The Response to Oxidative Stress in Listeria monocytogenes Is Temperature Dependent

 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial and Culture Conditions
2.2. Oxidative Stress Assay
2.3. RNA Extraction and Gene Expression
2.4. Statistical Analysis
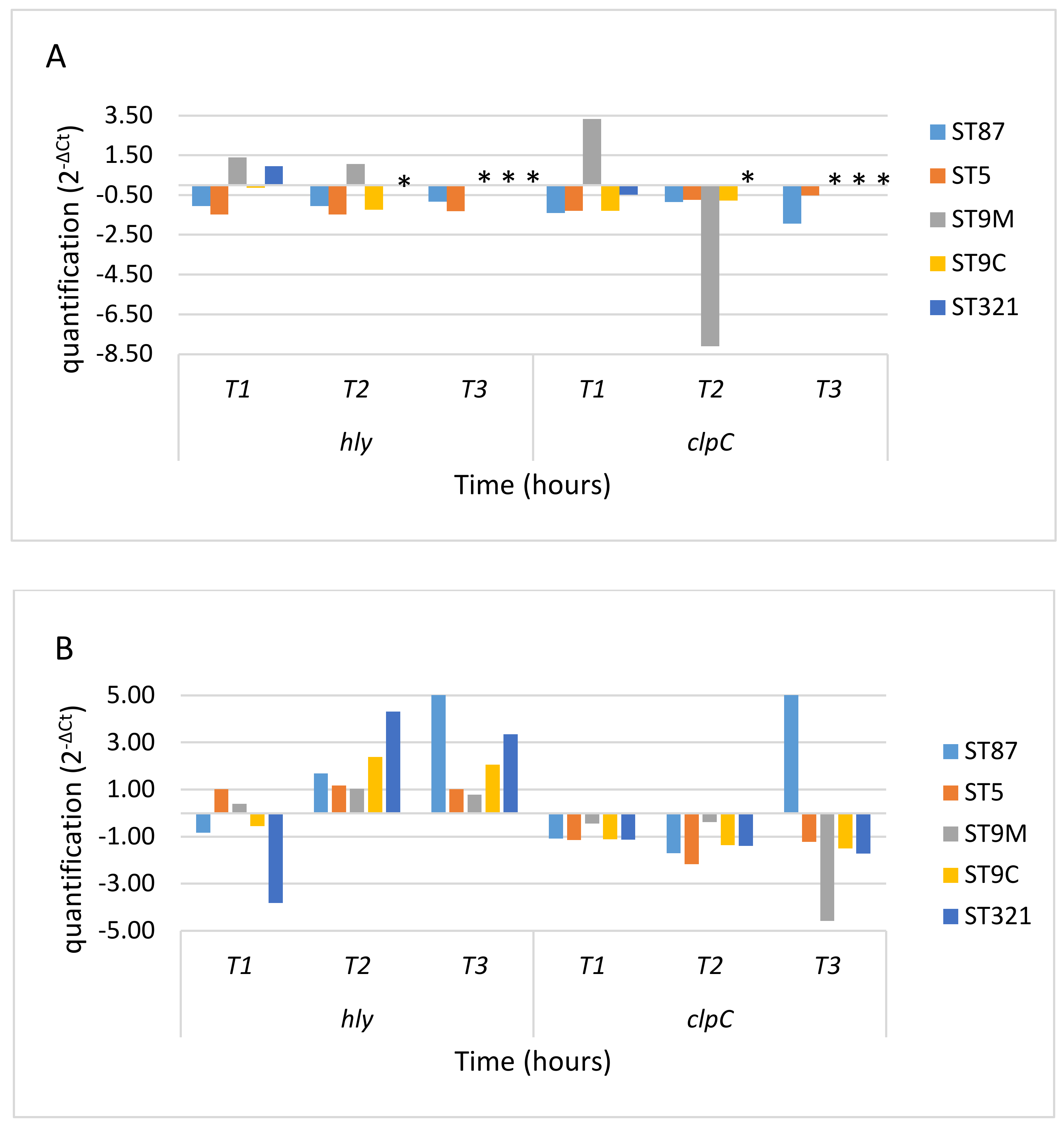
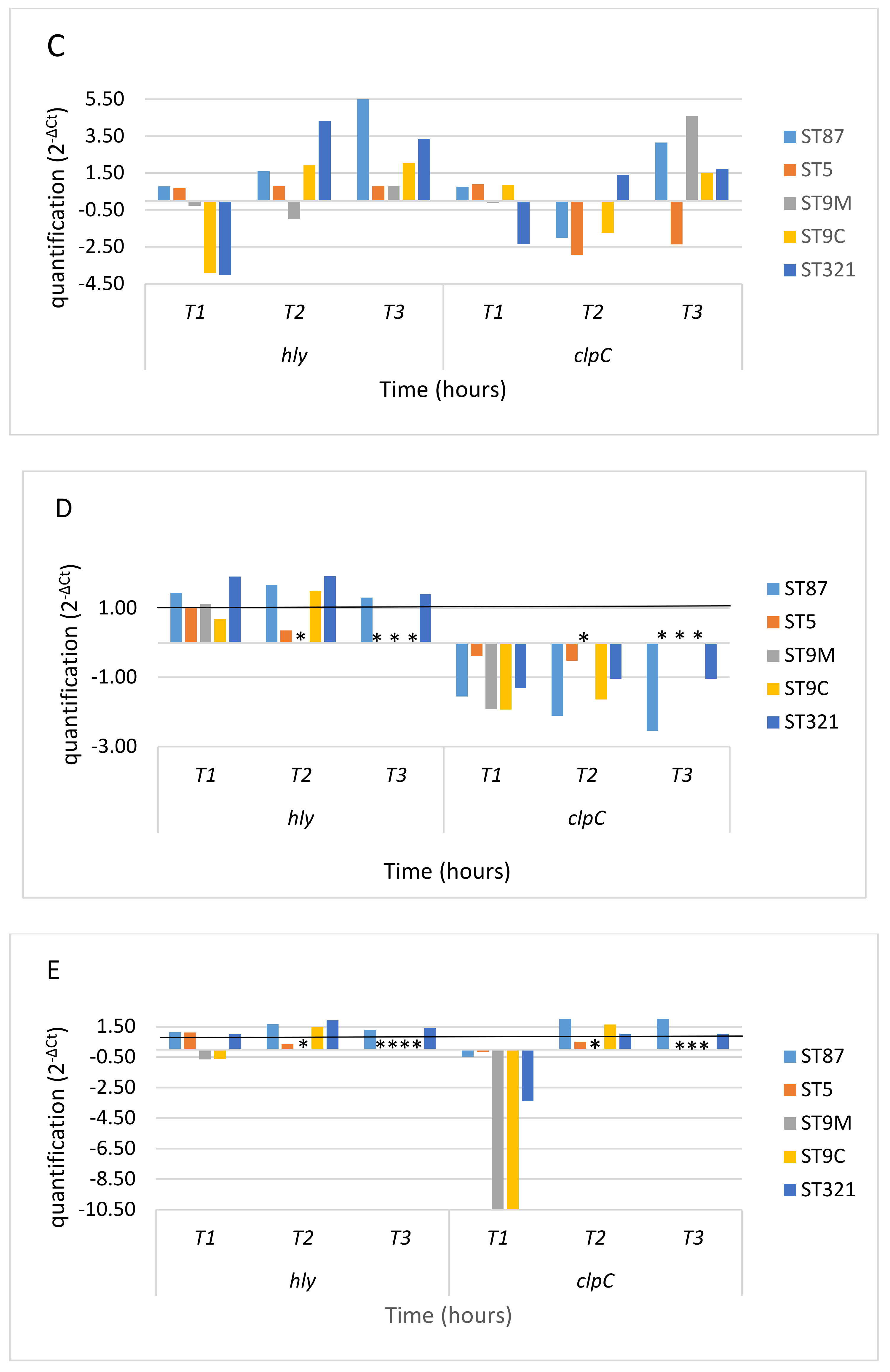
3. Results
3.1. Response to Oxidative Stress
3.2. Gene Expression in Oxidative Stress Conditions
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Ferreira, V.; Wiedmann, M.; Teixeira, P.; Stasiewicz, M.J. Listeria monocytogenes persistence in food-associated environments: Epidemiology, strain characteristics, and implications for public health. J. Food Prot. 2014, 77, 150–170. [Google Scholar] [CrossRef] [PubMed]
- Bergholz, T.M.; Shah, M.K.; Burall, L.S.; Rakic-Martinez, M.; Datta, A.R. Genomic and phenotypic diversity of Listeria monocytogenes clonal complexes associated with human listeriosis. Appl. Microbiol. Biotechnol. 2018, 102, 3475–3485. [Google Scholar] [CrossRef]
- Melero, B.; Stessl, B.; Manso, B.; Wagner, M.; Esteban-Carbonero, Ó.J.; Hernández, M.; Rovira, J.; Rodriguez-Lázaro, D. Listeria monocytogenes colonization in a newly established dairy processing facility. Int. J. Food Microbiol. 2019, 289, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Braschi, G.; Serrazanetti, D.I.; Siroli, L.; Patrignani, F.; De Angelis, M.; Lanciotti, R. Gene expression responses of Listeria monocytogenes Scott A exposed to sub-lethal concentrations of natural antimicrobials. Int. J. Food Microbiol. 2018, 286, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.A.; Alves, Â.; Ferreira, V.; Teixeira, P.C.M. The impact of environmental stresses in the virulence traits of Listeria monocytogenes relevant to food safety. In Listeria Monocytogenes; IntechOpen: London, UK, 2018. [Google Scholar]
- Tezel, U.; Pavlostathis, S.G. Quaternary ammonium disinfectants: Microbial adaptation, degradation and ecology. Curr. Opin. Biotechnol. 2015, 33, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Soumet, C.; Ragimbeau, C.; Maris, P. Screening of benzalkonium chloride resistance in Listeria monocytogenes strains isolated during cold smoked fish production. Lett. Appl. Microbiol. 2005, 41, 291–296. [Google Scholar] [CrossRef]
- Yun, H.S.; Kim, Y.; Oh, S.; Jeon, W.M.; Frank, J.F.; Kim, S.H. Susceptibility of Listeria monocytogenes biofilms and planktonic cultures to hydrogen peroxide in food processing environments. Biosci. Biotechnol. Biochem. 2012, 76, 2008–2013. [Google Scholar] [CrossRef]
- Harter, E.; Wagner, E.M.; Zaiser, A.; Halecker, S.; Wagner, M.; Rychli, K. The novel stress survival islet 2 (SSI-2), predominantly present in Listeria monocytogenes strains of ST121, is involved in alkaline and oxidative stress response. Appl. Environ. Microbiol. 2017, 83, e00827-17. [Google Scholar] [CrossRef]
- Freitag, N.E.; Port, G.C.; Miner, M.D. Listeria monocytogenes from saprophyte to intracellular pathogen. Nat. Rev. Microbiol. 2009, 7, 623. [Google Scholar] [CrossRef]
- Markkula, A.; Lindström, M.; Johansson, P.; Björkroth, J.; Korkeala, H. Role of four putative DEAD-box RNA helicase genes in growth of Listeria monocytogenes EGD-e under heat, pH, osmotic, ethanol, and oxidative stresses. Appl. Environ. Microbiol. 2012, 78, 6875–6882. [Google Scholar] [CrossRef]
- Suo, Y.; Huang, Y.; Liu, Y.; Shi, C.; Shi, X. The expression of superoxide dismutase (SOD) and a putative ABC transporter permease is inversely correlated during biofilm formation in Listeria monocytogenes 4b G. PLoS ONE 2012, 7, e48467. [Google Scholar] [CrossRef] [PubMed]
- Van der Veen, S.; Abee, T. Contribution of Listeria monocytogenes RecA to acid and bile survival and invasion of human intestinal Caco-2 cells. Int. J. Med. Microbiol. 2011, 301, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, M.; Goebel, W. Molecular virulence determinants of Listeria monocytogenes. Food Sci. Technol. 2017, 161, 111. [Google Scholar]
- Komora, N.; Bruschi, C.; Magalhães, R.; Ferreira, V.; Teixeira, P. Survival of Listeria monocytogenes with different antibiotic resistance patterns to food-associated stresses. Int. J. Food Microbiol. 2017, 245, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Manso, B.; Melero, B.; Stessl, B.; Fernández-Natal, I.; Jaime, I.; Hernández, M.; Wagner, M.; Rovira, J.; Rodríguez-Lázaro, D. Characterization of virulence and persistence abilities of Listeria monocytogenes strains isolated from food processing premises. J. Food Prot. 2019, 82, 1922–1930. [Google Scholar] [CrossRef]
- Melero, B.; Manso, B.; Stessl, B.; Hernandez, M.; Wagner, M.; Rovira, J.; Rodriguez-Lazaro, D. Distribution and persistence of Listeria monocytogenes in a heavily contaminated poultry processing facility. J. Food Prot. 2019, 82, 1524–1531. [Google Scholar] [CrossRef]
- Ciolacu, L.; Nicolau, A.I.; Wagner, M.; Rychli, K. Listeria monocytogenes isolated from food samples from a Romanian black market show distinct virulence profiles. Int. J. Food Microbiol. 2015, 209, 44–51. [Google Scholar] [CrossRef]
- Rea, R.; Hill, C.; Gahan, C.G. Listeria monocytogenes PerR mutants display a small-colony phenotype, increased sensitivity to hydrogen peroxide, and significantly reduced murine virulence. Appl. Environ. Microbiol. 2005, 71, 8314–8322. [Google Scholar] [CrossRef]
- Huang, Y.; Suo, Y.; Shi, C.; Szlavik, J.; Shi, X.M.; Knøchel, S. Mutations in gltB and gltC reduce oxidative stress tolerance and biofilm formation in Listeria monocytogenes 4b G. Int. J. Food Microbiol. 2013, 163, 223–230. [Google Scholar] [CrossRef]
- Bielecka, M.K. Exploration into the Virulence Mechanisms of Listeria. Ph.D. Thesis, University of Edinburgh, Edinburgh, UK, 2011. [Google Scholar]
- Rodríguez-Lázaro, D.; Pla, M.; Scortti, M.; Mozó, H.J.; Vázquez-Boland, J.A. A novel real—Time PCR for Listeria monocytogenes that monitors analytical performance via an internal amplification control. Appl. Environ. Microbiol. 2015, 71, 9008–9012. [Google Scholar] [CrossRef]
- Kim, H.; Marquis, H.; Boor, K.J. Sigma B contributes to Listeria monocytogenes invasion by controlling expression of inlA and inlB. Microbiology 2015, 151, 3215–3230. [Google Scholar] [CrossRef] [PubMed]
- Abeysundara, P.D.A.; Nannapaneni, R.; Soni, K.A.; Sharma, C.S.; Mahmoud, B. Induction and stability of oxidative stress adaptation in Listeria monocytogenes EGD (Bug600) and F1057 in sublethal concentrations of H2O2 and NaOH. Int. J. Food Microbiol. 2016, 238, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Al-Nabulsi, A.A.; Osaili, T.M.; Shaker, R.R.; Olaimat, A.N.; Jaradat, Z.W.; Elabedeen, N.A.Z.; Holley, R.A. Effects of osmotic pressure, acid, or cold stresses on antibiotic susceptibility of Listeria monocytogenes. Food Microbiol. 2015, 46, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Boura, M.; Keating, C.; Royet, K.; Paudyal, R.; O’Donoghue, B.; O’Byrne, C.P.; Karatzas, K.A. Loss of sigB in Listeria monocytogenes strains EGD-e and 10403S confers hyperresistance to hydrogen peroxide in stationary phase under aerobic conditions. Appl. Environ. Microbiol. 2016, 82, 4584–4591. [Google Scholar] [CrossRef]
- Dons, L.E.; Mosa, A.; Rottenberg, M.E.; Rosenkrantz, J.T.; Kristensson, K.; Olsen, J.E. Role of the Listeria monocytogenes 2-Cys peroxiredoxin homologue in protection against oxidative and nitrosative stress and in virulence. Pathog. Dis. 2014, 70, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, Y.; Yamada, F.; Yoshikawa, Y.; Mochizuki, M.; Takano, T.; Hondo, R.; Ueda, F. Sequential transition of the injury phenotype, temperature-dependent survival and transcriptional response in Listeria monocytogenes following lethal H2O2 exposure. Int. J. Food Microbiol. 2017, 259, 52–58. [Google Scholar] [CrossRef]
- Tasara, T.; Stephan, R. Cold stress tolerance of Listeria monocytogenes: A review of molecular adaptive mechanisms and food safety implications. J. Food Prot. 2016, 69, 1473–1484. [Google Scholar] [CrossRef]
- Ferreira, A.; O’Byrne, C.P.; Boor, K.J. Role of sigma B in heat, ethanol, acid, and oxidative stress resistance and during carbon starvation in Listeria monocytogenes. Appl. Environ. Microbiol. 2001, 67, 4454–4457. [Google Scholar] [CrossRef]
- Ivy, R.A.; Wiedmann, M.; Boor, K.J. Listeria monocytogenes grown at 7 °C shows reduced acid survival and an altered transcriptional response to acid shock compared to L. monocytogenes grown at 37 °C. Appl. Environ. Microbiol. 2012, 8, 3824–3836. [Google Scholar] [CrossRef]
- Huang, Y.; Morvay, A.A.; Shi, X.; Suo, Y.; Shi, C.; Knøchel, S. Comparison of oxidative stress response and biofilm formation of Listeria monocytogenes serotypes 4b and 1/2a. Food Control 2018, 85, 416–422. [Google Scholar] [CrossRef]
- Chastanet, A.; Derre, I.; Nair, S.; Msadek, T. clpB, a novel member of the Listeria monocytogenes CtsR regulon, is involved in virulence but not in general stress tolerance. J. Bacteriol. 2004, 186, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Chaturongakul, S.; Raengpradub, S.; Wiedmann, M.; Boor, K.J. Modulation of stress and virulence in Listeria monocytogenes. Trends Microbiol. 2018, 16, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Azizoglu, R.O.; Kathariou, S. Temperature-dependent requirement for catalase in aerobic growth of Listeria monocytogenes F2365. Appl Environ Microbiol. 2010, 76, 6998–7003. [Google Scholar] [CrossRef] [PubMed]
- Møretrø, T.; Fanebust, H.; Fagerlund, A.; Langsrud, S. Whole room disinfection with hydrogen peroxide mist to control Listeria monocytogenes in food industry related environments. Int. J. Food Microbiol. 2019, 292, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Boland, J.A.; Kuhn, M.; Berche, P.; Chakraborty, T.; Domínguez-Bernal, G.; Goebel, W.; González-Zorn, B.; Wehland, J.; Kreft, J. Listeria pathogenesis and molecular virulence determinants. Clin. Microbiol. Rev. 2001, 14, 584–640. [Google Scholar] [CrossRef] [PubMed]
- Eshwar, A.K.; Guldimann, C.; Oevermann, A.; Tasara, T. Cold-shock domain family proteins (Csps) are involved in regulation of virulence, cellular aggregation, and flagella based motility in Listeria monocytogenes. Front. Cell Infect. Microbiol. 2017, 7, 457. [Google Scholar] [CrossRef]
- Lee, T.; Jun, S.H.; Choi, C.W.; Kim, S.I.; Lee, J.C.; Shin, J.H. Salt stress affects global protein expression profiles of extracellular membrane- derived vesicles of Listeria monocytogenes. Microb. Pathog. 2017, 115, 272–279. [Google Scholar] [CrossRef]
- Nadon, C.A.; Bowen, B.M.; Wiedmann, M.; Boor, K.J. Sigma B contributes to PrfA mediated virulence in Listeria monocytogenes. Infect. Immun. 2002, 70, 3948–3952. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| CHP at 37 °C a | CHP at 10 °C | H2O2 at 10 °C b | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Food Industry | Sample Type | Lineage | Strains | T1 | T2 | T3 | Stn.Error c | T1 | T2 | T3 | Stn.Error | T1 | T2 | T3 | Stn.Error |
| Mean | Mean | Mean | Mean | Mean | Mean | Mean | Mean | Mean | |||||||
| Cheese making factory | Cheese crumbs | I d | ST5 f | 5.70 | 8.35 | 9.13 * | 0.430 | 0.93 | 1.43 | 1.93 | 0.256 | 1.15 | 1.98 | 1.64 | 0.212 |
| Floor | II e | ST7 | 5.61 | 7.49 | 9.05 * | 0.611 | 1.46 | 2.01 | 2.36 | 0.592 | 0.85 | 1.54 | 2.53 | 0.752 | |
| Cheese crumbs | I | ST6 | 4.47 | 7.77 | 8.95 * | 0.148 | 1.95 | 1.60 | 1.96 | 0.597 | 1.59 | 3.47 | 4.43 | 0.691 | |
| Cheese crumbs | II | ST9 # | 6.75 | 8.99 | 9.14 * | 0.106 | 0.75 | 1.64 | 2.24 | 0.237 | 1.39 | 3.64 | 9.15 * | 0.184 | |
| Meat processing plant | Drain | I | ST87 # | 5.02 | 7.45 | 8.20 | 0.831 | 1.37 | 2.12 | 2.77 | 0.130 | 0.11 | 0.68 | 1.20 | 0.150 |
| Boxes | I | ST5 # | 5.17 | 7.57 | 9.00 | 0.251 | 1.19 | 1.67 | 2.35 | 0.098 | −0.04 | 1.85 | 2.92 | 0.627 | |
| Floor | II | ST9 # | 6.07 | 8.95 | 9.25 * | 0.171 | 1.50 | 1.96 | 2.60 | 1.150 | 3.50 | 7.15 | 9.30 * | 1.312 | |
| Wall | II | ST9 | 7.23 | 9.23 * | 9.23 * | 0.301 | 2.09 | 2.46 | 3.18 | 1.061 | 3.79 | 9.12 * | 9.12 * | 0.668 | |
| Floor | I | ST1 | 4.19 | 6.08 | 7.88 | 0.746 | 1.88 | 1.72 | 1.59 | 1.037 | 0.94 | 2.41 | 4.74 | 0.106 | |
| Drain | II | ST199 | 6.42 | 7.97 | 8.71 * | 0.621 | 2.07 | 2.68 | 4.07 | 0.495 | 3.88 | 8.62 * | 8.62 * | 0.114 | |
| Seafood company | Water machine | II | ST321 # | 7.57 | 8.92 * | 8.92 * | 0.679 | 1.57 | 3.48 | 4.43 | 0.549 | 0.97 | 3.71 | 7.97 | 0.600 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manso, B.; Melero, B.; Stessl, B.; Jaime, I.; Wagner, M.; Rovira, J.; Rodríguez-Lázaro, D. The Response to Oxidative Stress in Listeria monocytogenes Is Temperature Dependent. Microorganisms 2020, 8, 521. https://doi.org/10.3390/microorganisms8040521
Manso B, Melero B, Stessl B, Jaime I, Wagner M, Rovira J, Rodríguez-Lázaro D. The Response to Oxidative Stress in Listeria monocytogenes Is Temperature Dependent. Microorganisms. 2020; 8(4):521. https://doi.org/10.3390/microorganisms8040521
Chicago/Turabian StyleManso, Beatriz, Beatriz Melero, Beatrix Stessl, Isabel Jaime, Martin Wagner, Jordi Rovira, and David Rodríguez-Lázaro. 2020. "The Response to Oxidative Stress in Listeria monocytogenes Is Temperature Dependent" Microorganisms 8, no. 4: 521. https://doi.org/10.3390/microorganisms8040521
APA StyleManso, B., Melero, B., Stessl, B., Jaime, I., Wagner, M., Rovira, J., & Rodríguez-Lázaro, D. (2020). The Response to Oxidative Stress in Listeria monocytogenes Is Temperature Dependent. Microorganisms, 8(4), 521. https://doi.org/10.3390/microorganisms8040521