Removal of Soil Microbes Alters Interspecific Competitiveness of Epichloë Endophyte-Infected over Endophyte-Free Leymus chinensis

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant and Fungal Material
2.2. Experimental Design
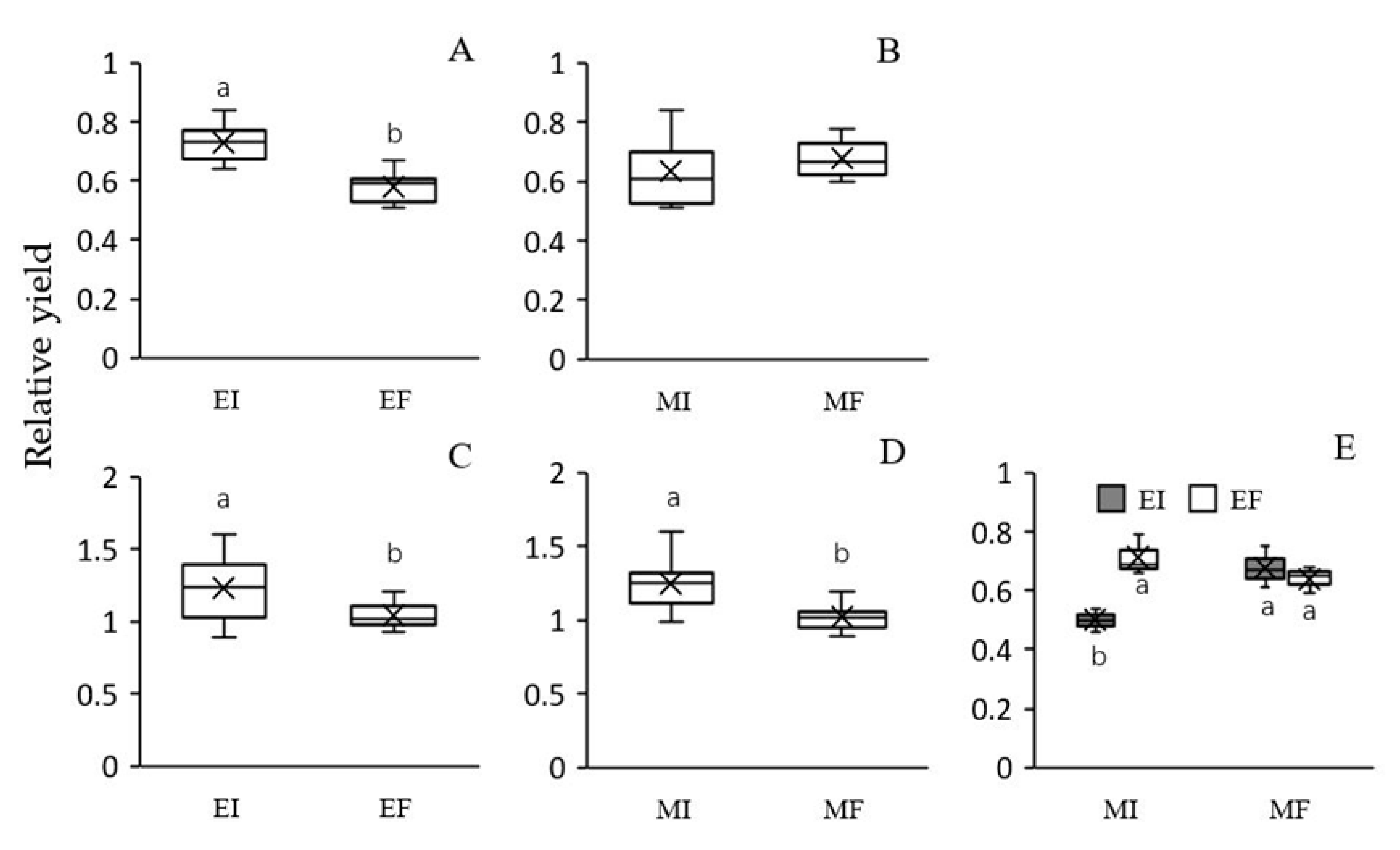
2.3. Biomass and Relative Yield (RY)
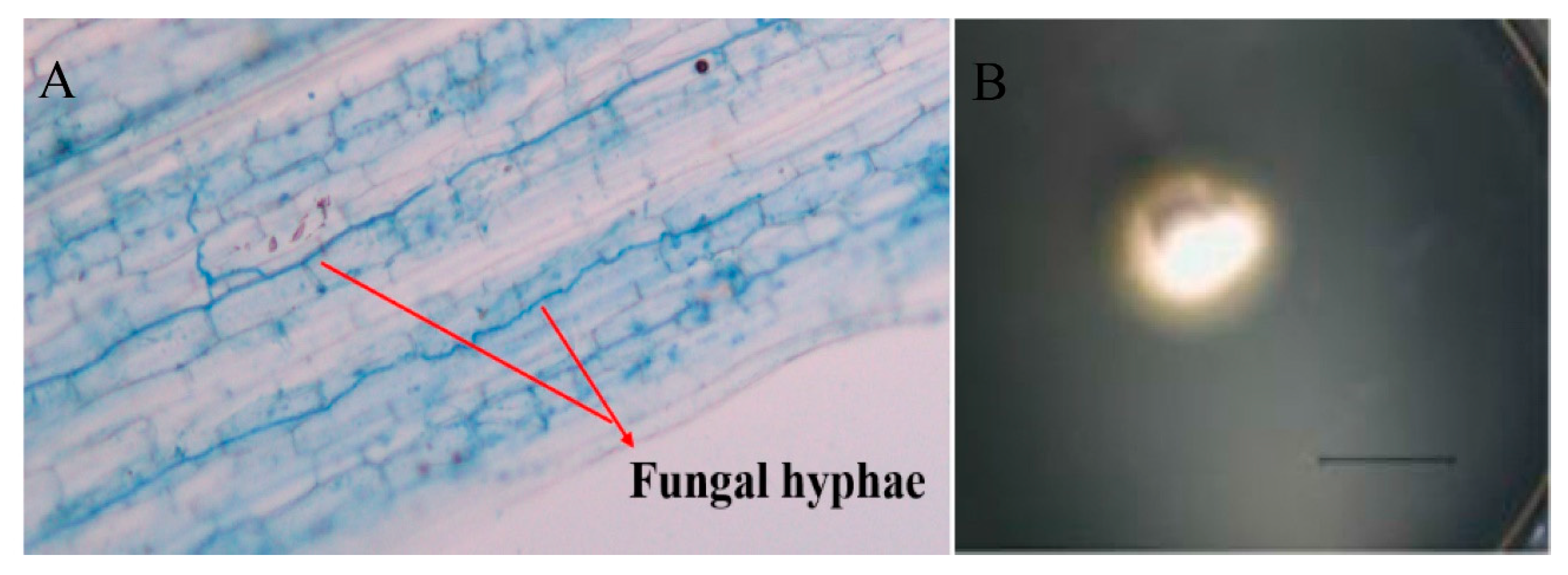
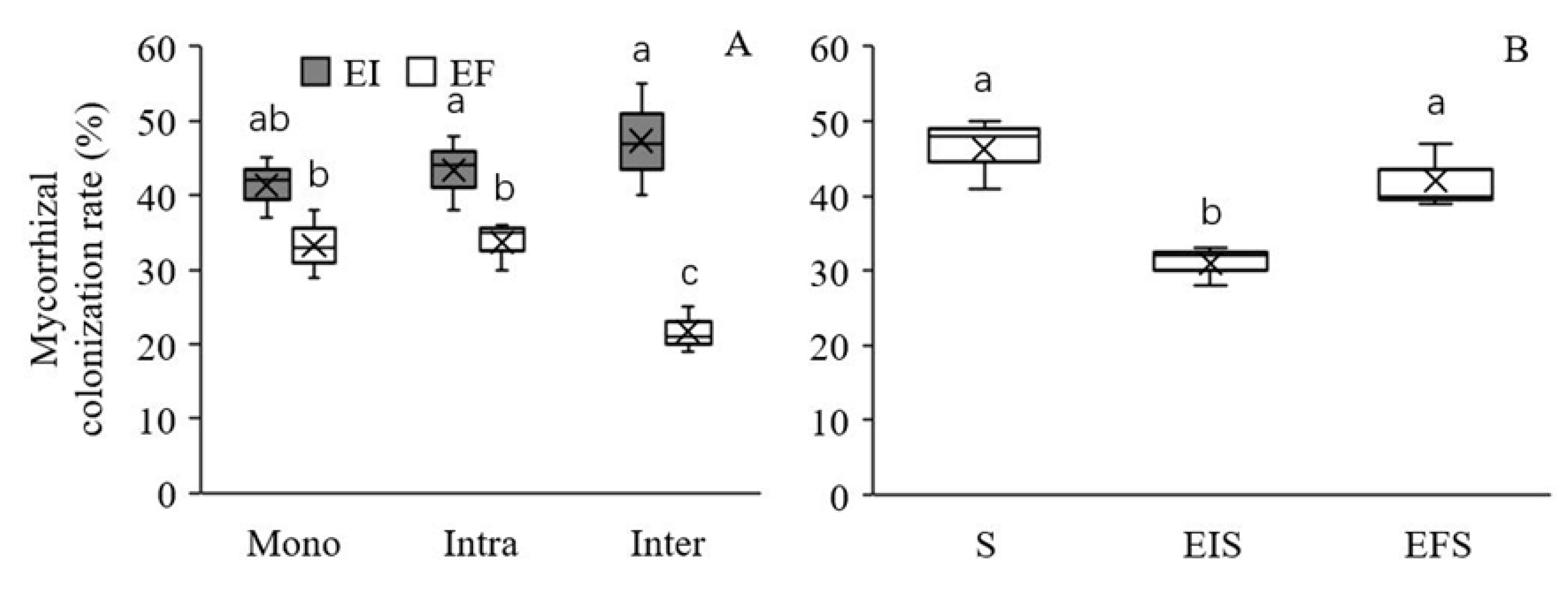
2.4. AMF Colonization Rate
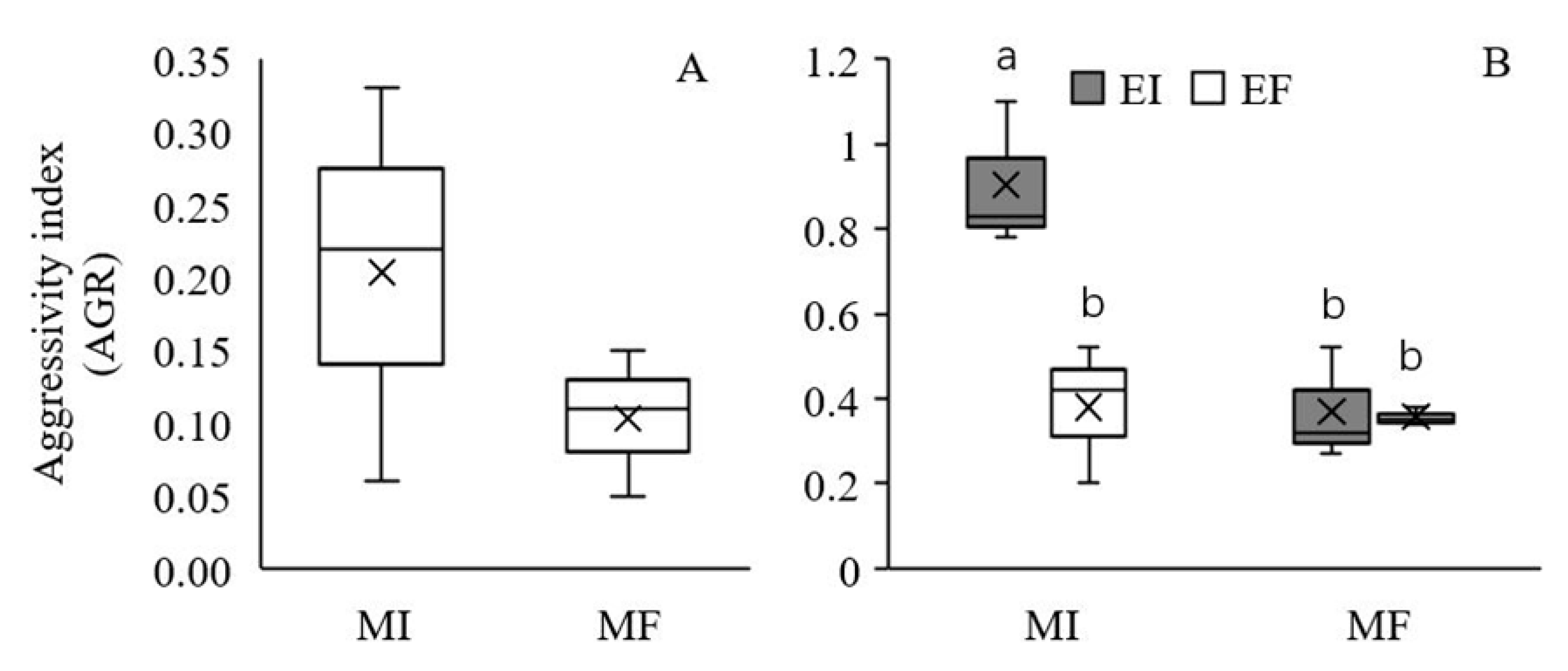
2.5. Competitiveness
2.6. Statistical Analyses
3. Results
3.1. Plant Growth Performance
3.2. AMF Colonization Rate
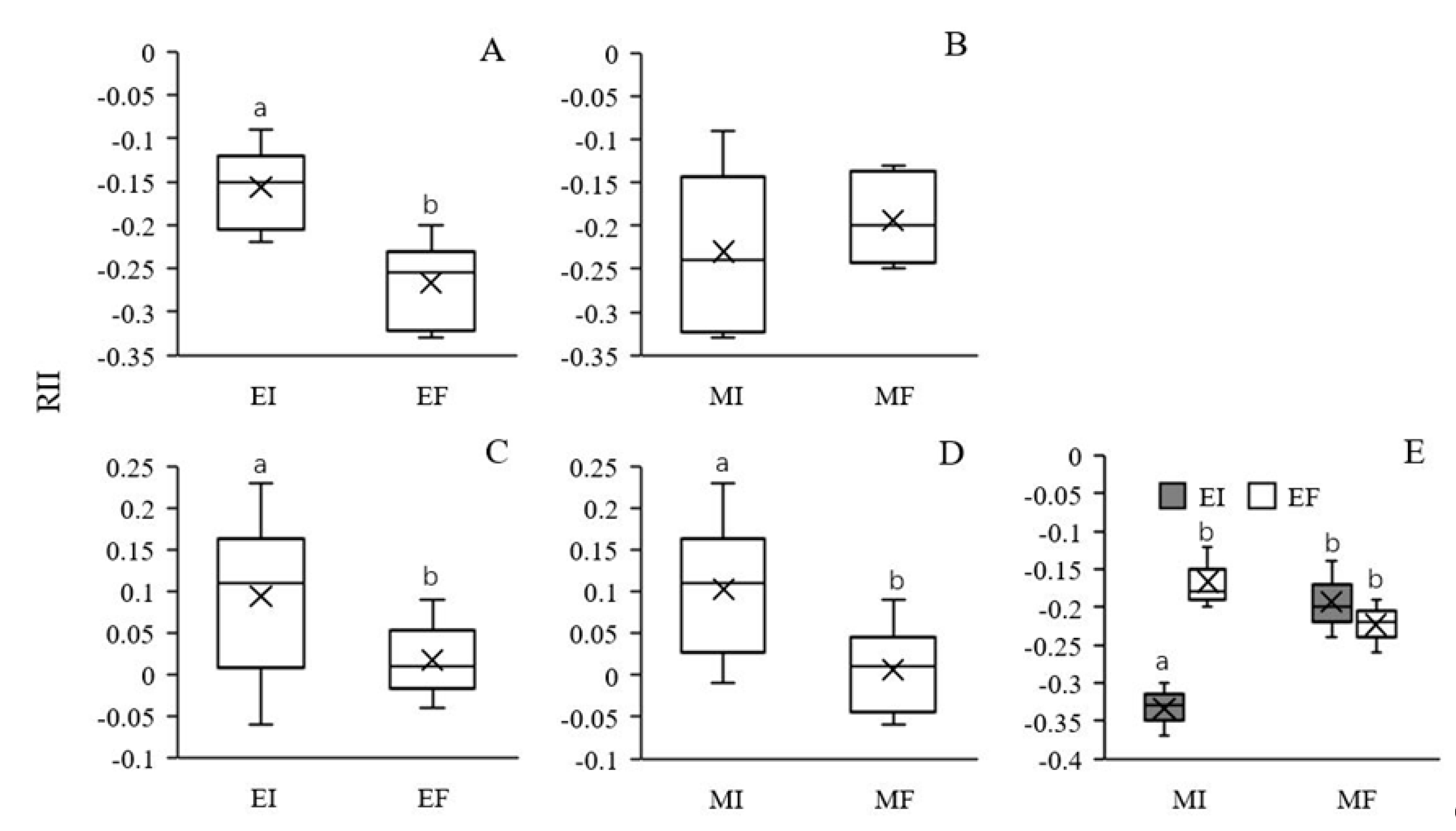
3.3. Competitiveness
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Arnold, A.E.; Maynard, Z.; Gilbert, G.S.; Coley, P.D.; Kursar, T.A. Are tropical fungal endophytes hyperdiverse? Ecol. Lett. 2000, 3, 267–274. [Google Scholar] [CrossRef]
- Clay, K. Fungal endophytes of grasses. Annu. Rev. Ecol. Syst. 1990, 21, 275–297. [Google Scholar] [CrossRef]
- Clay, K.; Schardl, C. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. Am. Nat. 2002, 160, S99–S127. [Google Scholar] [CrossRef] [PubMed]
- Saikkonen, K.; Wali, P.; Helander, M.; Faeth, S.H. Evolution of endophyte-plant symbioses. Trends Plant Sci. 2004, 9, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Bacon, C.W.; White, J.F. Physiology and Drought Tolerance of Endophyte-Infected Grasses; CRC Press: Boca Raton, FL, USA, 1994. [Google Scholar]
- Kannadan, S.; Rudgers, J.A. Endophyte symbiosis benefits a rare grass under low water availability. Funct. Ecol. 2008, 22, 706–713. [Google Scholar] [CrossRef]
- Liu, H.; Chen, W.; Wu, M.; Wu, R.H.; Zhou, Y.; Gao, Y.B.; Ren, A.Z. Arbuscular mycorrhizal fungusinoculation reduces the drought-resistance advantage of endophyte-infected versus endophyte-free Leymus chinensis. Mycorrhiza 2017, 27, 791–799. [Google Scholar] [CrossRef]
- Malinowski, D.P.; Belesky, D.P. Neotyphodium coenophialum endophyte infection affects the ability of tall fescue to use sparingly available phosphorus. J. Plant Nutr. 1999, 22, 835–853. [Google Scholar] [CrossRef]
- Malinowski, D.P.; Alloush, G.A.; Belesky, D.P. Leaf endophyte Neotyphodium coenophialum modifies mineral uptake in tall fescue. Plant Soil 2000, 227, 115–126. [Google Scholar] [CrossRef]
- Rahman, M.H.; Saiga, S. Endophytic fungi (Neotyphodium coenophialum) affect the growth and mineral uptake, transport and efficiency ratios in tall fescue (Festuca arundinacea). Plant Soil 2005, 272, 163–171. [Google Scholar] [CrossRef]
- Clay, K. Fungal endophytes of grasses: A defensive mutualism between plants and fungi. Ecology 1988, 69, 10–16. [Google Scholar] [CrossRef]
- Breen, J.P. Acremonium endophyte interactions with enhanced plant resistance to insects. Annu. Rev. Entomol. 1994, 39, 401–423. [Google Scholar] [CrossRef]
- Mahmood, T.; Gergerich, R.C.; Milus, E.A.; West, C.P.; Darcy, C.J. Barley yellow dwarf viruses in wheat, endophyte-infected and endophyte-free tall fescue, and other hosts in Arkansas. Plant Dis. 1993, 77, 225–228. [Google Scholar] [CrossRef]
- Wang, X.Y.; Qin, J.H.; Chen, W.; Zhou, Y.; Ren, A.Z.; Gao, Y.B. Pathogen resistant advantage of endophyte-infected over endophyte-free Leymus chinensis is strengthened by pre-drought treatment. Eur. J. Plant Pathol. 2016, 144, 477–486. [Google Scholar] [CrossRef]
- Omacini, M.; Eggers, T.; Bonkowski, M.; Gange, A.C.; Jones, T.H. Leaf endophytes affect mycorrhizal status and growth of coinfected and neighbouring plants. Funct. Ecol. 2006, 20, 226–232. [Google Scholar] [CrossRef]
- Vázquez-de-Aldana, B.R.; Zabalgogeazcoa, I.; Garcia-Ciudad, A.; Garcia-Criado, B. An Epichloë endophyte affects the competitive ability of Festuca rubra against other grassland species. Plant Soil 2013, 362, 201–213. [Google Scholar] [CrossRef]
- Wu, L.J.; Ren, A.Z.; Jing, Y.F.; Zhou, Y.; Wang, X.Y.; Qin, J.H.; Gao, Y.B. Endophytic benefit for a competitive host is neutralized by increasing ratios of infected plants. Acta Oecologica 2016, 70, 112–120. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, X.; Gao, Y.; Liu, H.; Gao, Y.B.; van der Heijden, M.G.A.; Ren, A.Z. Plant endophytes and arbuscular mycorrhizal fungi alter plant competition. Funct. Ecol. 2018, 32, 1168–1179. [Google Scholar] [CrossRef]
- Hill, N.S.; Belesky, D.P.; Stringer, W.C. Competitiveness of tall fescue as influenced by Acremomium coenophialum. Crop Sci. 1991, 31, 185–190. [Google Scholar] [CrossRef]
- Marks, S.; Clay, K.; Cheplick, G.P. Effects of fungal endophytes on interspecific and intraspecific competition in the grasses Festuca arundinacea and Lolium perenne. J. Appl. Ecol. 1991, 28, 194–204. [Google Scholar] [CrossRef]
- Malinowski, D.P.; Leuchtmann, A.; Schmidt, D.; Nösberger, J. Growth and water status in meadow fescue is affected by Neotyphodium and Phialophora species endophytes. Agron. J. 1997, 89, 673–678. [Google Scholar] [CrossRef]
- Brem, D.; Leuchtmann, A. Intraspecific competition of endophyte infected vs uninfected plants of two woodland grass species. Oikos 2002, 96, 281–290. [Google Scholar] [CrossRef]
- Takai, T.; Sanada, Y.; Yamada, T. Influence of the fungal endophyte Neotyphodium uncinatum on the persistency and competitive ability of meadow fescue (Festuca pratensis Huds.). Grassl. Sci. 2010, 56, 59–64. [Google Scholar] [CrossRef]
- Saikkonen, K.; Ruokolainen, K.; Huitu, O.; Gundel, P.E.; Piltti, T.; Hamilton, C.E.; Helander, M. Fungal endophytes help prevent weed invasions. Agric. Ecosyst. Environ. 2013, 165, 1–5. [Google Scholar] [CrossRef]
- Craig, S.; Kannadan, S.; Flory, S.L.; Seifert, E.K.; Whitney, K.D.; Rudgers, J.A. Potential for endophyte symbiosis to increase resistance of the native grass Poa alsodes to invasion by the nonnative grass Microstegium vimineum. Symbiosis 2011, 53, 17–28. [Google Scholar] [CrossRef]
- Richmond, D.; Grewal, P.; Cardina, J. Competition between Lolium perenne and Digitaria sanguinalis: Ecological consequences for harbouring an endosymbiotic fungus. J. Veg. Sci. 2003, 14, 835–840. [Google Scholar] [CrossRef]
- Faeth, S.H.; Helander, M.L.; Saikkonen, K.T. Asexual Neotyphodium endophytes in a native grass reduce competitive abilities. Ecol. Lett. 2004, 7, 304–313. [Google Scholar] [CrossRef]
- Cheplick, G.P.; Harrichandra, A.P.; Liu, A.A. Competitive outcomes depend on host genotype, but not clavicipitaceous fungal endophytes, in Lolium perenne (Poaceae). Am. J. Bot. 2014, 101, 2068–2078. [Google Scholar] [CrossRef]
- Dirihan, S.; Helander, M.L.; Saloniemi, I.; Gundel, P.E.; Saikkonen, K. Effects of systemic fungal endophytes on the performance of meadow fescue and tall fescue in mixtures with red clover. Grass Forage Sci. 2014, 70, 465–473. [Google Scholar] [CrossRef]
- Larimer, A.L.; Bever, J.D.; Clay, K. Consequences of simultaneous interactions of fungal endophytes and arbuscular mycorrhizal fungi with a shared host grass. Oikos 2012, 121, 2090–2096. [Google Scholar] [CrossRef]
- Arrieta, A.M.; Iannone, L.J.; Scervino, J.M.; Vignale, M.V.; Novas, M.V. A foliar endophyte increases the diversity of phosphorus solubilizing rhizospheric fungi and mycorrhizal colonization in the wild grass Bromus auleticus. Fungal Ecol. 2015, 17, 146–15432. [Google Scholar] [CrossRef]
- García Parisi, P.A.; Lattanzi, F.A.; Grimoldi, A.A.; Omacini, M. Multi-symbiotic systems: Functional implications of the coexistence of grass-endophyte and legume-rhizobia symbioses. Oikos 2015, 124, 553–560. [Google Scholar] [CrossRef]
- Bell-Dereske, L.; Takacs-Vesbach, C.; Kivlin, S.N.; Emery, S.M.; Rudgers, J.A. Leaf endophytic fungus interacts with precipitation to alter belowground microbial communities in primary successional dunes. FEMS Microbiol. Ecol. 2017, 93, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, X.; Qin, J.H.; Liu, H.; Chen, W.; Niu, Y.; Ren, A.Z.; Gao, Y.B. Effects of simultaneous infections of endophytic fungi and arbuscular mycorrhizal fungi on the growth of their shared host grass Achnatherum sibiricum under varying N and P supply. Fungal Ecol. 2016, 20, 56–65. [Google Scholar] [CrossRef]
- Zhu, M.J.; Ren, A.Z.; Wen, W.; Gao, Y.B. Diversity and taxonomy of endophytes from Leymus chinensis in the Inner Mongolia steppe of China. FEMS Microbiol. Lett. 2013, 340, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Latch, G.C.M.; Christensen, M.J.; Samuels, G.J. Five endophytes of Lolium and Festuca in New Zealand. Mycotaxon 1984, 167, 338–342. [Google Scholar]
- Wei, Y.K.; Gao, Y.B.; Xu, H.; Su, D.; Zhang, X.; Wang, Y.H.; Lin, F.; Chen, L.; Nie, L.Y.; Ren, A.Z. Occurrence of endophytes in grasses native to northern China. Grass Forage Sci. 2006, 61, 422–429. [Google Scholar] [CrossRef]
- de Wit, C.T. On competition. Versl. Landbouwkd. Onderz. 1960, 66, 1–82. [Google Scholar]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- McGonigle, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular-arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- McGilchrist, C.A.; Trenbath, B.R. A revised analysis of plant competition experiments. Biometrics 1971, 27, 659–671. [Google Scholar] [CrossRef]
- Aschehoug, E.T.; Metlen, K.L.; Callaway, R.M.; Newcombe, G. Fungal endophytes directly increase the competitive effects of an invasive forb. Ecology 2012, 93, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Lekberg, Y.; Bever, J.D.; Bunn, R.A.; Callaway, R.M.; Hart, M.M.; Kivlin, S.N.; Klironomos, J.; Larkin, B.G.; Maron, J.L.; Reinhart, K.O.; et al. Relative importance of competition and plant-soil feedback, their synergy, context dependency and implications for coexistence. Ecol. Lett. 2018, 21, 1268–1281. [Google Scholar] [CrossRef] [PubMed]
- Franzluebbers, A.J. Short-term responses of soil C and N fractions to tall fescue endophyte infection. Plant Soil 2006, 282, 153–164. [Google Scholar] [CrossRef]
- Jenkins, M.B.; Franzluebbers, A.J.; Humayoun, S.B. Assessing short-term responses of prokaryotic communities in bulk and rhizosphere soils to tall fescue endophyte infection. Plant Soil 2006, 289, 309–320. [Google Scholar] [CrossRef]
- Buyer, J.S.; Zuberer, D.A.; Nichols, K.A.; Alan, J.; Franzluebbers, A.J. Soil microbial community function, structure, and glomalin in response to tall fescue endophyte infection. Plant Soil 2011, 339, 401–412. [Google Scholar] [CrossRef]
- Casas, C.; Omacini, M.; Montecchia, M.S.; Correa, O.S. Soil microbial community responses to the fungal endophyte Neotyphodium in Italian ryegrass. Plant Soil 2011, 340, 347–355. [Google Scholar] [CrossRef]
- Bowatte, S.; Barrett, B.; Luscombe, C.; Hume, D.E.; Luo, D.; Theobald, P.; Newton, P.C.D. Effect of grass species and fungal endophyte on soil nitrification potential. N. Z. J. Agric. Res. 2011, 54, 275–284. [Google Scholar] [CrossRef]
- Rudgers, J.A.; Orr, S. Non-native grass alters growth of native tree species via leaf and soil microbes. J. Ecol. 2009, 97, 247–255. [Google Scholar] [CrossRef]
- Matthews, J.W.; Clay, K. Influence of fungal endophyte infection on plant-soil feedback and community interactions. Ecology 2001, 82, 500–509. [Google Scholar]
- Cripps, M.G.; Edwards, G.R.; Mckenzie, S.L. Grass species and their fungal symbionts affect subsequent forage growth. Basic Appl. Ecol. 2013, 14, 225–234. [Google Scholar] [CrossRef]
- Garcia-Parisi, P.A.; Lattanzi, F.A.; Grimoldi, A.A.; Druille, M.; Omacini, M. Three symbionts involved in interspecific plant-soil feedback: Epichloid endophytes and mycorrhizal fungi affect the performance of rhizobia-legume symbiosis. Plant Soil 2017, 412, 151–162. [Google Scholar] [CrossRef]
- Vignale, M.V.; Iannone, L.J.; Scervino, J.M.; Novas, M.V. Epichloë exudates promote in vitro and in vivo arbuscular mycorrhizal fungi development and plant growth. Plant Soil 2018, 422, 267–281. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Intraspecific Competition | Interspecific Competition | |||||||
|---|---|---|---|---|---|---|---|---|
| Monoculture | Mixture | Monoculture | Mixture | |||||
| MI | EI | EF | EI and EF | EI | EF | S | EIS | EFS |
| MF | EI | EF | EI and EF | EI | EF | S | EIS | EFS |
| Relative Yield | Mycorrhizal Colonization Rate | RII | AGR | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Intra | Inter | Intra | Inter | Intra | Inter | Intra | Inter | |||||||||
| F | P | F | P | F | P | F | P | F | P | F | P | F | P | F | P | |
| Leymus chinensis | ||||||||||||||||
| Epichloë endophyte (E) | 16.573 | 0.004 | 6.287 | 0.037 | 12.945 | 0.007 | 33.228 | <0.001 | 18.441 | 0.003 | 5.672 | 0.044 | 12.097 | 0.008 | ||
| Mycorrhiza (M) | 1.362 | 0.277 | 9.583 | 0.015 | 1.971 | 0.198 | 9.557 | 0.015 | 1.417 | 0.300 | 12.847 | 0.007 | ||||
| Competition (C) | 0.226 | 0.647 | 0.941 | 0.360 | ||||||||||||
| E×M | 1.769 | 0.220 | 2.915 | 0.126 | 2.242 | 0.173 | 2.416 | 0.159 | 10.713 | 0.011 | ||||||
| E×C | 0.115 | 0.743 | 9.150 | 0.016 | ||||||||||||
| M×C | ||||||||||||||||
| E×M×C | ||||||||||||||||
| Stipa krylovii | ||||||||||||||||
| Epichloë endophyte (E) | 7.873 | 0.023 | ||||||||||||||
| Mycorrhiza (M) | 2.243 | 0.173 | 3.102 | 0.116 | ||||||||||||
| Competition (C) | 6.605 | 0.033 | 11.634 | 0.009 | ||||||||||||
| E×M | 16.626 | 0.004 | ||||||||||||||
| M×C | 15.096 | 0.005 | ||||||||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Chen, J.; Qin, T.; Shi, X.; Gao, Y.; Ren, A. Removal of Soil Microbes Alters Interspecific Competitiveness of Epichloë Endophyte-Infected over Endophyte-Free Leymus chinensis. Microorganisms 2020, 8, 219. https://doi.org/10.3390/microorganisms8020219
Liu H, Chen J, Qin T, Shi X, Gao Y, Ren A. Removal of Soil Microbes Alters Interspecific Competitiveness of Epichloë Endophyte-Infected over Endophyte-Free Leymus chinensis. Microorganisms. 2020; 8(2):219. https://doi.org/10.3390/microorganisms8020219
Chicago/Turabian StyleLiu, Hui, Jing Chen, Tianzi Qin, Xinjian Shi, Yubao Gao, and Anzhi Ren. 2020. "Removal of Soil Microbes Alters Interspecific Competitiveness of Epichloë Endophyte-Infected over Endophyte-Free Leymus chinensis" Microorganisms 8, no. 2: 219. https://doi.org/10.3390/microorganisms8020219
APA StyleLiu, H., Chen, J., Qin, T., Shi, X., Gao, Y., & Ren, A. (2020). Removal of Soil Microbes Alters Interspecific Competitiveness of Epichloë Endophyte-Infected over Endophyte-Free Leymus chinensis. Microorganisms, 8(2), 219. https://doi.org/10.3390/microorganisms8020219