Antibiotic Sensitivity Screening of Klebsiella spp. and Raoultella spp. Isolated from Marine Bivalve Molluscs Reveal Presence of CTX-M-Producing K. pneumoniae

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Sample Preparation
2.3. Isolation and Identification of Presumptive Klebsiella spp.
2.4. Enumeration of E. coli
2.5. Antimicrobial Susceptibility Testing
2.6. Whole Genome Sequencing and Sequence Analysis
2.7. PCR Amplification of blaSHV
2.8. Filter Conjugation
3. Results
3.1. Distribution of Klebsiella spp. and Raoultella spp. in Marine Bivalves
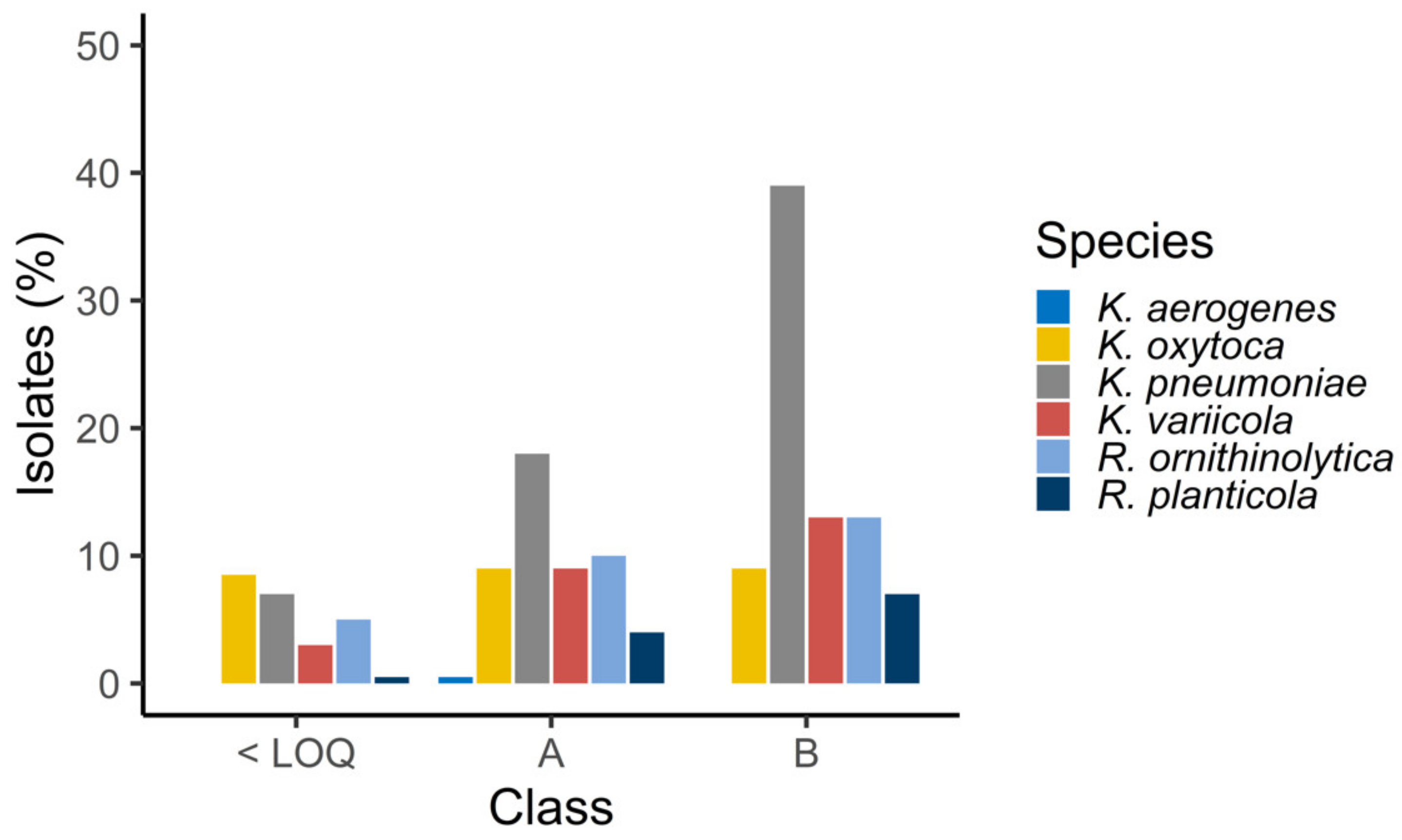
3.2. Antimicrobial Susceptibility Patterns of Klebsiella spp. and Raoultella spp.
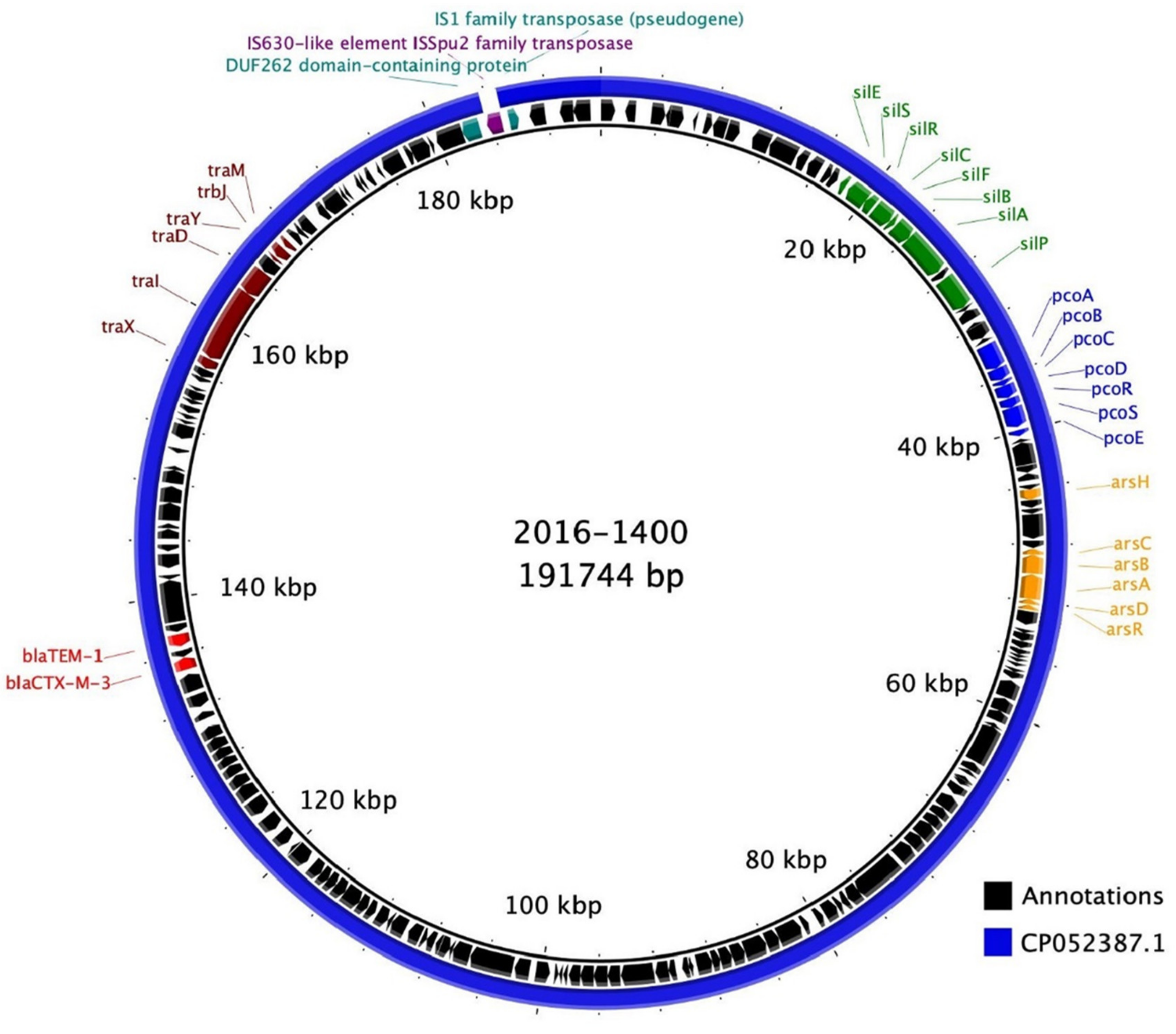
3.3. Genome Sequencing
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- Podschun, R.; Ullmann, U. Klebsiella spp. as nosocomial pathogens: Epidemiology, taxonomy, typing methods and pathogenicity factors. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Medina, N.; Barrios-Camacho, H.; Duran-Bedolla, J.; Garza-Ramos, U. Klebsiella variicola: An emerging pathogen in humans. Emerg. Microbes Infect. 2019, 8, 973–988. [Google Scholar] [CrossRef] [PubMed]
- Breurec, S.; Melot, B.; Hoen, B.; Passet, V.; Schepers, K.; Bastian, S.; Brisse, S. Liver abscess caused by infection with community-acquired Klebsiella quasipneumoniae subsp. quasipneumoniae. Emerg. Infect. Dis. 2016, 22, 529–531. [Google Scholar] [CrossRef] [PubMed]
- Shankar, C.; Veeraraghavan, B.; Nabarro, L.E.B.; Ravi, R.; Ragupathi, N.K.D.; Rupali, P. Whole genome analysis of hypervirulent Klebsiella pneumoniae isolates from community and hospital acquired bloodstream infection. BMC Microbiol. 2018, 18, 6. [Google Scholar] [CrossRef]
- Brisse, S.; Grimont, F.; Grimont, P.A.D. The Genus Klebsiella. In Proteobacteria: Gamma Subclass; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 6. [Google Scholar]
- Drancourt, M.; Bollet, C.; Carta, A.; Rousselier, P. Phylogenetic analyses of Klebsiella species delineate Klebsiella and Raoultella gen. nov., with description of Raoultella ornithinolytica comb. nov., Raoultella terrigena comb. nov. and Raoultella planticola comb. nov. Int. J. Syst. Evol. Microbiol. 2001, 51, 925–932. [Google Scholar] [CrossRef]
- Sękowska, A. Raoultella spp.—Clinical significance, infections and susceptibility to antibiotics. Folia Microbiol. 2017, 62, 221–227. [Google Scholar] [CrossRef]
- Brisse, S.; Passet, V.; Grimont, P.A.D. Description of Klebsiella quasipneumoniae sp. nov., isolated from human infections, with two subspecies, Klebsiella quasipneumoniae subsp. quasipneumoniae subsp. nov. and Klebsiella quasipneumoniae subsp. similipneumoniae subsp. nov. and demonstration that Klebsiella singaporensis is a junior heterotypic synonym of Klebsiella variicola. Int. J. Syst. Evol. Microbiol. 2014, 64, 3146–3152. [Google Scholar] [CrossRef]
- Rodrigues, C.; Passet, V.; Rakotondrasoa, A.; Diallo, T.A.; Criscuolo, A.; Brisse, S. Description of Klebsiella africanensis sp. nov., Klebsiella variicola subsp. tropicalensis subsp. nov. and Klebsiella variicola subsp. variicola subsp. nov. Res. Microbiol. 2019, 170, 165–170. [Google Scholar] [CrossRef]
- Wyres, K.L.; Lam, M.M.C.; Holt, K.E. Population genomics of Klebsiella pneumoniae. Nat. Rev. Genet. 2020, 18, 344–359. [Google Scholar] [CrossRef]
- Long, S.W.; Linson, S.E.; Saavedra, M.O.; Cantu, C.; Davis, J.J.; Brettin, T.; Olsen, R.J. Whole-genome sequencing of a human clinical isolate of the novel species Klebsiella quasivariicola sp. nov. Genome Announc. 2017, 5, e01057–e02017. [Google Scholar] [CrossRef]
- Holt, K.E.; Wyres, K.L. Klebsiella pneumoniae as a key trafficker of drug resistance genes from environmental to clinically important bacteria. Curr. Opin. Microbiol. 2018, 45, 131–139. [Google Scholar] [CrossRef]
- ECDC. Surveillance of Antimicrobial Resistance in Europe: Annual Report of the European Antimicrobial Resistance Surveillance Network (EARS-Net) 2018; ECDC: Stockholm, Sweden, 2019; Available online: https://www.ecdc.europa.eu/en/publications-data/surveillance-antimicrobial-resistance-europe-2018 (accessed on 17 October 2020).
- Bevan, E.R.; Jones, A.M.; Hawkey, P.M. Global epidemiology of CTX-M β-lactamases: Temporal and geographical shifts in genotype. J. Antimicrob. Chemother. 2017, 72, 2145–2155. [Google Scholar] [CrossRef] [PubMed]
- NORM/NORM-VET. NORM/NORM-VET: Usage of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Norway 2018; Norwegian Surveillance System for Antibiotic Resistance in Microbes/Norwegian Veterinary Institute/Norwegian Institute of Public Health: Tromsø/Oslo, Norway, 2018; Available online: https://www.vetinst.no/en/surveillance-programmes/norm-norm-vet-report (accessed on 17 October 2020).
- NORM/NORM-VET. NORM/NORM-VET: Usage of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Norway 2010; Norwegian Surveillance System for Antibiotic Resistance in Microbes/Norwegian Veterinary Institute/Norwegian Institute of Public Health: Tromsø/Oslo, Norway, 2010; Available online: https://www.vetinst.no/en/surveillance-programmes/norm-norm-vet-report (accessed on 17 October 2020).
- Martin, R.M.; Bachman, M. Colonization, infection and the accessory genome of Klebsiella pneumoniae. Front. Cell. Infect. Microbiol. 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Holt, K.E. Klebsiella pneumoniae Population genomics and antimicrobial-resistant clones. Trends Microbiol. 2016, 24, 944–956. [Google Scholar] [CrossRef] [PubMed]
- Von Wintersdorff, C.J.H.; Penders, J.; Van Niekerk, J.M.; Mills, N.D.; Majumder, S.; Van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef]
- Martínez, J. Antibiotics and antibiotic resistance genes in natural environments. Science 2008, 321, 365–367. [Google Scholar] [CrossRef]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Buergmann, H.; Sørum, H.; Norström, M.; Pons, M.-N.; et al. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Genet. 2015, 13, 310–317. [Google Scholar] [CrossRef]
- Grevskott, D.H.; Svanevik, C.S.; Sunde, M.; Wester, A.L.; Lunestad, B.T. Marine bivalve mollusks as possible indicators of multidrug-resistant Escherichia coli and other species of the Enterobacteriaceae family. Front. Microbiol. 2017, 8, 24. [Google Scholar] [CrossRef]
- Jørgensen, S.; Søraas, A.V.; Arnesen, L.S.; Leegaard, T.M.; Sundsfjord, A.; Jenum, P.A. A comparison of extended spectrum β-lactamase producing Escherichia coli from clinical, recreational water and wastewater samples associated in time and location. PLoS ONE 2017, 12, e0186576. [Google Scholar] [CrossRef]
- Bernard, F.R. Uptake and elimination of coliform bacteria by four marine bivalve mollusks. Can. J. Fish. Aquat. Sci. 1989, 46, 1592–1599. [Google Scholar] [CrossRef]
- Kibria, G.; Hossain, M.; Mallick, D.; Lau, T.; Wu, R. Monitoring of metal pollution in waterways across Bangladesh and ecological and public health implications of pollution. Chemosphere 2016, 165, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Runcharoen, C.; Moradigaravand, D.; Blane, B.; Paksanont, S.; Thammachote, J.; Anun, S.; Parkhill, J.; Chantratita, N.; Peacock, S.J. Whole genome sequencing reveals high-resolution epidemiological links between clinical and environmental Klebsiella pneumoniae. Genome Med. 2017, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Struve, C.; Krogfelt, K.A. Pathogenic potential of environmental Klebsiella pneumoniae isolates. Environ. Microbiol. 2004, 6, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Podschun, R.; Pietsch, S.; Höller, C.; Ullmann, U. Incidence of Klebsiella species in surface waters and their expression of virulence factors. Appl. Environ. Microbiol. 2001, 67, 3325–3327. [Google Scholar] [CrossRef] [PubMed]
- Van Kregten, E.; Westerdaal, N.A.; Willers, J.M. New, simple medium for selective recovery of Klebsiella pneumoniae and Klebsiella oxytoca from human feces. J. Clin. Microbiol. 1984, 20, 936–941. [Google Scholar] [CrossRef] [PubMed]
- ISO. ISO16649-3. Microbiology of the Food Chain–Horizontal Method for the Enumeration of Beta-Glucuronidase-Positive Escherichia Coli–Part 3: Detection and Most Probable Number Technique Using 5-bromo-4-chloro-3-indolyl-β-D-Glucuronide; International Organization for Standardization (ISO): Geneva, Switzerland, 2015. [Google Scholar]
- EU. Regulation (EC) No 854/2004 of the European Parliament and of the Council of 29 April 2004 laying down specific rules for the organisation of official controls on products of animal origin intended for human consumption. Off. J. Eur. Communities 2004, 24, 83–127. [Google Scholar]
- EUCAST. Antimicrobial Susceptibility Testing-EUCAST Disk Diffusion Method. Available online: https://www.eucast.org (accessed on 9 January 2020).
- EUCAST. Breakpoint Tables for Interpreation of MICs and Zone Diameters. Available online: https://www.eucast.org/ (accessed on 10 July 2020).
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.; Carmeli, Y.; Falagas, M.; Giske, C.; Harbarth, S.J.; Hindler, J.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Feldgarden, M.; Brover, V.; Haft, D.H.; Prasad, A.B.; Slotta, D.J.; Tolstoy, I.; Tyson, G.H.; Zhao, S.; Hsu, C.-H.; McDermott, P.F.; et al. Validating the AMRFinder tool and resistance gene database by using antimicrobial resistance genotype-phenotype correlations in a collection of isolates. Antimicrob. Agents Chemother. 2019, 63, e00483-19. [Google Scholar] [CrossRef]
- Chen, L.; Zheng, D.; Liu, B.; Yang, J.; Jin, Q. VFDB 2016: Hierarchical and refined dataset for big data analysis—10 years on. Nucleic Acids Res. 2016, 44, D694–D697. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Alikhan, N.-F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.S.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Paterson, D.L.; Hujer, K.M.; Hujer, A.M.; Yeiser, B.; Bonomo, M.D.; Rice, L.B.; Bonomo, R.A. Extended-spectrum β-lactamases in Klebsiella pneumoniae bloodstream isolates from seven countries: Dominance and widespread prevalence of SHV- and CTX-M-type β-lactamases. Antimicrob. Agents Chemother. 2003, 47, 3554–3560. [Google Scholar] [CrossRef]
- Jutkina, J.; Rutgersson, C.; Flach, C.-F.; Larsson, D.G.J. An assay for determining minimal concentrations of antibiotics that drive horizontal transfer of resistance. Sci. Total. Environ. 2016, 584, 131–138. [Google Scholar] [CrossRef]
- Gniadkowski, M.; Schneider, I.; Pal/ucha, A.; Jungwirth, R.; Mikiewicz, B.; Bauernfeind, A. Cefotaxime-resistant enterobacteriaceae isolates from a hospital in Warsaw, Poland: Identification of a new CTX-M-3 cefotaxime-hydrolyzing β-lactamase that is closely related to the CTX-M-1/MEN-1 enzyme. Antimicrob. Agents Chemother. 1998, 42, 827–832. [Google Scholar] [CrossRef]
- Cantón, R.; Egonzalez-Alba, J.M.; Galan, J.-C. CTX-M enzymes: Origin and diffusion. Front. Microbiol. 2012, 3, 110. [Google Scholar] [CrossRef]
- Klein, E.Y.; Van Boeckel, T.P.; Martinez, E.M.; Pant, S.; Gandra, S.; Levin, S.A.; Goossens, H.; Laxminarayan, R. Global increase and geographic convergence in antibiotic consumption between 2000 and 2015. Proc. Natl. Acad. Sci. USA 2018, 115, E3463–E3470. [Google Scholar] [CrossRef]
- Rawat, D.; Nair, D. Extended-spectrum β-lactamases in gram negative bacteria. J. Glob. Infect. Dis. 2010, 2, 263–274. [Google Scholar] [CrossRef]
- Bush, K.; Jacoby, G.A. Updated functional classification of β-lactamases. Antimicrob. Agents Chemother. 2009, 54, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Bialek-Davenet, S.; Criscuolo, A.; Ailloud, F.; Passet, V.; Jones, L.; Delannoy-Vieillard, A.-S.; Garin, B.; Le Hello, S.; Arlet, G.; Nicolas-Chanoine, M.-H.; et al. Genomic definition of hypervirulent and multidrug-resistant Klebsiella pneumoniae clonal groups. Emerg. Infect. Dis. 2014, 20, 1812–1820. [Google Scholar] [CrossRef]
- Rodrigues, C.; D’Humières, C.; Papin, G.; Passet, V.; Ruppé, E.; Brisse, S. Community-acquired infection caused by the uncommon hypervirulent Klebsiella pneumoniae ST66-K2 lineage. Microb. Genom. 2020, 6, e000419. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Green, N.W.; Schøyen, M.; Hjermann, D.Ø.; Øxnevad, S.; Ruus, A.; Lusher, A.; Beylich, B.; Lund, E.; Tveiten, L.A.; Håvardstun, J.; et al. Contaminants in Coastal Waters of Norway 2017. Available online: https://www.miljodirektoratet.no/globalassets/publikasjoner/m1120/m1120.pdf (accessed on 17 October 2020).
- Cubero, M.; Grau, I.; Tubau, F.; Pallarés, R.; Dominguez, M.; Liñares, J.; Ardanuy, C. Hypervirulent Klebsiella pneumoniae clones causing bacteraemia in adults in a teaching hospital in Barcelona, Spain (2007–2013). Clin. Microbiol. Infect. 2016, 22, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Aires-De-Sousa, M.; Kudyba, P.; Kieffer, N.; Nordmann, P. Screening and characterization of multidrug-resistant gram-negative bacteria from a remote African area, São Tomé and Príncipe. Antimicrob. Agents Chemother. 2018, 62, 01018–01021. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.K.; Kaya, H.; Roer, L.; Hansen, F.; Skovgaard, S.; Justesen, U.S.; Hansen, D.S.; Andersen, L.P.; Knudsen, J.D.; Røder, B.L.; et al. Molecular characterization of Danish ESBL/AmpC-producing Klebsiella pneumoniae from bloodstream infections. J. Glob. Antimicrob. Resist. 2020. [Google Scholar] [CrossRef] [PubMed]
- Higashino, M.; Murata, M.; Morinaga, Y.; Akamatsu, N.; Matsuda, J.; Takeda, K.; Kaku, N.; Kosai, K.; Uno, N.; Hasegawa, H.; et al. Fluoroquinolone resistance in extended-spectrum β-lactamase-producing Klebsiella pneumoniae in a Japanese tertiary hospital: Silent shifting to CTX-M-15-producing K. pneumoniae. J. Med Microbiol. 2017, 66, 1476–1482. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.-J.; Fang, H.-C.; Lin, T.-L.; Hsieh, P.-F.; Tsai, F.-C.; Keynan, Y.; Wang, J.-T. Capsular polysaccharide synthesis regions in Klebsiella pneumoniae serotype K57 and a new capsular serotype. J. Clin. Microbiol. 2008, 46, 2231–2240. [Google Scholar] [CrossRef]
- Hsu, C.-R.; Liao, C.-H.; Lin, T.-L.; Yang, H.-R.; Yang, F.-L.; Hsieh, P.-F.; Wu, S.-H.; Wang, J.-T. Identification of a capsular variant and characterization of capsular acetylation in Klebsiella pneumoniae PLA-associated type K. Sci. Rep. 2016, 6, 31946. [Google Scholar] [CrossRef]
- Lepuschitz, S.; Schill, S.; Stoeger, A.; Pekard-Amenitsch, S.; Huhulescu, S.; Inreiter, N.; Hartl, R.; Kerschner, H.; Sorschag, S.; Springer, B.; et al. Whole genome sequencing reveals resemblance between ESBL-producing and carbapenem resistant Klebsiella pneumoniae isolates from Austrian rivers and clinical isolates from hospitals. Sci. Total. Environ. 2019, 662, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Håkonsholm, F.; Lunestad, B.T.; Sánchez, J.R.A.; Martinez-Urtaza, J.; Marathe, N.P.; Svanevik, C.S. Vibrios from the Norwegian marine environment: Characterization of associated antibiotic resistance and virulence genes. Microbiology 2020, 9, e1093. [Google Scholar] [CrossRef]
- Grevskott, D.H.; Salvà-Serra, F.; Moore, E.R.B.; Marathe, N.P. Nanopore sequencing reveals genomic map of CTX-M-type extended-spectrum β-lactamases carried by Escherichia coli strains isolated from blue mussels (Mytilus edulis) in Norway. BMC Microbiol. 2020, 20, 10. [Google Scholar] [CrossRef] [PubMed]
- Jelic, M.; Hrenović, J.; Dekić, S.; Goić-Barišić, I.; Andrašević, A.T. First evidence of KPC-producing ST258 Klebsiella pneumoniae in river water. J. Hosp. Infect. 2019, 103, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, T.; Cantamessa, R.; Melo, L.; Fernandes, M.R.; Fraga, E.; Dropa, M.; Sato, M.I.; Cerdeira, L.; Lincopan, N. International high-risk clones of Klebsiella pneumoniae KPC-2/CC258 and Escherichia coli CTX-M-15/CC10 in urban lake waters. Sci. Total. Environ. 2017, 598, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Piedra-Carrasco, N.; Fàbrega, A.; Calero-Cáceres, W.; Cornejo-Sánchez, T.; Brown-Jaque, M.; Mir-Cros, A.; Muniesa, M.; González-López, J.J. Carbapenemase-producing enterobacteriaceae recovered from a Spanish river ecosystem. PLoS ONE 2017, 12, e0175246. [Google Scholar] [CrossRef] [PubMed]
- Ponce-Alonso, M.; Rodríguez-Rojas, L.; Del Campo, R.; Cantón, R.; Morosini, M.-I. Comparison of different methods for identification of species of the genus Raoultella: Report of 11 cases of Raoultella causing bacteraemia and literature review. Clin. Microbiol. Infect. 2016, 22, 252–257. [Google Scholar] [CrossRef]
- Babini, G.S.; Livermore, D.M. Are SHV β-lactamases universal in Klebsiella pneumoniae? Antimicrob. Agents Chemother. 2000, 44, 2230. [Google Scholar] [CrossRef]
- Potter, R.F.; Lainhart, W.; Twentyman, J.; Wallace, M.A.; Wang, B.; Burnham, C.-A.D.; Rosen, D.A.; Dantas, G. Population structure, antibiotic resistance and uropathogenicity of Klebsiella variicola. mBio 2018, 9, e02481–e03018. [Google Scholar] [CrossRef]
- Hartantyo, S.H.P.; Chau, M.L.; Koh, T.H.; Yap, M.; Yi, T.; Cao, D.Y.H.; Gutiȳrrez, R.A.; Ng, L.C. Foodborne Klebsiella pneumoniae: Virulence potential, antibiotic resistance and risks to food safety. J. Food Prot. 2020, 83, 1096–1103. [Google Scholar] [CrossRef]
- Matsen, J.M.; Spindler, J.A.; Blosser, R.O. Characterization of Klebsiella isolates from natural receiving waters and comparison with human isolates. Appl. Microbiol. 1974, 28, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zhang, F.; Zhang, W.; Chen, X.; Zhao, Y.; Ma, J.; Bao, L.; Song, W.; Ohsugi, T.; Urano, T.; et al. Differential expression of blaSHV related to susceptibility to ampicillin in Klebsiella pneumoniae. Int. J. Antimicrob. Agents 2007, 29, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Imhoff, J.F. Enterobacteriales. In Bergey’s Manual® of Systematic Bacteriology: Volume Two the Proteobacteria Part B the Gammaproteobacteria; Brenner, D.J., Krieg, N.R., Staley, J.T., Garrity, G.M., Boone, D.R., De Vos, P., Goodfellow, M., Rainey, F.A., Schleifer, K.-H., Eds.; Springer: Boston, MA, USA, 2005; pp. 587–850. [Google Scholar]
- Bagley, S.T.; Seidler, R.J.; Brenner, D.J. Klebsiella planticola sp. nov.: A new species of enterobacteriaceae found primarily in nonclinical environments. Curr. Microbiol. 1981, 6, 105–109. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Bivalve Species | |||||
|---|---|---|---|---|---|
| Species | M. edulis (n = 384) | P. maximus (n = 24) | C. gigas (n = 48) | P. rhomboides (n = 2) | Total No. Isolates |
| K. pneumoniae | 70 | 2 | 5 | 1 | 78 |
| K. oxytoca | 40 | 0 | 1 | 0 | 41 |
| K. variicola | 29 | 1 | 3 | 0 | 33 |
| K. aerogenes | 1 | 0 | 0 | 0 | 1 |
| R. ornithinolytica | 25 | 5 | 8 | 0 | 38 |
| R. planticola | 12 | 0 | 1 | 0 | 13 |
| Antibacterial Agent | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | AMP | MEL | AMC | TZP | CHL | GEN | CIP | NIT | SXT | TET | TGC | CTX | CAZ | FOX | CXM | ATM | MEM |
| K. pneumoniae (n = 78) | 74 | 0 | 3 | 3 * | 2 | 0 | 1 | 2 | 2 | 3 | 0 | 1 | 0 | 0 | 1 | 1 * | 0 |
| K. oxytoca (n = 41) | 39 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| K. variicola (n = 33) | 26 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| K. aerogenes (n = 1) | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 |
| R. ornithinolytica (n = 38) | 35 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| R. planticola (n = 13) | 13 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Isolate | ARGs | HRGs | VGs | Accession nos. |
|---|---|---|---|---|
| 2016-1400 | blaCTX-M-3, blaTEM-1, oqxA, oqxB, fosA, erm(D) | pcoA, pcoB, pcoC, pcoD, pcoR, pcoS, pcoE, arsB, arsA, arsD, arsR, arsC, arsH, SilE, SilS, SilR, SilC, SilF, SilB, SilA, SilP | ent, fep, ecp, mgt, ompA | CP065034, CP065035 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Håkonsholm, F.; Hetland, M.A.K.; Svanevik, C.S.; Sundsfjord, A.; Lunestad, B.T.; Marathe, N.P. Antibiotic Sensitivity Screening of Klebsiella spp. and Raoultella spp. Isolated from Marine Bivalve Molluscs Reveal Presence of CTX-M-Producing K. pneumoniae. Microorganisms 2020, 8, 1909. https://doi.org/10.3390/microorganisms8121909
Håkonsholm F, Hetland MAK, Svanevik CS, Sundsfjord A, Lunestad BT, Marathe NP. Antibiotic Sensitivity Screening of Klebsiella spp. and Raoultella spp. Isolated from Marine Bivalve Molluscs Reveal Presence of CTX-M-Producing K. pneumoniae. Microorganisms. 2020; 8(12):1909. https://doi.org/10.3390/microorganisms8121909
Chicago/Turabian StyleHåkonsholm, Fredrik, Marit A. K. Hetland, Cecilie S. Svanevik, Arnfinn Sundsfjord, Bjørn Tore Lunestad, and Nachiket P. Marathe. 2020. "Antibiotic Sensitivity Screening of Klebsiella spp. and Raoultella spp. Isolated from Marine Bivalve Molluscs Reveal Presence of CTX-M-Producing K. pneumoniae" Microorganisms 8, no. 12: 1909. https://doi.org/10.3390/microorganisms8121909
APA StyleHåkonsholm, F., Hetland, M. A. K., Svanevik, C. S., Sundsfjord, A., Lunestad, B. T., & Marathe, N. P. (2020). Antibiotic Sensitivity Screening of Klebsiella spp. and Raoultella spp. Isolated from Marine Bivalve Molluscs Reveal Presence of CTX-M-Producing K. pneumoniae. Microorganisms, 8(12), 1909. https://doi.org/10.3390/microorganisms8121909