Rhizosphere Microbiome of Arid Land Medicinal Plants and Extra Cellular Enzymes Contribute to Their Abundance

 ,
, 
Abstract
1. Introduction
2. Methods
2.1. Study Site
2.2. DNA Extraction and MiSeq Sequencing
2.3. Data Processing and Analyses
2.4. Microbial Products in Rhizosphere Soils
2.5. Statistical Analysis
3. Results
3.1. Soil Variations among Three RHIZOSPHERES
3.2. Microbial Diversity in the Rhizosphere of the Three Plants
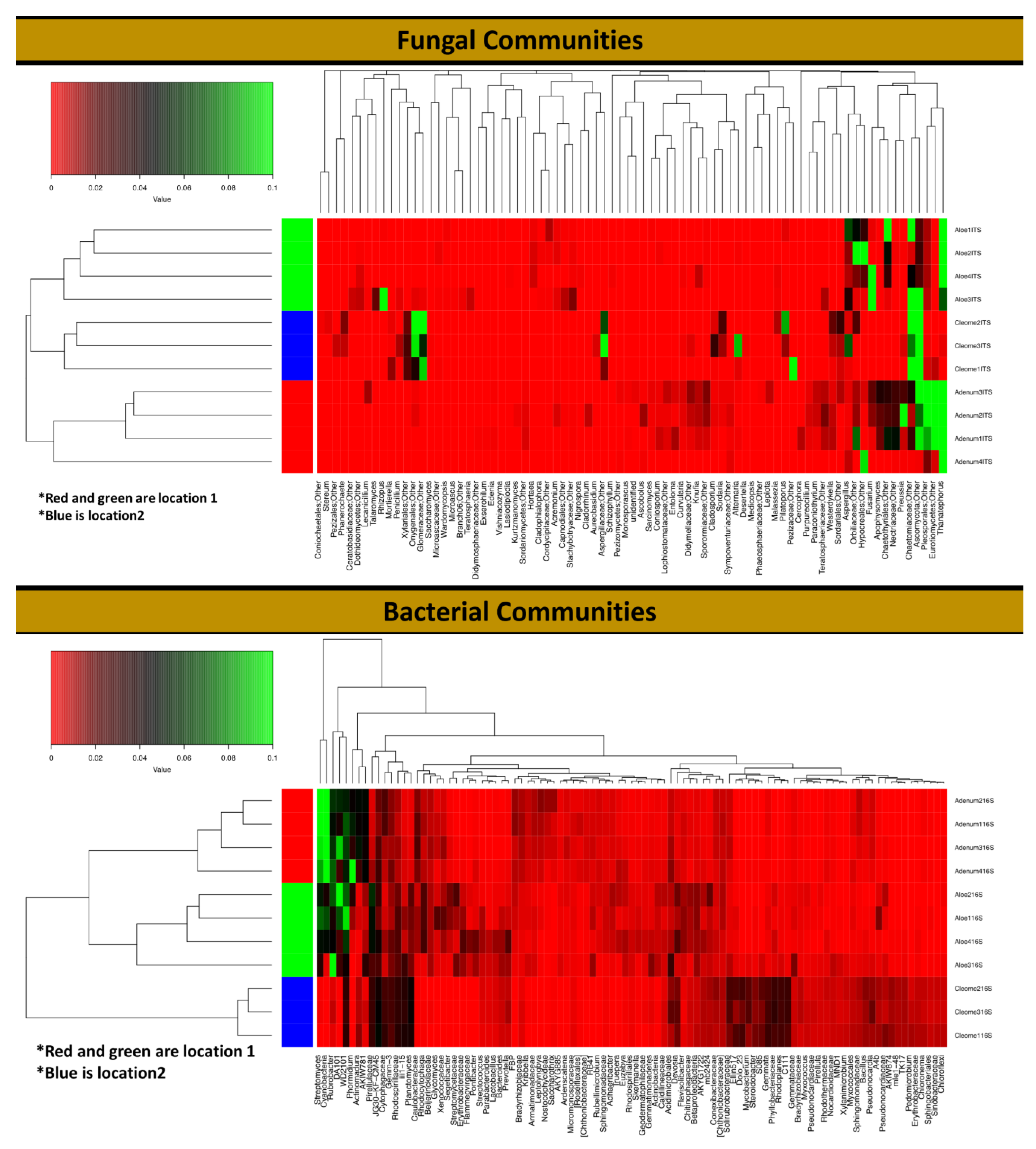
3.3. Rhizosphere Fungal Diversity
3.4. Rhizosphere Bacterial Diversity
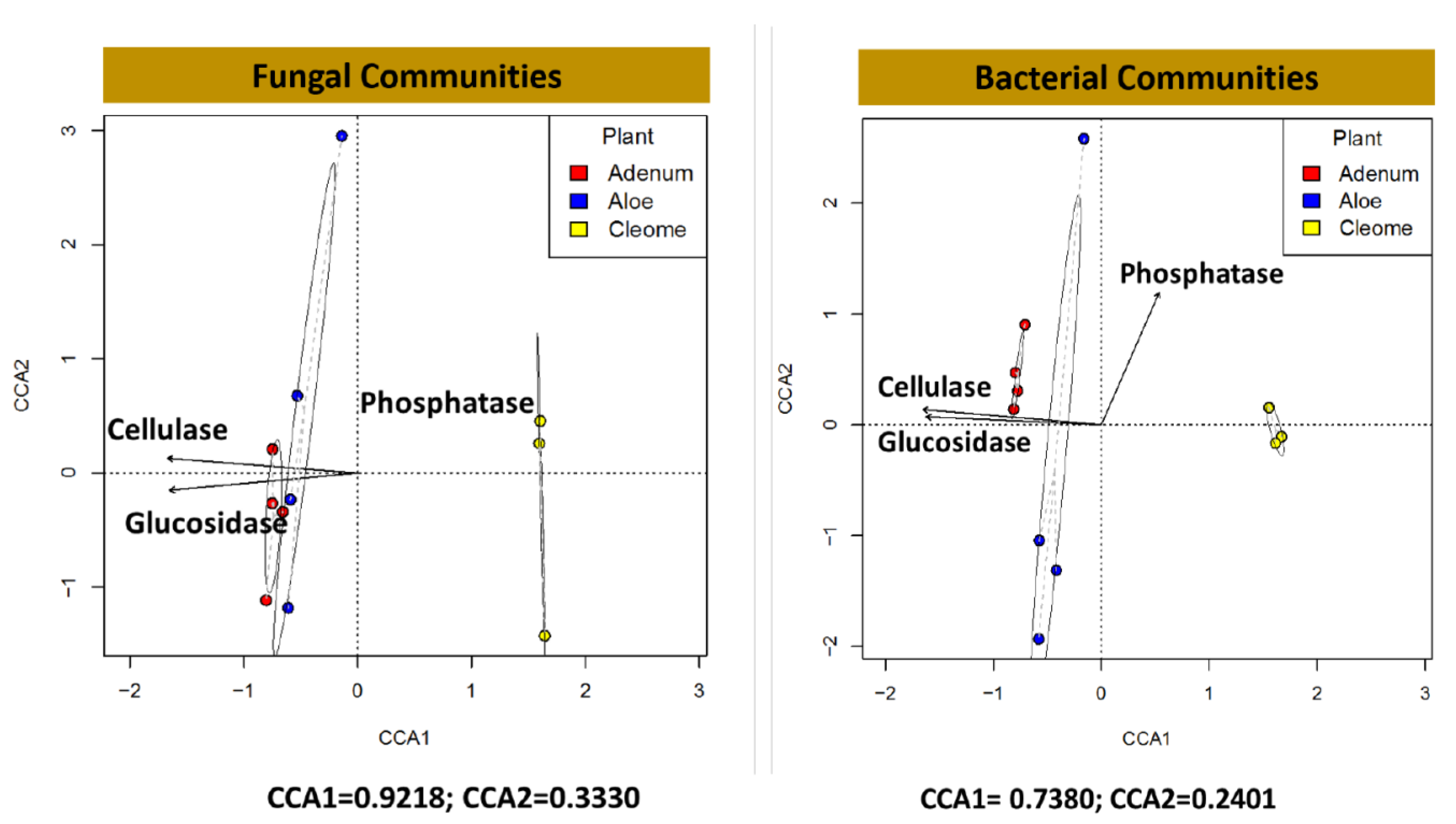
3.5. Exozymes, ACC Deaminase and IAA in the Rhizosphere
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nothers, M.; Segev, N.; Kreyling, J.; Hjazin, A.; Groner, E. Desert Vegetation Forty Years after an Oil Spill. J. Environ. Qual. 2017, 46, 568–575. [Google Scholar] [CrossRef] [PubMed]
- Ardley, J. Legumes of the Thar desert and their nitrogen fixing Ensifer symbionts. Plant Soil 2017, 410, 517–520. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Bae, H. Functional genomics and signaling events in mycorrhizal symbiosis. J. Plant Interact. 2015, 10. [Google Scholar] [CrossRef]
- Pudake, R.N.; Mehta, C.M.; Mohanta, T.K.; Sharma, S.; Varma, A.; Sharma, A.K. Expression of four phosphate transporter genes from Finger millet (Eleusine coracana L.) in response to mycorrhizal colonization and Pi stress. 3 Biotech 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Knief, C.; Delmotte, N.; Chaffron, S.; Stark, M.; Innerebner, G.; Wassmann, R.; von Mering, C.; Vorholt, J.A. Metaproteogenomic analysis of microbial communities in the phyllosphere and rhizosphere of rice. ISME J. 2012, 6, 1378–1390. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.H.; Harris, R.F. The Ecology and Biogeography of Microorganisms on Plant Surfaces. Annu. Rev. Phytopathol. 2000, 38, 145–180. [Google Scholar] [CrossRef]
- Lindow, S.E.; Brandl, M.T. Microbiology of the Phyllosphere. Appl. Environ. Microbiol. 2003, 69, 1875–1883. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef]
- Humphrey, P.T.; Nguyen, T.T.; Villalobos, M.M.; Whiteman, N.K. Diversity and abundance of phyllosphere bacteria are linked to insect herbivory. Mol. Ecol. 2014, 23, 1497–1515. [Google Scholar] [CrossRef]
- Ritpitakphong, U.; Falquet, L.; Vimoltust, A.; Berger, A.; Métraux, J.-P.; L’Haridon, F. The microbiome of the leaf surface of Arabidopsis protects against a fungal pathogen. New Phytol. 2016, 210, 1033–1043. [Google Scholar] [CrossRef]
- Strong, P.J.; Claus, H. Laccase: A Review of Its Past and Its Future in Bioremediation. Crit. Rev. Environ. Sci. Technol. 2011, 41, 373–434. [Google Scholar] [CrossRef]
- Arnosti, C. Microbial Extracellular Enzymes and their Role in Dissolved Organic Matter Cycling. In Aquatic Ecology; Findlay, S.E.G., Sinsabaugh, R.L.B.T.-A.E., Eds.; Academic Press: Burlington, NC, Canada, 2003; pp. 315–342. ISBN 978-0-12-256371-3. [Google Scholar]
- Loperena, L.; Soria, V.; Varela, H.; Lupo, S.; Bergalli, A.; Guigou, M.; Pellegrino, A.; Bernardo, A.; Calviño, A.; Rivas, F.; et al. Extracellular enzymes produced by microorganisms isolated from maritime Antarctica. World J. Microbiol. Biotechnol. 2012, 28, 2249–2256. [Google Scholar] [CrossRef]
- Leo, V.V.; Passari, A.K.; Joshi, J.B.; Mishra, V.K.; Uthandi, S.; Ramesh, N.; Gupta, V.K.; Saikia, R.; Sonawane, V.C.; Singh, B.P. A Novel Triculture System (CC3) for Simultaneous Enzyme Production and Hydrolysis of Common Grasses through Submerged Fermentation. Front. Microbiol. 2016, 7, 447. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.L.; Al-Harrasi, A.; Al-Rawahi, A.; Al-Farsi, Z.; Al-Mamari, A.; Waqas, M.; Asaf, S.; Elyassi, A.; Mabood, F.; Shin, J.-H.; et al. Endophytic Fungi from Frankincense Tree Improves Host Growth and Produces Extracellular Enzymes and Indole Acetic Acid. PLoS ONE 2016, 11, e0158207. [Google Scholar] [CrossRef] [PubMed]
- Vandenkoornhuyse, P.; Quaiser, A.; Duhamel, M.; Le Van, A.; Dufresne, A. The importance of the microbiome of the plant holobiont. New Phytol. 2015, 206, 1196–1206. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Pershina, E.V.; Ivanova, E.A.; Korvigo, I.O.; Chirak, E.L.; Sergaliev, N.H.; Abakumov, E.V.; Provorov, N.A.; Andronov, E.E. Investigation of the core microbiome in main soil types from the East European plain. Sci. Total Environ. 2018, 631–632, 1421–1430. [Google Scholar] [CrossRef]
- Warren-Rhodes, K.; Lee, K.; Archer, S.; Lacap, D.; Ng-Boyle, L.; Wttergreen, D.; Zacny, K.; Demergasso, C.; Moersch, J.; Chong, G.; et al. Soil microbial habitats in an extreme desert Mars-analogue environment. bioRxiv 2018, 269605. [Google Scholar] [CrossRef]
- Mandakovic, D.; Maldonado, J.; Pulgar, R.; Cabrera, P.; Gaete, A.; Urtuvia, V.; Seeger, M.; Cambiazo, V.; González, M. Microbiome analysis and bacterial isolation from Lejía Lake soil in Atacama Desert. Extremophiles 2018, 22, 665–673. [Google Scholar] [CrossRef]
- Marasco, R.; Rolli, E.; Ettoumi, B.; Vigani, G.; Mapelli, F.; Borin, S.; Abou-Hadid, A.F.; El-Behairy, U.A.; Sorlini, C.; Cherif, A.; et al. A Drought Resistance-Promoting Microbiome Is Selected by Root System under Desert Farming. PLoS ONE 2012, 7, e48479. [Google Scholar] [CrossRef]
- Crits-Christoph, A.; Robinson, C.K.; Barnum, T.; Fricke, W.F.; Davila, A.F.; Jedynak, B.; McKay, C.P.; DiRuggiero, J. Colonization patterns of soil microbial communities in the Atacama Desert. Microbiome 2013, 1, 28. [Google Scholar] [CrossRef] [PubMed]
- Finkel, O.M.; Delmont, T.O.; Post, A.F.; Belkin, S. Metagenomic Signatures of Bacterial Adaptation to Life in the Phyllosphere of a Salt-Secreting Desert Tree. Appl. Environ. Microbiol. 2016, 82, 2854–2861. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.; Law, A.D.; Sahib, M.R.; Pervaiz, Z.H.; Zhang, Q. Impact of root system architecture on rhizosphere and root microbiome. Rhizosphere 2018, 6, 47–51. [Google Scholar] [CrossRef]
- Pérez-Jaramillo, J.E.; Carrión, V.J.; Bosse, M.; Ferrão, L.F.V.; de Hollander, M.; Garcia, A.A.F.; Ramírez, C.A.; Mendes, R.; Raaijmakers, J.M. Linking rhizosphere microbiome composition of wild and domesticated Phaseolus vulgaris to genotypic and root phenotypic traits. ISME J. 2017, 11, 2244–2257. [Google Scholar] [CrossRef] [PubMed]
- Chaparro, J.M.; Badri, D.V.; Vivanco, J.M. Rhizosphere microbiome assemblage is affected by plant development. ISME J. 2014, 8, 790–803. [Google Scholar] [CrossRef] [PubMed]
- Coleman-Derr, D.; Desgarennes, D.; Fonseca-Garcia, C.; Gross, S.; Clingenpeel, S.; Woyke, T.; North, G.; Visel, A.; Partida-Martinez, L.P.; Tringe, S.G. Plant compartment and biogeography affect microbiome composition in cultivated and native Agave species. New Phytol. 2016, 209, 798–811. [Google Scholar] [CrossRef]
- Li, J.; Zhao, G.-Z.; Chen, H.-H.; Wang, H.-B.; Qin, S.; Zhu, W.-Y.; Xu, L.-H.; Jiang, C.-L.; Li, W.-J. Antitumour and antimicrobial activities of endophytic streptomycetes from pharmaceutical plants in rainforest. Lett. Appl. Microbiol. 2008, 47, 574–580. [Google Scholar] [CrossRef]
- Zhao, K.; Penttinen, P.; Chen, Q.; Guan, T.; Lindström, K.; Ao, X.; Zhang, L.; Zhang, X. The rhizospheres of traditional medicinal plants in Panxi, China, host a diverse selection of actinobacteria with antimicrobial properties. Appl. Microbiol. Biotechnol. 2012, 94, 1321–1335. [Google Scholar] [CrossRef]
- Zhao, K.; Penttinen, P.; Guan, T.; Xiao, J.; Chen, Q.; Xu, J.; Lindström, K.; Zhang, L.; Zhang, X.; Strobel, G.A. The Diversity and Anti-Microbial Activity of Endophytic Actinomycetes Isolated from Medicinal Plants in Panxi Plateau, China. Curr. Microbiol. 2011, 62, 182–190. [Google Scholar] [CrossRef]
- Wu, L.; Wang, H.; Zhang, Z.; Lin, R.; Zhang, Z.; Lin, W. Comparative Metaproteomic Analysis on Consecutively Rehmannia glutinosa-Monocultured Rhizosphere Soil. PLoS ONE 2011, 6, e20611. [Google Scholar] [CrossRef]
- Qi, X.; Wang, E.; Xing, M.; Zhao, W.; Chen, X. Rhizosphere and non-rhizosphere bacterial community composition of the wild medicinal plant Rumex patientia. World J. Microbiol. Biotechnol. 2012, 28, 2257–2265. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.; Ma, P.; Mu, J.; Chen, S.; Xiao, P.; Peng, Y.; Huo, L.; Xu, L.; Sun, C. De novo characterization of the root transcriptome of a traditional Chinese medicinal plant Polygonum cuspidatum. Sci. China Life Sci. 2012, 55, 452–466. [Google Scholar] [CrossRef] [PubMed]
- Akinsanya, M.A.; Goh, J.K.; Lim, S.P.; Ting, A.S.Y. Metagenomics study of endophytic bacteria in Aloe vera using next-generation technology. Genomics Data 2015, 6, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Debnath, R.; Yadav, A.; Gupta, V.K.; Singh, B.P.; Handique, P.J.; Saikia, R. Rhizospheric Bacterial Community of Endemic Rhododendron arboreum Sm. Ssp. delavayi along Eastern Himalayan Slope in Tawang. Front. Plant Sci. 2016, 7, 1345. [Google Scholar] [CrossRef] [PubMed]
- Pascual, J.; Blanco, S.; García-López, M.; García-Salamanca, A.; Bursakov, S.A.; Genilloud, O.; Bills, G.F.; Ramos, J.L.; van Dillewijn, P. Assessing Bacterial Diversity in the Rhizosphere of Thymus zygis Growing in the Sierra Nevada National Park (Spain) through Culture-Dependent and Independent Approaches. PLoS ONE 2016, 11, e0146558. [Google Scholar] [CrossRef] [PubMed]
- Favell, P. Aloe dhufarensis: The miracle Aloe of Oman. Cact. Succ. J. 1995, 67, 35–36. [Google Scholar]
- Marwah, R.G.; Fatope, M.O.; Mahrooqi, R.A.; Varma, G.B.; Abadi, H.A.; Al-Burtamani, S.K.S. Antioxidant capacity of some edible and wound healing plants in Oman. Food Chem. 2007, 101, 465–470. [Google Scholar] [CrossRef]
- Hossain, M.A. A review on Adenium obesum: A potential endemic medicinal plant in Oman. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 559–563. [Google Scholar] [CrossRef]
- Adhikari, P.; Shukla, M.K.; Mexal, J.G.; Sharma, P. Assessment of the soil physical and chemical properties of desert soils irrigated with treated wastewater using principal component analysis. Soil Sci. 2011, 176, 356–366. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef]
- Deshpande, V.; Wang, Q.; Greenfield, P.; Charleston, M.; Porras-Alfaro, A.; Kuske, C.R.; Cole, J.R.; Midgley, D.J.; Tran-Dinh, N. Fungal identification using a Bayesian classifier and the Warcup training set of internal transcribed spacer sequences. Mycologia 2016, 108, 1–5. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a Chimera-Checked 16S rRNA Gene Database and Workbench Compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069. [Google Scholar] [CrossRef] [PubMed]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. MicrobiomeAnalyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P.; et al. Ordination Methods, Diversity Analysis and Other Functions for Community and Vegetation Ecologists. Version 2.5-6. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 25 December 2019).
- Canonical Correlation Analysis. Applied Multivariate Statistical Analysis; Härdle, W., Simar, L., Eds.; Springer Berlin Heidelberg: Berlin/Heidelberg, Germany, 2007; pp. 321–330. ISBN 978-3-540-72244-1. [Google Scholar]
- Marx, M.-C.; Wood, M.; Jarvis, S.C. A microplate fluorimetric assay for the study of enzyme diversity in soils. Soil Biol. Biochem. 2001, 33, 1633–1640. [Google Scholar] [CrossRef]
- Shaharoona, B.; Arshad, M.; Zahir, Z.A. Effect of plant growth promoting rhizobacteria containing ACC-deaminase on maize (Zea mays L.) growth under axenic conditions and on nodulation in mung bean (Vigna radiata L.). Lett. Appl. Microbiol. 2006, 42, 155–159. [Google Scholar] [CrossRef]
- Lundberg, D.S.; Lebeis, S.L.; Paredes, S.H.; Yourstone, S.; Gehring, J.; Malfatti, S.; Tremblay, J.; Engelbrektson, A.; Kunin, V.; Rio, T.G.d.; et al. Defining the core Arabidopsis thaliana root microbiome. Nature 2012, 488, 86. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; de Jonge, R.; Berendsen, R.L. The Soil-Borne Supremacy. Trends Plant Sci. 2016, 21, 171–173. [Google Scholar] [CrossRef]
- Köberl, M.; Schmidt, R.; Ramadan, E.M.; Bauer, R.; Berg, G. The microbiome of medicinal plants: Diversity and importance for plant growth, quality and health. Front. Microbiol. 2013, 4, 400. [Google Scholar] [CrossRef]
- Tian, B.-Y.; Cao, Y.; Zhang, K.-Q. Metagenomic insights into communities, functions of endophytes, and their associates with infection by root-knot nematode, Meloidogyne incognita, in tomato roots. Sci. Rep. 2015, 5, 17087. [Google Scholar] [CrossRef]
- Garcia, A.; Polonio, J.C.; Polli, A.D.; Santos, C.M.; Rhoden, S.A. Rhizosphere bacteriome of the medicinal plant Sapindus saponaria L. revealed by pyrosequencing. Genet. Mol. Res. 2016, 15, gmr15049020. [Google Scholar] [CrossRef]
- Berg, G.; Rybakova, D.; Grube, M.; Köberl, M. The plant microbiome explored: Implications for experimental botany. J. Exp. Bot. 2016, 67, 995–1002. [Google Scholar] [CrossRef]
- Kembel, S.W.; Eisen, J.A.; Pollard, K.S.; Green, J.L. The Phylogenetic Diversity of Metagenomes. PLoS ONE 2011, 6, e23214. [Google Scholar] [CrossRef]
- Horton, M.W.; Bodenhausen, N.; Beilsmith, K.; Meng, D.; Muegge, B.D.; Subramanian, S.; Vetter, M.M.; Vilhjálmsson, B.J.; Nordborg, M.; Gordon, J.I.; et al. Genome-wide association study of Arabidopsis thaliana leaf microbial community. Nat. Commun. 2014, 5, 5320. [Google Scholar] [CrossRef] [PubMed]
- Lebeis, S.L.; Paredes, S.H.; Lundberg, D.S.; Breakfield, N.; Gehring, J.; McDonald, M.; Malfatti, S.; Glavina del Rio, T.; Jones, C.D.; Tringe, S.G.; et al. Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa. Science 2015, 349, 860–864. [Google Scholar] [CrossRef] [PubMed]
- Leibold, M.A.; McPeek, M.A. Coexistence of the niche and neutral perspectives in community ecology. Ecology 2006, 87, 1399–1410. [Google Scholar] [CrossRef]
- Nasseri, M.A.; Behravesh, S.; Allahresani, A. Essential oil composition of Cleome heratensis (Capparaceae) at different growing stages. Iran. Chem. Commun. 2017, 5, 364–371. [Google Scholar]
- Abubakar, S.; Akanbi, B.O.; Osuji, C.; Olajide, O.O.; Phillip, E.A. Evaluation of pharmacological potentials of Jatropha curcas Linn sap Euphobiaceae family. Pharm. Biol. Eval. 2016, 3, 334–342. [Google Scholar]
- Guo, M.; Gong, Z.; Miao, R.; Su, D.; Li, X.; Jia, C.; Zhuang, J. The influence of root exudates of maize and soybean on polycyclic aromatic hydrocarbons degradation and soil bacterial community structure. Ecol. Eng. 2017, 99, 22–30. [Google Scholar] [CrossRef]
- Rasmann, S.; Turlings, T.C.J. Root signals that mediate mutualistic interactions in the rhizosphere. Curr. Opin. Plant Biol. 2016, 32, 62–68. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Herrera, J.; Natvig, D.O.; Lipinski, K.; Sinsabaugh, R.L. Diversity and distribution of soil fungal communities in a semiarid grassland. Mycologia 2011, 103, 10–21. [Google Scholar] [CrossRef]
- Al-Yahyai, R.; Khan, M. Date Palm Status and Perspective in Oman. In Date Palm Genetic Resources and Utilization: Volume 2: Asia and Europe; Springer: Dordrecht, The Netherlands, 2015; pp. 207–240. ISBN 978-94-017-9706-1. [Google Scholar]
- Loro, M.; Valero-Jiménez, C.A.; Nozawa, S.; Márquez, L.M. Diversity and composition of fungal endophytes in semiarid Northwest Venezuela. J. Arid Environ. 2012, 85, 46–55. [Google Scholar] [CrossRef]
- Tan, Y.; Cui, Y.; Li, H.; Kuang, A.; Li, X.; Wei, Y.; Ji, X. Diversity and composition of rhizospheric soil and root endogenous bacteria in Panax notoginseng during continuous cropping practices. J. Basic Microbiol. 2017, 57, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Golinska, P.; Wypij, M.; Agarkar, G.; Rathod, D.; Dahm, H.; Rai, M. Endophytic actinobacteria of medicinal plants: Diversity and bioactivity. Antonie Van Leeuwenhoek 2015, 108, 267–289. [Google Scholar] [CrossRef] [PubMed]
- Randle-Boggis, R.J.; Helgason, T.; Sapp, M.; Ashton, P.D. Evaluating techniques for metagenome annotation using simulated sequence data. FEMS Microbiol. Ecol. 2016, 92, fiw095. [Google Scholar] [CrossRef] [PubMed]
- Quiza, L.; St-Arnaud, M.; Yergeau, E. Harnessing phytomicrobiome signaling for rhizosphere microbiome engineering. Front. Plant Sci. 2015, 6, 507. [Google Scholar] [CrossRef] [PubMed]
- Wingender, J.; Neu, T.; Flemming, H. What are bacterial extracellular polymeric substances. In Microbial Extracellular Polymeric Substances; Wingender, J.N., Thomas, R., Flemming, H.-C., Eds.; Springer: Dordrecht, The Netherlands, 1999; pp. 1–19. [Google Scholar]
- Walker, L.P.; Wilson, D.B. Enzymatic hydrolysis of cellulose: An overview. Bioresour. Technol. 1991, 36, 3–14. [Google Scholar] [CrossRef]
- Kim, Y.-M.; Saburi, W.; Yu, S.; Nakai, H.; Maneesan, J.; Kang, M.-S.; Chiba, S.; Kim, D.; Okuyama, M.; Mori, H.; et al. A novel metabolic pathway for glucose production mediated by α-glucosidase-catalyzed conversion of 1,5-anhydrofructose. J. Biol. Chem. 2012, 287, 22441–22444. [Google Scholar] [CrossRef]
- Yuan, H.; Li, N.; Lai, Y. Evaluation of in Vitro Models for Screening Alkaline Phosphatase- Mediated Bioconversion of Phosphate Ester Prodrugs. Drug Metab. Diposition 2009, 37, 1443–1447. [Google Scholar] [CrossRef]
- Razaq, M.; Zhang, P.; Shen, H. Salahuddin Influence of nitrogen and phosphorous on the growth and root morphology of Acer mono. PLoS ONE 2017, 12, e0171321. [Google Scholar] [CrossRef]
- Abdolzadeh, A.; Wang, X.; Veneklaas, E.J.; Lambers, H. Effects of phosphorus supply on growth, phosphate concentration and cluster-root formation in three Lupinus species. Ann. Bot. 2010, 105, 365–374. [Google Scholar] [CrossRef]
- Lee, M.H.; Lee, S.-W. Bioprospecting potential of the soil metagenome: Novel enzymes and bioactivities. Genom. Inform. 2013, 11, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Rasche, F.; Marco-Noales, E.; Velvis, H.; van Overbeek, L.S.; López, M.M.; van Elsas, J.D.; Sessitsch, A. Structural characteristics and plant-beneficial effects of bacteria colonizing the shoots of field grown conventional and genetically modified T4-lysozyme producing potatoes. Plant Soil 2006, 289, 123–140. [Google Scholar] [CrossRef]
- Goudjal, Y.; Toumatia, O.; Sabaou, N.; Barakate, M.; Mathieu, F.; Zitouni, A. Endophytic actinomycetes from spontaneous plants of Algerian Sahara: Indole-3-acetic acid production and tomato plants growth promoting activity. World J. Microbiol. Biotechnol. 2013, 29, 1821–1829. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | A. obesum | A. dhufarensis | C. austroarabica |
|---|---|---|---|
| Temperature (°C) | 37 | 37.3 | 41.3 |
| Moisture contents (%) | 18.7 | 19 | 10.1 |
| Clay (%) | 8.1 ± 1.01a | 6.6 ± 0.9b | 2.4 ± 0.4c |
| Sand (%) | 34 ± 2.1c | 38.5 ± 2.1a | 78.3 ± 0.6a |
| Silt (%) | 19.6 ± 1.01a | 17.6 ± 0.8a | 11.8. ± 1.5b |
| Gravel (%) | 38.3 ± 1.01a | 37.3 ± 0.8a | 2.5. ± 0.3b |
| Bulk density (%) | 4.7 ± 0.8a | 3.9 ± 0.7a | 1.1 ± 0.4b |
| Organic matter (%) | 4.2 ± 0.6a | 5.2 ± 0.4a | 0.9 ± 0.1b |
| Texture | sandy loam | sandy loam | sandy |
| EC (dSm−1) | 31 ± 1.4a | 30 ± 1.9a | 2.1 ± 0.7b |
| pH | 7.3 ± 0.8a | 7.6 ± 0.4a | 6.8 ± 0.4b |
| Nitrates (mg/kg) | 3.9 ± 0.4a | 3.4 ± 0.7a | 0.8 ± 0.2b |
| Phosphorus(mg/kg) | 2.1 ± 0.3a | 2.2 ± 0.4a | 0.4 ± 0.1b |
| Plant | Replicate | Total No. of Sequences | No. of OTUs * | Chao1 | Shannon |
|---|---|---|---|---|---|
| Fungal communities | |||||
| A. dhufarensis | 1 | 69673 | 131 | 58.5 | 1.9 |
| 2 | 48156 | 104 | 47.3 | 1.7 | |
| 3 | 26999 | 85 | 45.2 | 2.2 | |
| A. obesum | 1 | 65158 | 176 | 88.0 | 2.7 |
| 2 | 51709 | 196 | 81.5 | 3.0 | |
| 3 | 78490 | 229 | 108.0 | 3.0 | |
| C. austroarabica | 1 | 42156 | 50 | 43 | 2.4 |
| 2 | 39003 | 49 | 44 | 2.6 | |
| 3 | 33497 | 63 | 41 | 2.7 | |
| Bacterial communities | |||||
| A. dhufarensis | 1 | 150605 | 3966 | 1768.5 | 5.7 |
| 2 | 133210 | 3241 | 1690.5 | 5.7 | |
| 3 | 126210 | 4834 | 1694.7 | 6.0 | |
| A. obesum | 1 | 206761 | 3622 | 1754.2 | 5.5 |
| 2 | 176014 | 3296 | 1748.8 | 5.3 | |
| 3 | 210956 | 3717 | 1788.0 | 5.5 | |
| C. austroarabica | 1 | 152820 | 2812 | 1232.4 | 6.0 |
| 2 | 151785 | 2854 | 1232.7 | 6.0 | |
| 3 | 149156 | 4616 | 1375.7 | 6.1 | |
| Assays | Enzyme Properties | Plant Species | ||||
|---|---|---|---|---|---|---|
| Function | Substrate | Conc. (μM) | A. dhufarensis | A. obesum | C. austroarabica | |
| Cellulase (1,4- β –cellobiosidase; (μmol h−1 g−1) | Cellulose to disaccharide | 4-MUB-phosphate 3.1.3.1 | 10–100 | 157.1 ± 1.2 a | 134.2 ± 1.0 b | 105.8 ± 1.0 c |
| Phosphatase (μmol h−1 g−1) | Phosphomonoesters to phosphate | 4-MUB-β-D-cellobioside 3.2.1.91 | 10–100 | 9.6 ± 0.2 b | 11.4 ± 0.9 a | 5.2 ± 0.6 c |
| Glucosidase (β-1,4-glucosidase; (μmol h−1 g−1) | Cellulose to glucose | 4-MUB-β-D-glucopyranoside 3.2.1.21 | 10–100 | 16.9 ± 0.8 a | 13.8 ± 0.4 b | 5.5 ± 0.7 c |
| Esterase (μmol h−1 g−1) | Acid to alcohol (hydrolyses) | 4-Methylumbelliferyl butyrate | 10–100 | 152.0 ± 1.01 a | 131.1 ± 2.0 b | 117.1 ± 2.7 c |
| IAA (µmol/mL) | Plant root development | - | 100 | 129.5 ± 2.8 b | 159.8 ± 2.9 a | 122.6 ± 3.4 bc |
| ACC deaminase (nmol α-ketobutyrate mg−1 h−1) | Lowering plant ethylene levels | ACC | 100 | 181.2 ± 2.8 a | 98.1 ± 2.8 b | 39.8 ± 2.8 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, A.L.; Asaf, S.; M. Abed, R.M.; Ning Chai, Y.; N. Al-Rawahi, A.; Mohanta, T.K.; Al-Rawahi, A.; Schachtman, D.P.; Al-Harrasi, A. Rhizosphere Microbiome of Arid Land Medicinal Plants and Extra Cellular Enzymes Contribute to Their Abundance. Microorganisms 2020, 8, 213. https://doi.org/10.3390/microorganisms8020213
Khan AL, Asaf S, M. Abed RM, Ning Chai Y, N. Al-Rawahi A, Mohanta TK, Al-Rawahi A, Schachtman DP, Al-Harrasi A. Rhizosphere Microbiome of Arid Land Medicinal Plants and Extra Cellular Enzymes Contribute to Their Abundance. Microorganisms. 2020; 8(2):213. https://doi.org/10.3390/microorganisms8020213
Chicago/Turabian StyleKhan, Abdul Latif, Sajjad Asaf, Raeid M. M. Abed, Yen Ning Chai, Ahmed N. Al-Rawahi, Tapan Kumar Mohanta, Ahmed Al-Rawahi, Daniel P. Schachtman, and Ahmed Al-Harrasi. 2020. "Rhizosphere Microbiome of Arid Land Medicinal Plants and Extra Cellular Enzymes Contribute to Their Abundance" Microorganisms 8, no. 2: 213. https://doi.org/10.3390/microorganisms8020213
APA StyleKhan, A. L., Asaf, S., M. Abed, R. M., Ning Chai, Y., N. Al-Rawahi, A., Mohanta, T. K., Al-Rawahi, A., Schachtman, D. P., & Al-Harrasi, A. (2020). Rhizosphere Microbiome of Arid Land Medicinal Plants and Extra Cellular Enzymes Contribute to Their Abundance. Microorganisms, 8(2), 213. https://doi.org/10.3390/microorganisms8020213