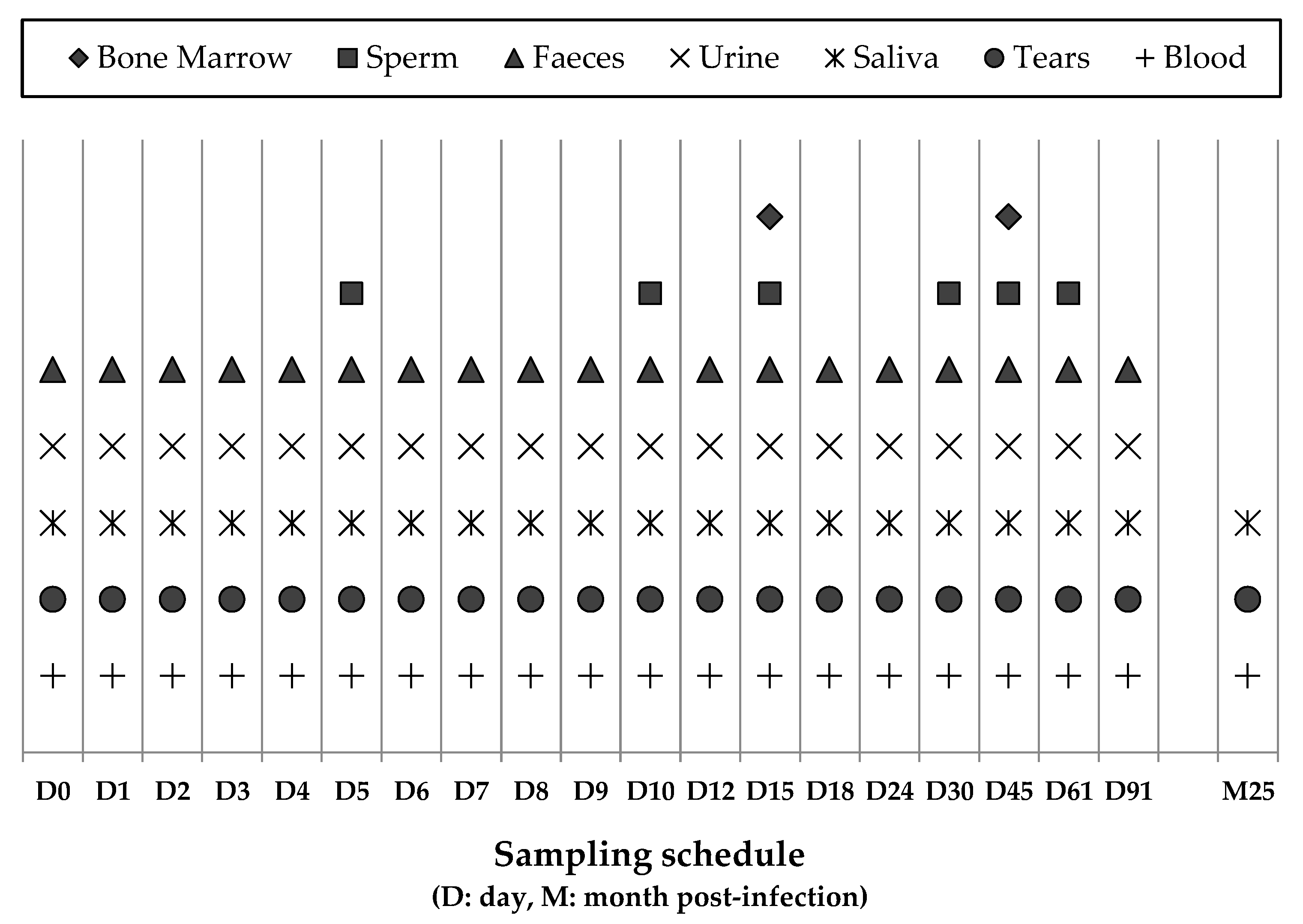
2.1. Experimental Design, Sampling, and Clinical Examination of Dogs
2.2. RT-qPCR Diagnosis
2.3. Sensitivity and Limit of Detection (LoD)
2.4. Neutralizing Antibody Detection
2.5. Statistical Methods
3. Results
3.1. Clinical, Hematological and Biochemical Responses of Dogs Inoculated with SFSV or TOSV
3.2. TOSV and SFSV Detection and Viral Loads
3.3. Virus Neutralizing Antibodies in Dogs Inoculated with TOSV or SFSV
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alkan, C.; Bichaud, L.; de Lamballerie, X.; Alten, B.; Gould, E.A.; Charrel, R.N. Sandfly-borne phleboviruses of Eurasia and Africa: Epidemiology, genetic diversity, geographic range, control measures. Antivir. Res. 2013, 100, 54–74. [Google Scholar] [CrossRef]
- Sabin, A.B. Experimental studies on Phlebotomus (pappataci, sandfly) fever during World War II. Arch. für die Gesamte Virusforsch. 1951, 4, 367–410. [Google Scholar] [CrossRef]
- Ayhan, N.; Charrel, R.N. Of phlebotomines (sandflies) and viruses: A comprehensive perspective on a complex situation. Curr. Opin. Insect Sci. 2017, 22, 117–124. [Google Scholar] [CrossRef]
- De Ory, F.; Gegúndez, M.I.; Fedele, C.G.; Sánchez-Seco, M.P. Virus Toscana, West Nile y de la coriomeningitis linfocitaria como causantes de meningitis aséptica en España. Med. Clínica 2009, 132, 587–590. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Garcia, F.A.; Moreno-Docon, A.; Lopez-Lopez, M.; Albert-Lacal, L.; Martinez-Toldos, M.C.; Segovia-Hernandez, M.; Fernandez-Barreiro, A. A case of meningitis due to Toscana virus in Murcia. Rev. Neurol. 2007, 45, 317–318. [Google Scholar]
- Nougairede, A.; Bichaud, L.; Thiberville, S.D.; Ninove, L.; Zandotti, C.; de Lamballerie, X.; Brouqui, P.; Charrel, R.N. Isolation of Toscana Virus from the Cerebrospinal Fluid of a Man with Meningitis in Marseille, France. Vector Borne Zoonotic Dis. 2010, 13, 685–688. [Google Scholar] [CrossRef]
- Moriconi, M.; Rugna, G.; Calzolari, M.; Bellini, R.; Albieri, A.; Angelini, P.; Cagarelli, R.; Landini, M.P.; Charrel, R.N.; Varani, S. Phlebotomine sand fly–borne pathogens in the Mediterranean Basin: Human leishmaniasis and phlebovirus infections. PLoS Negl. Trop. Dis. 2017, 11, e0005660. [Google Scholar] [CrossRef] [PubMed]
- Alwassouf, S.; Maia, C.; Ayhan, N.; Coimbra, M.; Cristovao, J.M.; Richet, H.; Bichaud, L.; Campino, L.; Charrel, R.N. Neutralization-based seroprevalence of Toscana virus and sandfly fever Sicilian virus in dogs and cats from Portugal. J. Gen. Virol. 2016, 97, 2816–2823. [Google Scholar] [CrossRef] [PubMed]
- Alwassouf, S.; Christodoulou, V.; Bichaud, L.; Ntais, P.; Mazeris, A.; Antoniou, M.; Charrel, R.N. Seroprevalence of Sandfly-Borne Phleboviruses Belonging to Three Serocomplexes (Sandfly fever Naples, Sandfly fever Sicilian and Salehabad) in Dogs from Greece and Cyprus Using Neutralization Test. PLoS Negl. Trop. Dis. 2016, 10, e0005063. [Google Scholar] [CrossRef] [PubMed]
- Dahmani, M.; Alwassouf, S.; Grech-Angelini, S.; Marié, J.L.; Davoust, B.; Charrel, R.N. Seroprevalence of Toscana virus in dogs from Corsica, France. Parasites Vectors 2016, 9, 381. [Google Scholar] [CrossRef] [PubMed]
- Maia, C.; Alwassouf, S.; Cristóvão, J.M.; Ayhan, N.; Pereira, A.; Charrel, R.N.; Campino, L. Serological association between Leishmania infantum and sand fly fever Sicilian (but not Toscana) virus in sheltered dogs from southern Portugal. Parasites Vectors 2017, 10, 92. [Google Scholar] [CrossRef] [PubMed]
- Sakhria, S.; Alwassouf, S.; Fares, W.; Bichaud, L.; Dachraoui, K.; Alkan, C.; Zoghlami, Z.; de Lamballerie, X.; Zhioua, E.; Charrel, R.N. Presence of sandfly-borne phleboviruses of two antigenic complexes (Sandfly fever Naples virus and Sandfly fever Sicilian virus) in two different bio-geographical regions of Tunisia demonstrated by a microneutralisation-based seroprevalence study in dogs. Parasites Vectors 2014, 7, 476. [Google Scholar] [PubMed]
- Tahir, D.; Alwassouf, S.; Loudahi, A.; Davoust, B.; Charrel, R.N. Seroprevalence of Toscana virus in dogs from Kabylia (Algeria). Clin. Microbiol. Infect 2016, 22, e16–e17. [Google Scholar] [CrossRef] [PubMed]
- Alvar, J.; Canavate, C.; Molina, R.; Moreno, J.; Nieto, J. Canine leishmaniasis. Adv. Parasitol. 2004, 57, 1–88. [Google Scholar] [PubMed]
- Navarro-Mari, J.M.; Palop-Borras, B.; Pérez-Ruiz, M.; Sanbonmatsu-Gámez, S. Serosurvey Study of Toscana Virus in Domestic Animals, Granada, Spain. Vector Borne Zoonotic Dis. 2011, 11, 583–587. [Google Scholar] [CrossRef] [PubMed]
- Dincer, E.; Gargari, S.; Ozkul, A.; Ergunay, K. Potential animal reservoirs of Toscana virus and coinfections with Leishmania infantum in Turkey. Am. J. Trop. Med. Hyg. 2015, 92, 690–697. [Google Scholar] [CrossRef]
- Weidmann, M.; Sanchez-Seco, M.P.; Sall, A.A.; Ly, P.O.; Thiongane, Y.; Lô, M.M.; Schley, H.; Hufert, F.T. Rapid detection of important human pathogenic Phleboviruses. J. Clin. Virol. 2008, 41, 138–142. [Google Scholar] [CrossRef]
- Mackay, I.M. Real-time PCR in Microbiology: From Diagnosis to Characterization; Caister Academic Press: Norfolk, UK, 2007; p. 454. [Google Scholar]
- Sakhria, S.; Bichaud, L.; Mensi, M.; Salez, N.; Dachraoui, K.; Thirion, L.; Cherni, S.; Chelbi, I.; De Lamballerie, X.; Zhioua, E.; et al. Co-Circulation of Toscana Virus and Punique Virus in Northern Tunisia: A Microneutralisation-Based Seroprevalence Study. PLoS Negl. Trop. Dis. 2013, 7, e2429. [Google Scholar] [CrossRef]
- Watts, D.M.; MacDonald, C.; Bailey, C.L.; Meegan, J.M.; Peters, C.J.; McKee, K.T., Jr. Experimental infection of Phlebotomus papatasi with sand fly fever Sicilian virus. Am. J. Trop. Med. Hyg. 1988, 39, 611–616. [Google Scholar] [CrossRef]
- Bartelloni, P.J.; Tesh, R.B. Clinical and serologic responses of volunteers infected with phlebotomus fever virus (Sicilian type). Am. J. Trop. Med. Hyg. 1976, 25, 456–462. [Google Scholar] [CrossRef]
- Charrel, R.N.; Gallian, P.; Navarro-Marí, J.M.; Nicoletti, L.; Papa, A.; Sánchez-Seco, M.P.; Tenorio, A.; De Lamballerie, X. Emergence of Toscana virus in Europe. Emerg. Infect. Dis. 2005, 11, 1657–1663. [Google Scholar] [CrossRef]
- Charrel, R.N. Toscana Virus Infection. Emerging Infectious Diseases: Clinical Case Studies; Ergönül, Ö., Can, F., Madoff, L., Akova, M., Eds.; Academic Press: New York, NY, USA, 2014; pp. 111–119. [Google Scholar]
- Tesh, R.B.; Lubroth, J.; Guzman, H. Simulation of arbovirus overwintering: Survival of Toscana virus (Bunyaviridae: Phlebovirus) in its natural sand fly vector Phlebotomus perniciosus. Am. J. Trop. Med. Hyg. 1992, 47, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Cusi, M.G.; Gandolfo, C.; Terrosi, C.; Savellini, G.G.; Belmonte, G.; Miracco, C. Toscana virus infects dendritic and endothelial cells opening the way for the central nervous system. J. Neurovirol. 2016, 22, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Cusi, M.G.; Savellini, G.G.; Terrosi, C.; Di Genova, G.; Valassina, M.; Valentini, M.; Bartolommei, S.; Miracco, C. Development of a mouse model for the study of Toscana virus pathogenesis. Virology 2005, 333, 66–73. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Dog # | Virus | Infection Dose | Age (Months) at D0 | Body Temperature Range (°C) | Body Weight (kg) at the Start of the Experiment (D0) | Body Weight (kg) at 91 Days Postinoculation (dpi) |
|---|---|---|---|---|---|---|
| 10011 | TOSV 1 | High 3 | 10 | 37.8–38.8 | 14.2 | 15.2 |
| 10326 | TOSV | High | 20 | 38.3–39.3 | 17.5 | 17.5 |
| 10130 | TOSV | Low 4 | 10 | 38.2–38.8 | 15.8 | 17.4 |
| 10325 | TOSV | Low | 6 | 38.1–38.9 | 17.9 | 21.5 |
| 10092 | TOSV | None 5 | 10 | 38.4–39.4 | 13.3 | 16.5 |
| 10095 | SFSV 2 | High | 10 | 38.0–39.4 | 19.3 | 21.1 |
| 10096 | SFSV | High | 10 | 38.2–38.9 | 15.2 | 17.0 |
| 10323 | SFSV | Low | 7 | 38.4–39.1 | 11.3 | 13.3 |
| 10324 | SFSV | Low | 7 | 37.8–39.3 | 11.6 | 15.1 |
| 10129 | SFSV | None | 10 | 37.9–38.7 | 13.6 | 15.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñoz, C.; Ayhan, N.; Ortuño, M.; Ortiz, J.; Gould, E.A.; Maia, C.; Berriatua, E.; Charrel, R.N. Experimental Infection of Dogs with Toscana Virus and Sandfly Fever Sicilian Virus to Determine Their Potential as Possible Vertebrate Hosts. Microorganisms 2020, 8, 596. https://doi.org/10.3390/microorganisms8040596
Muñoz C, Ayhan N, Ortuño M, Ortiz J, Gould EA, Maia C, Berriatua E, Charrel RN. Experimental Infection of Dogs with Toscana Virus and Sandfly Fever Sicilian Virus to Determine Their Potential as Possible Vertebrate Hosts. Microorganisms. 2020; 8(4):596. https://doi.org/10.3390/microorganisms8040596
Chicago/Turabian StyleMuñoz, Clara, Nazli Ayhan, Maria Ortuño, Juana Ortiz, Ernest A. Gould, Carla Maia, Eduardo Berriatua, and Remi N. Charrel. 2020. "Experimental Infection of Dogs with Toscana Virus and Sandfly Fever Sicilian Virus to Determine Their Potential as Possible Vertebrate Hosts" Microorganisms 8, no. 4: 596. https://doi.org/10.3390/microorganisms8040596
APA StyleMuñoz, C., Ayhan, N., Ortuño, M., Ortiz, J., Gould, E. A., Maia, C., Berriatua, E., & Charrel, R. N. (2020). Experimental Infection of Dogs with Toscana Virus and Sandfly Fever Sicilian Virus to Determine Their Potential as Possible Vertebrate Hosts. Microorganisms, 8(4), 596. https://doi.org/10.3390/microorganisms8040596