Fungal Microbiota of Sea Buckthorn Berries at Two Ripening Stages and Volatile Profiling of Potential Biocontrol Yeasts

 ,
, 
 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sample Collection
2.3. Metagenomic Analysis of Sea Buckthorn Fungal Microbiota
2.4. Bioinformatics Analysis
2.5. Cultivable Yeast Identification
2.6. Sampling and Identification of Volatile Organic Compounds (VOCs) Produced by Yeasts
2.7. Statistical Data Analysis
3. Results
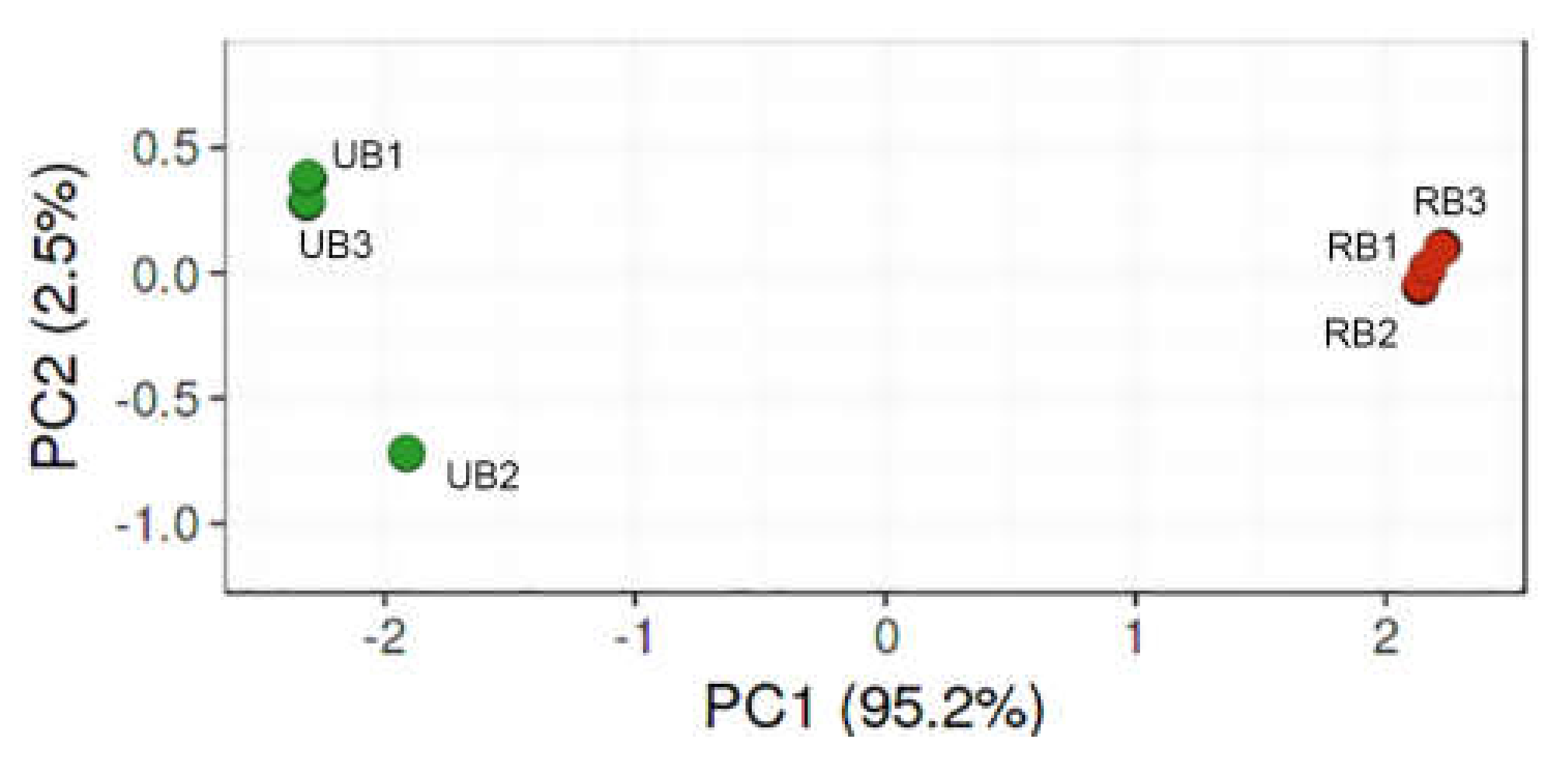
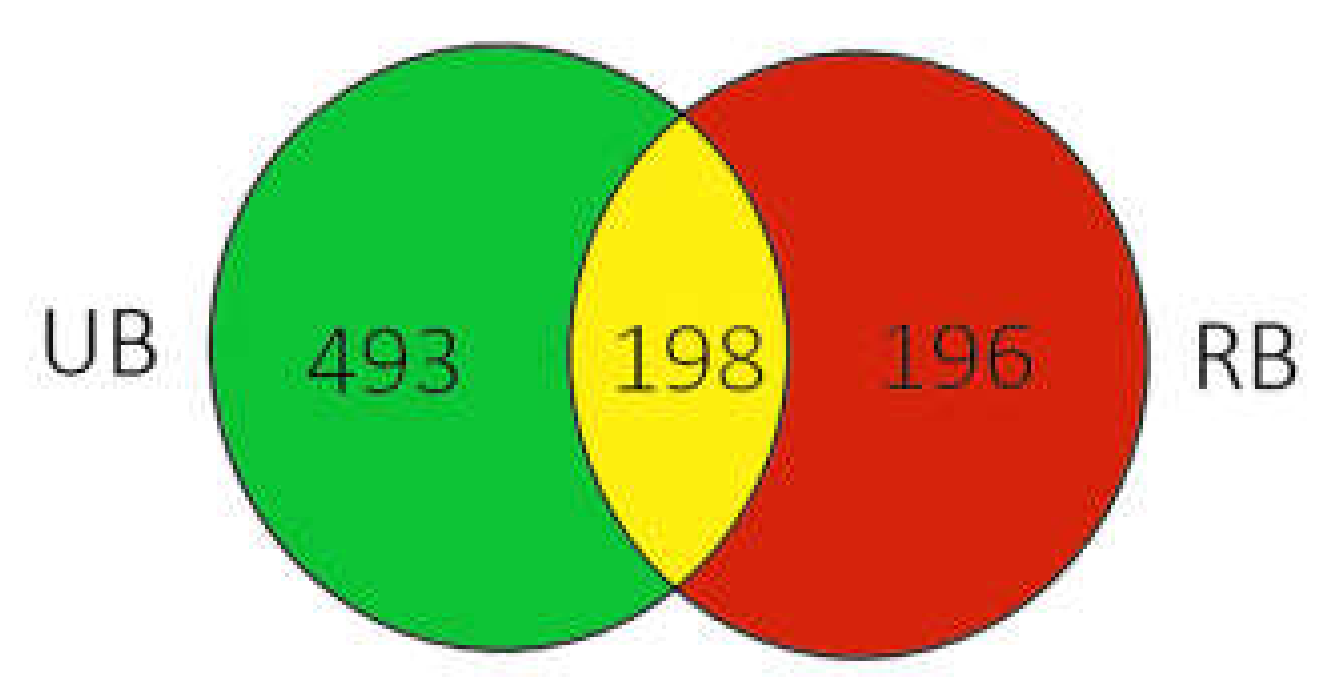
3.1. Diversity and Richness of Fungal Microorganism Communities
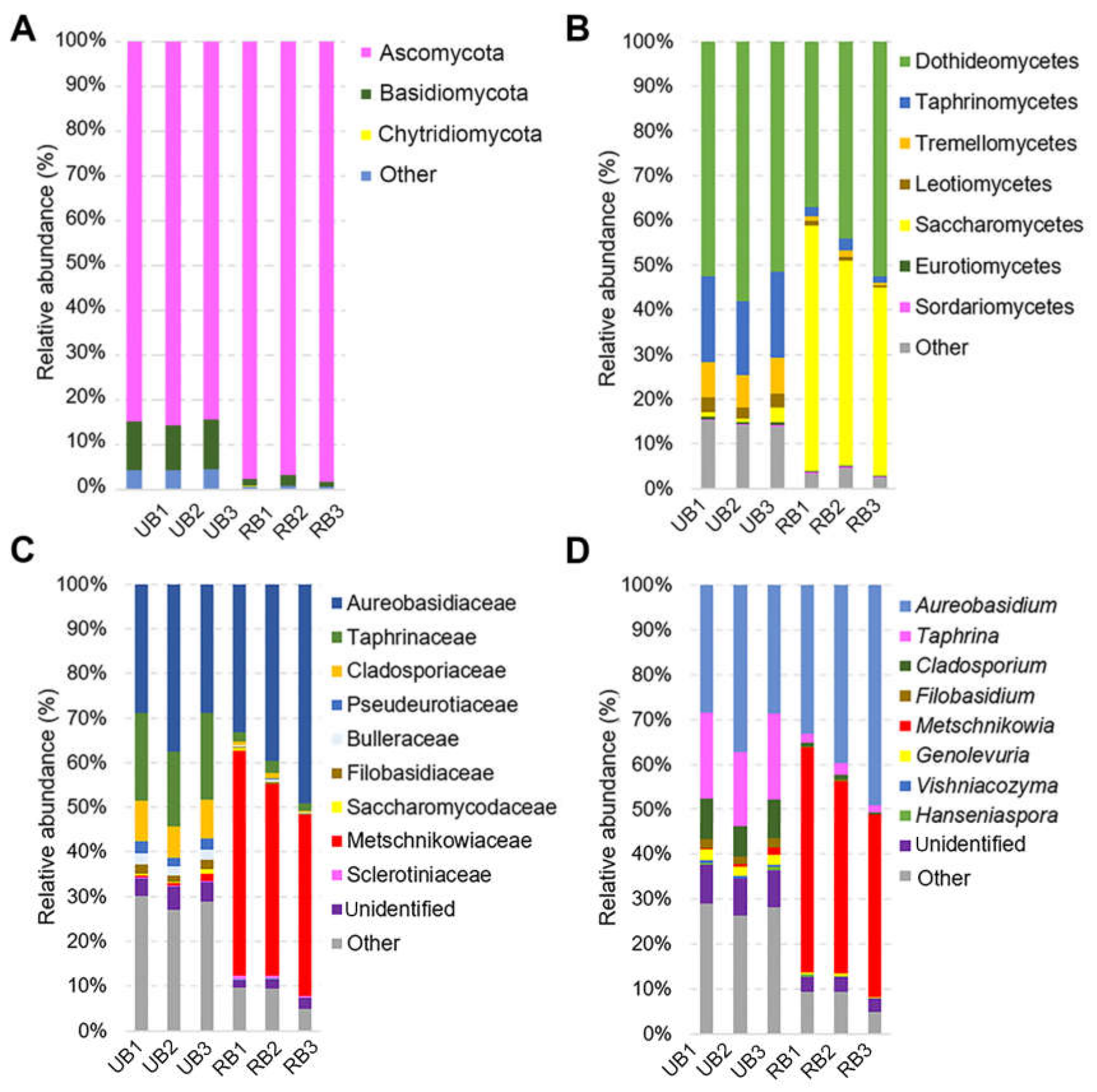
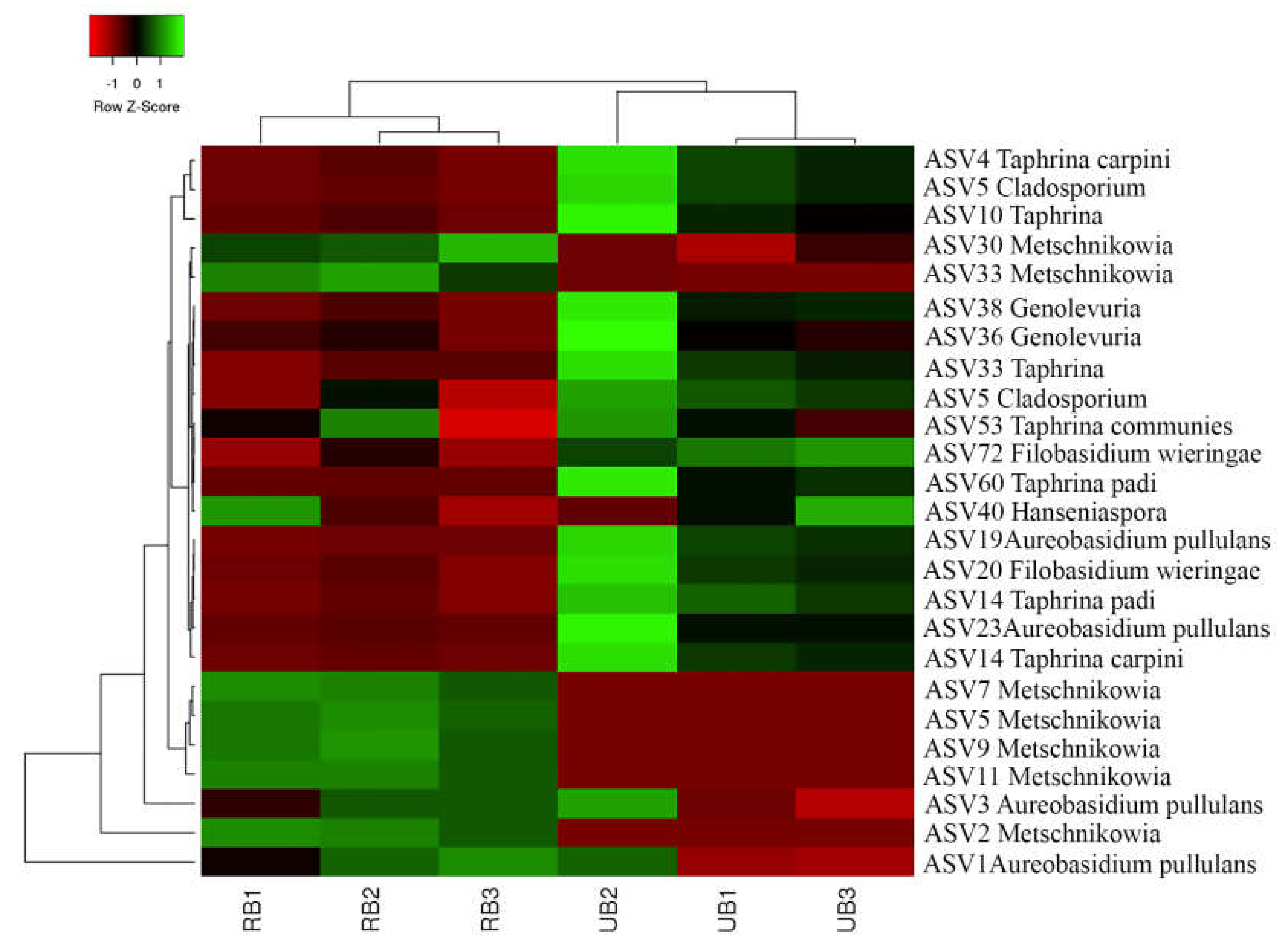
3.2. Composition of Fungal Microbiota on Unripe and Ripe Sea Buckthorn Berries
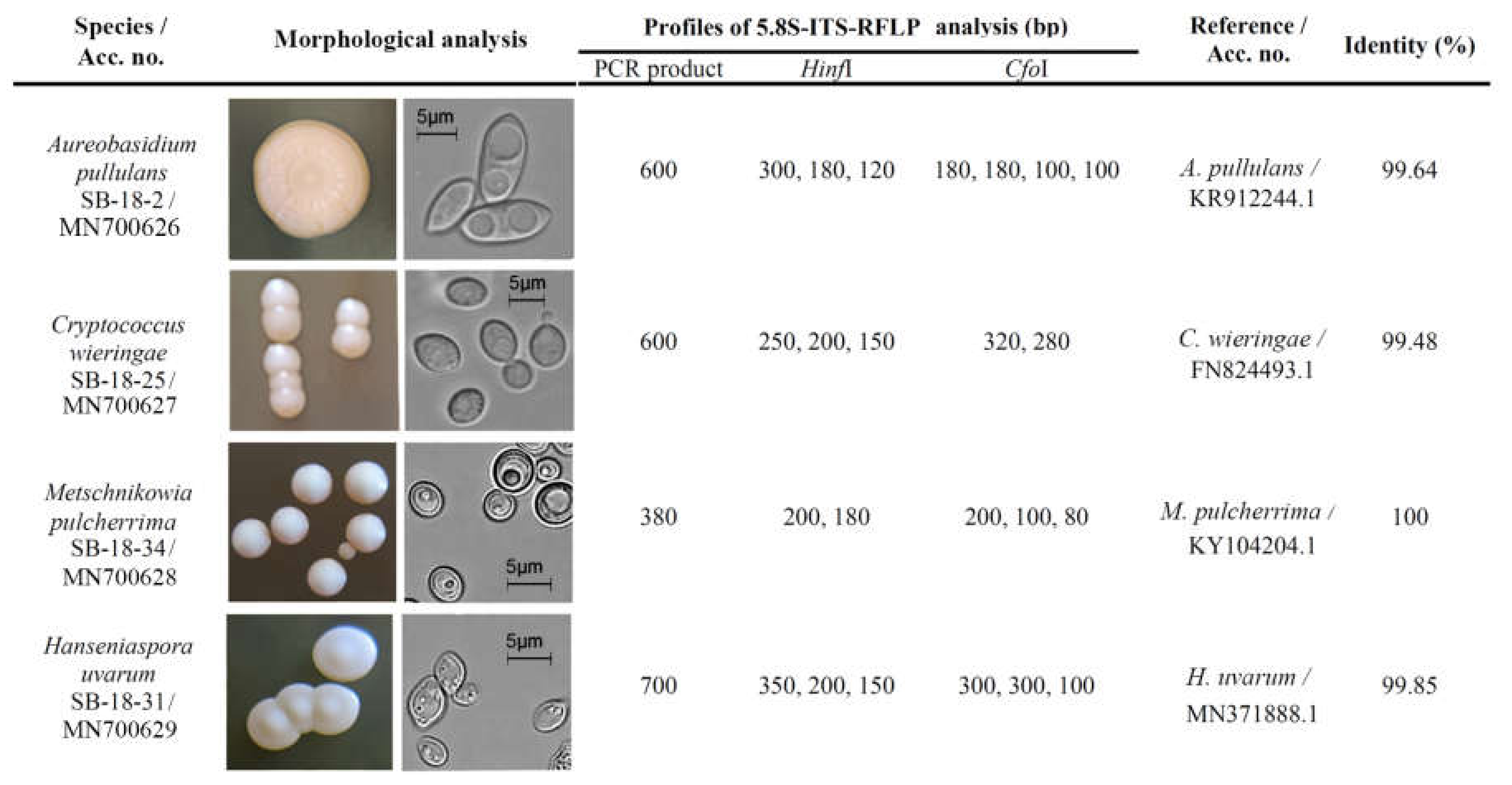
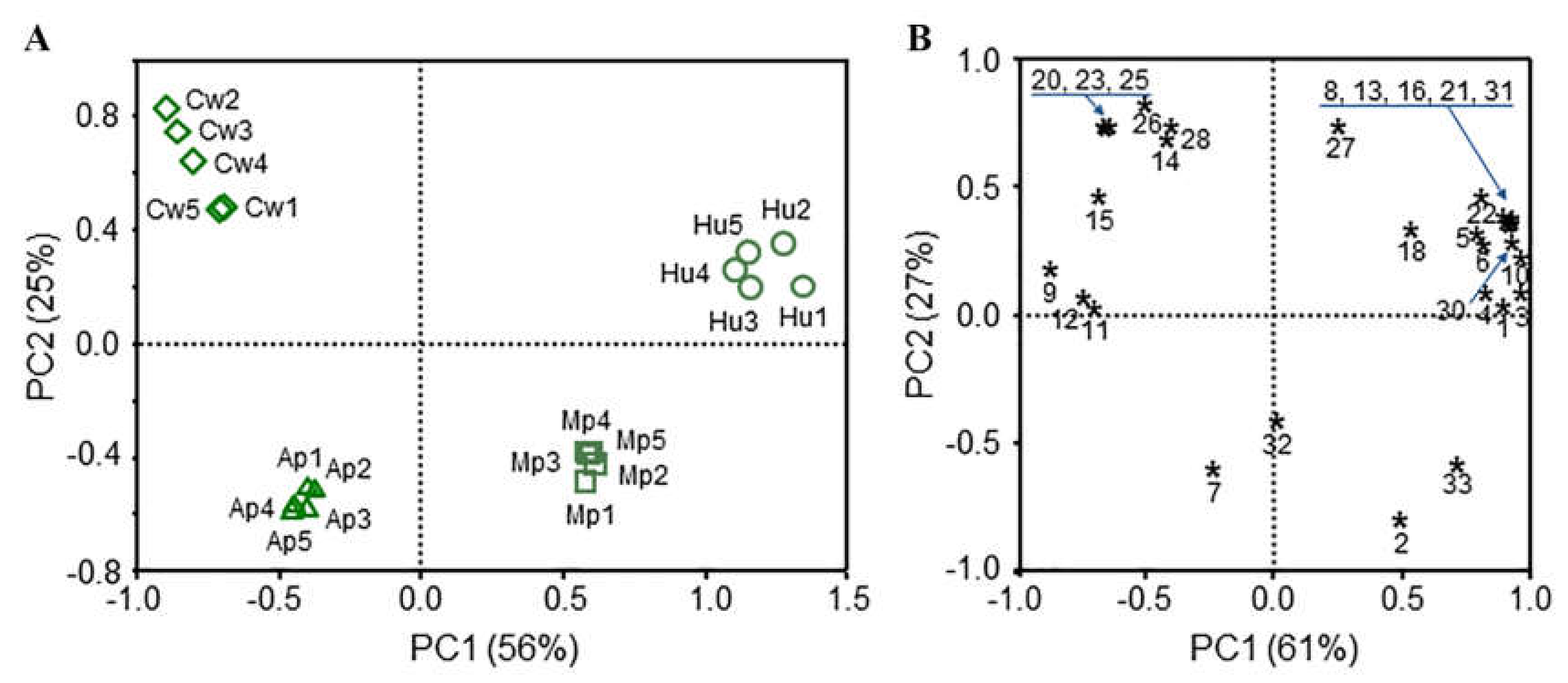
3.3. Sea-Buckthorn-Associated Cultivable Potential Biocontrol Yeasts and Their Produced Volatiles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ruan, C.J.; Rumpunen, K.; Nybom, H. Advances in improvement of quality and resistance in a multipurpose crop: sea buckthorn. Crit. Rev. Biotechnol. 2013, 33, 126–144. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Du, S.; Guo, K. Evaluation of limiting climatic factors and simulation of a climatically suitable habitat for Chinese sea buckthorn. PLoS ONE 2015, 10, e0131659. [Google Scholar]
- Li, T.S.C.; Schroeder, W.R. Sea buckthorn (Hippophae rhamnoides L.): a multipurpose plant. Hort. Technol. 1996, 6, 370–380. [Google Scholar] [CrossRef]
- Patel, C.A.; Divakar, K.; Santani, D.; Solanki, H.K.; Thakkar, J.H. Remedial Prospective of Hippophae rhamnoides Linn. (Sea Buckthorn). ISRN Pharmacol. 2012, 2012, 436857. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zheng, J.; Yang, B.; Trépanier, M.; Kallio, H. Effects of genotype, latitude, and weather conditions on the composition of sugars, sugar alcohols, fruit acids, and ascorbic acid in sea buckthorn (Hippophaë rhamnoides ssp. mongolica) berry juice. J. Agric. Food Chem. 2012, 60, 3180–3189. [Google Scholar] [CrossRef]
- Olas, B. The beneficial health aspects of sea buckthorn (Elaeagnus rhamnoides (L.) A. Nelson) oil. J. Ethnopharmacol. 2018, 213, 183–190. [Google Scholar] [CrossRef]
- Ulanowska, K.; Skalski, B.; Olas, B. Sea-buckthorn (Hippophae rhamnoides L.) as a source of compounds with antitumor and radioprotective activity. Postępy Higieny i Medycyny Doświadczalnej. 2018, 72, 240–252. [Google Scholar] [CrossRef]
- Martins, G.; Miot-Sertier, C.; Lauga, B.; Claisse, O.; Lonvaud-Funel, A.; Soulas, G.; Masneuf-Pomarède, I. Grape berry bacterial microbiota: Impact of the ripening process and the farming system. Int. J. Food Microbiol. 2012, 158, 93–100. [Google Scholar] [CrossRef]
- Taqarort, N.; Echairi, A.; Chaussod, R.; Nouaim, R.; Boubaker, H.; Benoumar, A.A.; Boudyach, E. Screening and identification of epiphytic yeasts with potential for biological control of green mold of citrus fruits. World J. Microbiol. Biotechnol. 2008, 24, 3031–3038. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Jurick, W.M.; Peter, K.A.; Kurtzman, C.P.; Buyer, J.S. Yeasts associated with plums and their potential for controlling brown rot after harvest. Yeast 2014, 31, 207–218. [Google Scholar] [CrossRef]
- Li, Z.T.; Janisiewicz, W.J.; Liu, Z.; Callahan, A.M.; Evans, B.E.; Jurick, W.M. II; Dardick. C. Exposure in vitro to an Environmentally Isolated Strain TC09 of Cladosporium sphaerospermum Triggers Plant Growth Promotion; Early Flowering; and Fruit Yield Increase. Front. Plant Sci. 2019, 9, 1959. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; Macarisin, D.; Wilson, C. Twenty years of postharvest biocontrol research: is it time for a new paradigm? Postharvest Biol. Technol. 2009, 52, 137–145. [Google Scholar] [CrossRef]
- Buser, C.C.; Newcomb, R.D.; Gaskett, A.C.; Goddard, M.R. Niche construction initiates the evolution of mutualistic interactions. Ecol. Lett. 2014, 17, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. Performance evaluation of volatile organic compounds by antagonistic yeasts immobilized on hydrogel spheres against gray; green and blue postharvest decays. Food Microbiol. 2017, 63, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Contarino, R.; Brighina, S.; Fallico, B.; Cirvilleri, G.; Parafati, L.; Restuccia, C. Volatile organic compounds (VOCs) produced by biocontrol yeasts. Food Microbiol. 2019, 82, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Freimoser, F.M.; Rueda-Mejia, M.P.; Tilocca, B.; Migheli, Q. Biocontrol yeasts: mechanisms and applications. World J. Microbiol. Biotech. 2019, 35, 154. [Google Scholar] [CrossRef] [PubMed]
- Becher, P.G.; Flick, G.; Rozpędowska, E.; Schmidt, A.; Hagman, A.; Lebreton, S.; Larsson, M.C.; Hansson, B.S.; Piškur, J.; Witzgall, P.; et al. Yeast, not fruit volatiles mediate Drosophila melanogaster attraction, oviposition and development. Funct. Ecol. 2012, 26, 822–828. [Google Scholar] [CrossRef]
- Davis, T.S.; Crippen, T.L.; Hofstetter, R.W.; Tomberlin, J.K. Microbial volatile emissions as insect semiochemicals. J. Chem. Ecol. 2013, 39, 840–859. [Google Scholar] [CrossRef]
- Pretorius, I.S. Tailoring wine yeast for the new millennium: novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Pinto, C.; Pinho, D.; Sousa, S.; Pinheiro, M.; Egas, C.; Gomes, A.C. Unravelling the diversity of grapevine microbiome. PLoS ONE 2014, 9, e85622. [Google Scholar] [CrossRef]
- Pinto, C.; Pinho, D.; Cardoso, R.; Custódio, V.; Fernandes, J.; Sousa, S.; Pinheiro, M.; Egas, C.; Gomes, A.C. Wine fermentation microbiome: A landscape from different Portuguese wine appellations. Front. Microbiol. 2015, 6, 905. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.; Moe, L.A. Multitrophic microbial interactions for eco- and agro-biotechnological processes: theory and practice. Trends Biotechnol. 2014, 32, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Abdelfattah, A.; Li Destri Nicosia, M.G.; Cacciola, S.O.; Droby, S.; Schena, L. Metabarcoding Analysis of Fungal Diversity in the Phyllosphere and Carposphere of Olive (Olea europaea). PLoS ONE 2015, 10, e0131069. [Google Scholar] [CrossRef] [PubMed]
- Abdelfattah, A.; Wisniewski, M.; Droby, S.; Schena, L. Spatial and compositional variation in the fungal communities of organic and conventionally grown apple fruit at the consumer point-of-purchase. Hortic Res. 2016, 3, 16047. [Google Scholar] [CrossRef] [PubMed]
- Abdelfattah, A.; Wisniewski, M.; Nicosia, M.G.L.D.; Cacciola, S.O.; Schena, L. Metagenomic analysis of fungal diversity on strawberry plants and the effect of management practices on the fungal community structure of aerial organs. PLoS ONE 2016, 11, e0160470. [Google Scholar] [CrossRef] [PubMed]
- Clooney, A.G.; Fouhy, F.; Sleator, R.D.; O’ Driscoll, A.; Stanton, C.; Cotter, P.D.; Claesson, M.J. Comparing apples and oranges?: Next Generation Sequencing and its impact on microbiome analysis. PLOS ONE 2016, 11, e0148028. [Google Scholar] [CrossRef]
- Droby, S.; Wisniewski, M.; Teixidó, N.; Spadaro, D.; Jijakli, M.H. The science, development, and commercialization of postharvest biocontrol products. Postharvest Biol. Tech. 2016, 122, 22–29. [Google Scholar] [CrossRef]
- Vepštaitė-Monstavičė, I.; Lukša, J.; Stanevičienė, R.; Strazdaitė-Žielienė, Ž.; Yurchenko, V.; Serva, S.; Servienė, E. Distribution of apple and blackcurrant microbiota in Lithuania and the Czech Republic. Microbiol. Res. 2018, 206, 1–8. [Google Scholar]
- Janisiewicz, W.J.; Kurtzman, C.P.; Buyer, J.S. Yeasts associated with nectarines and their potential for biological control of brown rot. Yeast 2010, 27, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Paramithiotis, S.; Kouretas, K.; Drosinos, E.H. Effect of ripening stage on the development of the microbial community during spontaneous fermentation of green tomatoes. J. Sci. Food Agric. 2014, 94, 1600–1606. [Google Scholar] [CrossRef] [PubMed]
- Diskin, S.; Feygenberg, O.; Maurer, D.; Droby, S.; Prusky, D.; Alkan, N. Microbiome alterations are correlated with occurrence of postharvest stem-end rot in mango fruit. Phytobiomes J. 2017, 1, 117–127. [Google Scholar] [CrossRef]
- Vadkertiová, R.; Molnárová, J.; Vránová, D.; Sláviková, E. Yeasts and yeast-like organisms associated with fruits and blossoms of different fruit trees. Can. J. Microbiol. 2012, 58, 1344–1352. [Google Scholar] [CrossRef] [PubMed]
- Lukša, J.; Vepštaitė-Monstavičė, I.; Yurchenko, V.; Serva, S.; Servienė, E. High content analysis of sea buckthorn, black chokeberry, red and white currants microbiota – A pilot study. Food Res. Int. 2018, 111, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yang, D.; Yu, X. Soil microbial communities and enzyme activities in sea-buckthorn (Hippophae rhamnoides) plantation at different ages. PLoS ONE 2018, 13, e0190959. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Sagar, A. Microbial associates of Hippophae rhamnoides (Seabuckthorn). Plant Pathol. J. 2007, 6, 299–305. [Google Scholar]
- Toju, H.; Tanabe, A.S.; Yamamoto, S.; Sato, H. High-coverage ITS primers for the DNA-based identification of ascomycetes and basidiomycetes in environmental samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. QIIME 2: Reproducible, interactive, scalable, and extensible microbiome data science. PeerJ Preprints 2018, 6, e27295v2. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Nilsson, R.H.; Larsson, K.H.; Taylor, A.F.S.; Bengtsson-Palme, J.; Jeppesen, T.S.; Schigel, D.; Kennedy, P.; Picard, K.; Glöckner, F.O.; Tedersoo, L.; et al. The UNITE database for molecular identification of fungi: handling dark taxa and parallel taxonomic classifications. Nucleic Acids Res. 2018, 47, D259–D264. [Google Scholar] [CrossRef]
- Tilocca, B.; Cao, A.; Migheli, Q. Scent of a Killer: Microbial Volatilome and Its Role in the Biological Control of Plant Pathogens. Front. Microbiol. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of yeasts by RFLP analysis of the 5.8S rRNA gene and the two ribosomal internal transcribed spacers. Int. J. Syst. Bacteriol. 1999, 49, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Barata, A.; Malfeito-Ferreira, M.; Loureiro, V. The microbial ecology of wine grape berries. Int. J. Food Microbiol. 2012, 153, 243–259. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, A.; Ugolini, L.; Lazzeri, L.; Mari, M. Production of volatile organic compounds by Aureobasidium pullulans as apotential mechanism of action against postharvest fruit pathogens. Biol. Control. 2015, 81, 8–14. [Google Scholar] [CrossRef]
- Wisniewski, M.; Wilson, C.; Droby, S.; Chalutz, E.; El Ghaouth, A.; Stevens, C. Biological Control: A Global Perspective; CABI: Cambridge, MA, USA, 2007; pp. 262–273. [Google Scholar]
- Piombo, E.; Sela, N.; Wisniewski, M.; Hoffmann, M.; Gullino, M.L.; Allard, M.W.; Levin, E.; Spadaro, D.; Droby, S. Genome sequence, assembly and characterization of two Metschnikowia fructicola strains used as biocontrol agents of postharvest diseases. Front. Microbiol. 2018, 9, 593. [Google Scholar] [CrossRef]
- Parafati, L.; Vitale, A.; Restuccia, C.; Cirvilleri, G. Biocontrol ability and action mechanism of food-isolated yeast strains against Botrytis cinerea causing post-harvest bunch rot of table grape. Food Microbiol. 2015, 47, 85–92. [Google Scholar] [CrossRef]
- Masoud, W.; Poll, L.; Jakobsen, M. Influence of volatile compounds produced by yeasts predominant during processing of Coffea arabica in East Africa on growth and ochratoxin A (OTA) production by Aspergillus ochraceus. Yeast 2005, 22, 1133–1142. [Google Scholar] [CrossRef]
- Elad, Y.; Köhl, J.; Fokkema, N. Control of infection and sporulation of Botrytis cinerea on bean and tomato by saprophytic yeasts. Phytopathol. 1994, 84, 1193–1200. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Zheng, X.D.; Yu, T. Biological control of postharvest diseases of peach with Cryptococcus laurentii. Food Control. 2007, 18, 287–291. [Google Scholar] [CrossRef]
- Hashem, M.; Alamri, S.A.; Hesham, A.E.L.; Al-Qahtani, F.M.H.; Kilany, M. Biocontrol of apple blue mould by new yeast strains: Cryptococcus albidus KKUY0017 and Wickerhamomyces anomalus KKUY0051 and their mode of action. Biocontrol. Sci. Technol. 2014, 24, 1137–1152. [Google Scholar] [CrossRef]
- Lederer, M.A.; Nielsen, D.S.; Toldam-Andersen, T.B.; Herrmann, J.V.; Arneborg, N. Yeast species associated with different wine grape varieties in Denmark. Acta Agr. Scand. B-S. P. 2013, 63, 89–96. [Google Scholar] [CrossRef]
- Grube, M.; Schmid, F.; Berg, G. Black fungi and associated bacterial communities in the phyllosphere of grapevine. Fungal Biol. 2011, 115, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Parsa, S.; García-Lemos, A.M.; Castillo, K.; Ortiz, V.; López-Lavalle, L.A.B.; Braun, J.; Vega, F.E. Fungal endophytes in germinated seeds of the common bean, Phaseolus vulgaris. Fungal Biol. 2016, 120, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Tsai, I.J.; Tanaka, E.; Masuya, H.; Tanaka, R.; Hirooka, Y.; Endoh, R.; Sahashi, N.; Kikuchi, T. Comparative genomics of taphrina fungi causing varying degrees of tumorous deformity in plants. Genome Biol. Evol. 2014, 6, 861–872. [Google Scholar] [CrossRef]
- Wang, X.; Radwan, M.M.; Taráwneh, A.H.; Gao, J.; Wedge, D.E.; Rosa, L.H.; Cutler, H.G.; Cutler, S.J. Antifungal activity against plant pathogens of metabolites from the endophytic fungus cladosporium cladosporioides. J. Agric. Food Chem. 2013, 61, 4551–4555. [Google Scholar] [CrossRef] [PubMed]
- Wit, P.J.G.M.; van der Burgt, A.; Ökmen, B.; Stergiopoulos, I.; Abd-Elsalam, K.A.; Aerts, A.L.; Bahkali, A.H.; Beenen, H.G.; Chettri, P.; Cox, M.P.; et al. The genomes of the fungal plant pathogens Cladosporium fulvum and Dothistroma septosporum reveal adaptation to different hosts and lifestyles but also signatures of common ancestry. PLoS Genetics 2012, 8, e1005775. [Google Scholar]
- Sandoval-Denis, M.; Gené, J.; Sutton, D.A.; Wiederhold, N.P.; Cano-Lira, J.F.; Guarro, J. New species of Cladosporium associated with human and animal infections. Persoonia. 2016, 36, 281–298. [Google Scholar] [CrossRef]
- Bernal-Martinez, L.; Gomez-Lopez, A.; Castelli, M.V.; Mesa-Arango, A.C.; Zaragoza, O.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M. Susceptibility profile of clinical isolates of non-Cryptococcus neoformans/non-Cryptococcus gattii Cryptococcus species and literature review. Med. Mycol. 2010, 48, 90–96. [Google Scholar] [CrossRef]
- Santo, D.E.; Galego, L.; Gonçalves, T.; Quintas, C. Yeast diversity in the Mediterranean strawberry tree (Arbutus unedo L.) fruits’ fermentations. Food Res. Int. 2012, 47, 45–50. [Google Scholar] [CrossRef]
- Graça, A.; Santo, D.; Esteves, E.; Nunes, C.; Abadias, M.; Quintas, C. Evaluation of microbial quality and yeast diversity in fresh-cut apple. Food Microbiol. 2015, 51, 179–185. [Google Scholar] [CrossRef]
- Liu, H.M.; Guo, J.H.; Cheng, Y.J.; Luo, L.; Liu, P.; Wang, B.Q.; Deng, B.X.; Long, C. Control of gray mold of grape by Hanseniaspora uvarum and its effects on postharvest quality parameters. Ann. Microbiol. 2010, 60, 31–35. [Google Scholar] [CrossRef]
- Janisiewicz, W.J.; Tworkoski, T.J.; Kurtzman, C.P. Biocontrol potential of Metschnikowia pulcherrima strains against blue mold of apple. Phytopathol. 2001, 91, 1098–1108. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, D.; Ciavorella, A.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Metschnikowia pulcherrima strain MACH1 outcompetes Botrytis cinerea, Alternaria alternata and Penicillium expansum in apples through iron depletion. Postharvest Biol. Technol. 2008, 49, 121–128. [Google Scholar] [CrossRef]
- Suh, S.O.; McHugh, J.V.; Pollock, D.D.; Blackwell, M. The beetle gut: a hyperdiverse source of novel yeasts. Mycol. Res. 2005, 109, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Pozo, M.I.; Lachance, M.A.; Herrera, C.M. Nectar yeasts of two southern Spanish plants: The roles of immigration and physiological traits in community assembly. FEMS Microbiol. Ecol. 2012, 80, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Ebert, B.E.; Halbfeld, C.; Blank, L.M. Exploration and exploitation of the yeast volatilome. Curr. Metabolomics 2017, 5, 102–118. [Google Scholar] [CrossRef]
- Mozūraitis, R.; Aleknavičius, D.; Vepštaitė-Monstavičė, I.; Stanevičienė, R.; Emamic, S.N.; Apšegaitė, V.; Radžiutė, S.; Blažytė-Čereškienė, L.; Servienė, E.; Būda, V. Hippophae rhamnoides berry related Pichia kudriavzevii yeast volatiles modify behaviour of Rhagoletis batava flies. J. Adv. Res. 2020, 21, 71–77. [Google Scholar] [CrossRef]
- Bakht, J.; Noor, N.; Iqbal, A.; Shafi, M. Antimicrobial activity of different solvent extracted samples from the leaves and fruits of Capsicum annuum. Pak. J. Pharm. Sci. 2020, 33, 27–32. [Google Scholar]
- Huang, R.; Li, G.Q.; Zhang, J.; Yang, L.; Che, H.J.; Jiang, D.H.; Huang, H.C. Control of post-harvest botrytis fruit rot of strawberry by volatile organic compounds of Candida intermedia. Phytopathol. 2011, 101, 859–869. [Google Scholar] [CrossRef]
- Piper, A.M.; Farnier, K.; Linder, T.; Speight, R.; Cunningham, J.P. Two gut-associated yeasts in a tephritid fruit fly have contrasting effects on adult attraction and larval survival. J. Chem. Ecol. 2017, 43, 891–901. [Google Scholar] [CrossRef]
- Babcock, T.; Borden, J.H.; Gries, R.; Carroll, C.; Lafontaine, J.P.; Moore, M.; Gries, G. Inter-kingdom signaling - symbiotic yeasts produce semiochemicals that attract their yellowjacket hosts. Entomol. Exp. Appl. 2019, 167, 220–230. [Google Scholar] [CrossRef]
- Becher, P.G.; Hagman, A.; Verschut, V.; Chakraborty, A.; Rozpedowska, E.; Lebreton, S.; Bengtsson, M.; Flick, G.; Witzgall, P.; Piškur, J.; et al. Chemical signaling and insect attraction is a conserved trait in yeasts. Ecol. Evol. 2018, 8, 2962–2974. [Google Scholar] [CrossRef] [PubMed]
- Versace, E.; Eriksson, A.; Rocchi, F.; Castellan, I.; Sgado, P.; Haase, A. Physiological and behavioral responses in Drosophila melanogaster to odorants present at different plant maturation stages. Physiol. Behav. 2016, 163, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Dobzhansky, T.; Cooper, D.M.; Phaff, H.J.; Knapp, E.P.; Carson, H.L. Studies on the ecology of Drosophila in the Yosemite region of California differential attraction of species of Drosophila to different species of yeasts. Ecol. 1956, 37, 544–550. [Google Scholar] [CrossRef]
- Ganter, P.F. Yeast and invertebrate associations. In Biodiversity and Ecophysiology of Yeasts; Augusto, R.C., Gábor, P., Eds.; Springer: Berlin, Germany, 2006; pp. 303–370. [Google Scholar]
- Scheidler, N.H.; Liu, C.; Hamby, K.A.; Zalom, F.G.; Syed, Z. Volatile codes: Correlation of olfactory signals and reception in Drosophila-yeast chemical communication. Sci. Rep. 2015, 5, 14059. [Google Scholar] [CrossRef] [PubMed]
- Holighaus, G.; Rohlfs, M. Fungal allelochemicals in insect pest management. Appl. Microbiol. Biotechnol. 2016, 100, 5681–5689. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reads Obtained | High Quality Reads | ASV | Shanon Diversity | Pielou Eveness | Simpson Index | |
|---|---|---|---|---|---|---|
| UB1 | 103,024 | 49,891 | 300 | 5.26 | 0.64 | 0.94 |
| UB2 | 164,922 | 110,782 | 419 | 5.00 | 0.58 | 0.91 |
| UB3 | 89,720 | 43,458 | 295 | 5.37 | 0.66 | 0.94 |
| RB1 | 114,814 | 76,641 | 190 | 3.71 | 0.49 | 0.86 |
| RB2 | 135,098 | 92,822 | 240 | 3.795 | 0.48 | 0.85 |
| RB3 | 119,056 | 80,944 | 171 | 3.21 | 0.44 | 0.79 |
| Total | 726,634 | 454,538 | 1615 |
| No. | Compound | CAS No. | RI | GR | H. uvarum | M. pulcherrima | A. pullulans | C. wieringae | Control |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Ethyl acetate | 141-78-6 | 898 | ES | 26.26 ± 4.63 | 8.98 ± 0.28 | 0 | 0 | 0.06 ± 0.02 |
| 2 | Ethanol | 64-17-5 | 902 | OH | 4.58 ± 0.36 | 4.89 ± 0.42 | 6.26 ± 0.82 | 0.39 ± 0.03 | 0.10 ± 0.02 |
| 3 | Ethyl propionate | 1105-37-3 | 915 | ES | 1.00 ± 0.05 | 0.24 ± 0.01 | 0 | 0 | 0.01 ± 0.001 |
| 4 | 2-Methylprop-1-yl acetate | 110-19-0 | 985 | ES | 0.31 ± 0.09 | 0.07 ± 0.01 | 0 | 0 | 0 |
| 5 | Ethyl butanoate | 105-54-4 | 1013 | ES | 0.05 ± 0.03 | 0 | 0 | 0 | 0 |
| 6 | Ethyl 2-methylbutanoate | 7452-79-1 | 1033 | ES | 0.02 ± 0.01 | 0 | 0 | 0 | 0 |
| 7 | Ethyl 3-methylbutanoate | 108-64-5 | 1050 | ES | 0 | 0 | 0.04 ± 0.01 | 0 | 0 |
| 8 | Butyl acetate | 123-86-4 | 1063 | ES | 0.28 ± 0.03 | 0 | 0 | 0 | 0.10 ± 0.02 |
| 9 | 2-Methylpropanol | 78-83-1 | 1095 | OH | 0.42 ± 0.07 | 0.98 ± 0.08 | 1.04 ± 0.13 | 1.70 ± 0.11 | 0 |
| 10 | 3-Methylbutyl acetate | 123-92-2 | 1105 | ES | 13.28 ± 0.5 | 0.40 ± 0.02 | 0.15 ± 0.05ns | 0.07 ± 0.01ns | 0.10 ± 0.07 |
| 11 | Butanol | 71-36-3 | 1149 | OH | 0 | 0 | 0.13 ± 0.03 | 0.09 ± 0.01 | 0 |
| 12 | 2-Heptanone | 110-43-0 | 1170 | KT | 0 | 0 | 0.07 ± 0.01 | 0.06 ± 0.01 | 0 |
| 13 | 3-Methylbutyl propionate | 105-68-0 | 1176 | ES | 0.62 ± 0.09 | 0 | 0 | 0 | 0 |
| 14 | 2-Methylbutanol | 137-32-6 | 1207 | OH | 1.63 ± 0.28 | 1.77 ± 0.24 | 0.84 ± 0.18 | 6.47 ± 1.21 | 0 |
| 15 | 3-Methylbutanol | 123-51-3 | 1213 | OH | 3.28 ± 0.37 | 5.95 ± 0.56 | 4.07 ± 0.51 | 10.39 ± 0.36 | 0.21 ± 0.11 |
| 16 | Ethyl hexanoate | 123-66-0 | 1224 | ES | 0.02 ± 0.01 | 0 | 0 | 0 | 0 |
| 17 | Styrene* | 100-42-5 | 1238 | AR | 0.20 ± 0.04ns | 0.13 ± 0.03ns | 0.07 ± 0.01ns | 0.16 ± 0.01ns | 0.14 ± 0.06ns |
| 18 | 2-hydroxy-3-butanone | 513-86-0 | 1273 | KT | 0.15±0.04 | 0.14 ± 0.01 | 0 | 0.06 ± 0.01 | 0 |
| 19 | 2,5- Dimethyl pyrazine* | 123-32-0 | 1316 | PY | 0.02±0.01ns | 0.01 ± 0.001ns | 0.03 ± 0.01ns | 0.04 ± 0.01ns | 0.03 ± 0.02ns |
| 20 | 6-Methyl 5-hepten-2-one | 110-93-0 | 1327 | KT | 0 | 0 | 0 | 0.93 ± 0.10 | 0 |
| 21 | Ethyl octanoate | 106-32-1 | 1430 | ES | 0.12 ± 0.01 | 0 | 0 | 0 | 0 |
| 22 | Acetic acid | 64-19-7 | 1449 | FA | 1.15 ±0.25 | 0 | 0.01 ± 0.008 | 0.07 ± 0.02 | 0 |
| 23 | 6-Methyl-5-hepten-2-ol | 1569-60-4 | 1461 | OH | 0 | 0 | 0 | 0.08 ± 0.02 | 0 |
| 24 | 2-Ethylhexanol* | 104-76-7 | 1488 | OH | 0.02 ± 0.001ns | 0.02±0.01ns | 0.02±0.001ns | 0.02±0.004ns | 0.03±0.01ns |
| 25 | Propionic acid | 1979-09-04 | 1528 | FA | 0 | 0 | 0 | 0.03 ± 0.004 | 0 |
| 26 | 2-Methylpropionic acid | 79-31-2 | 1562 | FA | 0 | 0 | 0 | 0.18 ± 0.04 | 0 |
| 27 | Butanoic acid | 107-92-6 | 1634 | FA | 0.03 ± 0.01 | 0 | 0 | 0.03 ± 0.01 | 0 |
| 28 | 3-Methylbutanoic acid | 503-74-2 | 1703 | FA | 0.07 ± 0.02 | 0.01 ± 0.001 | 0.06 ± 0.04 | 0.71 ± 0.25 | 0 |
| 29 | Methoxy-phenyl-oxime* | 1767 | IM | 0.03 ± 0.01ns | 0.01 ± 0.003 | 0.01 ± 0.004 | 0.03 ± 0.01ns | 0.03 ± 0.01ns | |
| 30 | 2-Phenylethyl acetate | 103-45-7 | 1795 | ES/AR | 3.52 ± 0.64 | 0.04 ± 0.01 | 0 | 0 | 0 |
| 31 | 2-Phenylethyl propionate | 122-70-3 | 1858 | ES/AR | 0.11 ± 0.02 | 0 | 0 | 0 | 0 |
| 32 | Phenylmethanol | 1960-12-08 | 1856 | AR | 0 | 0.03 ± 0.01 | 0 | 0 | 0 |
| 33 | 2-Phenylethanol | 1960-12-08 | 1894 | AR | 3.60 ± 0.26 | 3.45 ± 0.65 | 2.28 ± 0.45 | 0.21 ± 0.03 | 0.03 ± 0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lukša, J.; Vepštaitė-Monstavičė, I.; Apšegaitė, V.; Blažytė-Čereškienė, L.; Stanevičienė, R.; Strazdaitė-Žielienė, Ž.; Ravoitytė, B.; Aleknavičius, D.; Būda, V.; Mozūraitis, R.; et al. Fungal Microbiota of Sea Buckthorn Berries at Two Ripening Stages and Volatile Profiling of Potential Biocontrol Yeasts. Microorganisms 2020, 8, 456. https://doi.org/10.3390/microorganisms8030456
Lukša J, Vepštaitė-Monstavičė I, Apšegaitė V, Blažytė-Čereškienė L, Stanevičienė R, Strazdaitė-Žielienė Ž, Ravoitytė B, Aleknavičius D, Būda V, Mozūraitis R, et al. Fungal Microbiota of Sea Buckthorn Berries at Two Ripening Stages and Volatile Profiling of Potential Biocontrol Yeasts. Microorganisms. 2020; 8(3):456. https://doi.org/10.3390/microorganisms8030456
Chicago/Turabian StyleLukša, Juliana, Iglė Vepštaitė-Monstavičė, Violeta Apšegaitė, Laima Blažytė-Čereškienė, Ramunė Stanevičienė, Živilė Strazdaitė-Žielienė, Bazilė Ravoitytė, Dominykas Aleknavičius, Vincas Būda, Raimondas Mozūraitis, and et al. 2020. "Fungal Microbiota of Sea Buckthorn Berries at Two Ripening Stages and Volatile Profiling of Potential Biocontrol Yeasts" Microorganisms 8, no. 3: 456. https://doi.org/10.3390/microorganisms8030456
APA StyleLukša, J., Vepštaitė-Monstavičė, I., Apšegaitė, V., Blažytė-Čereškienė, L., Stanevičienė, R., Strazdaitė-Žielienė, Ž., Ravoitytė, B., Aleknavičius, D., Būda, V., Mozūraitis, R., & Servienė, E. (2020). Fungal Microbiota of Sea Buckthorn Berries at Two Ripening Stages and Volatile Profiling of Potential Biocontrol Yeasts. Microorganisms, 8(3), 456. https://doi.org/10.3390/microorganisms8030456