From the Ocean to the Lab—Assessing Iron Limitation in Cyanobacteria: An Interface Paper

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. The History of Iron Limitation Research
3. The Role and Distribution of Iron in Marine Ecosystems
3.1. Sources and Distribution of Iron
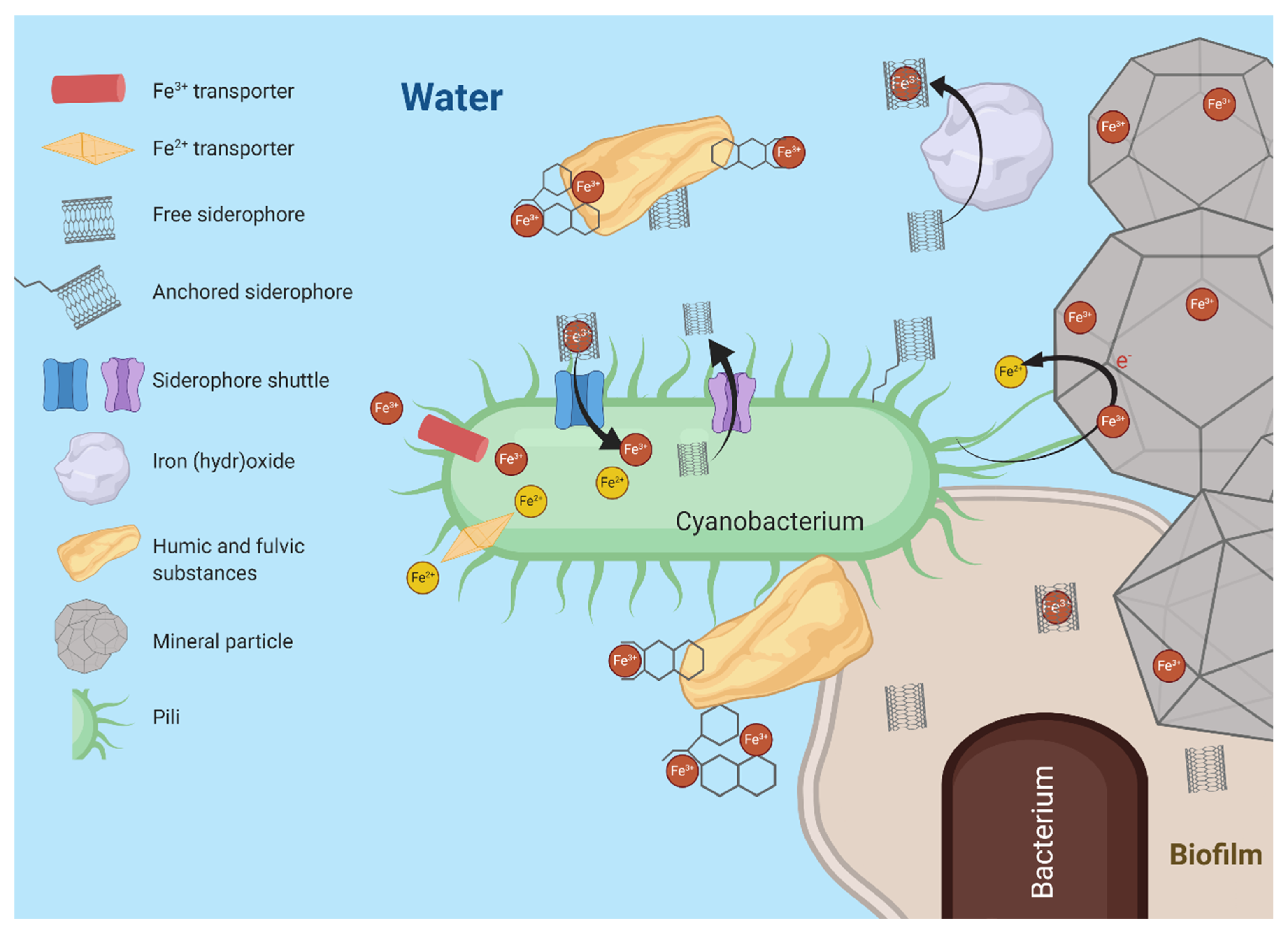
3.2. Marine Cyanobacteria in an Iron Limited World
4. From the Ocean to the Lab
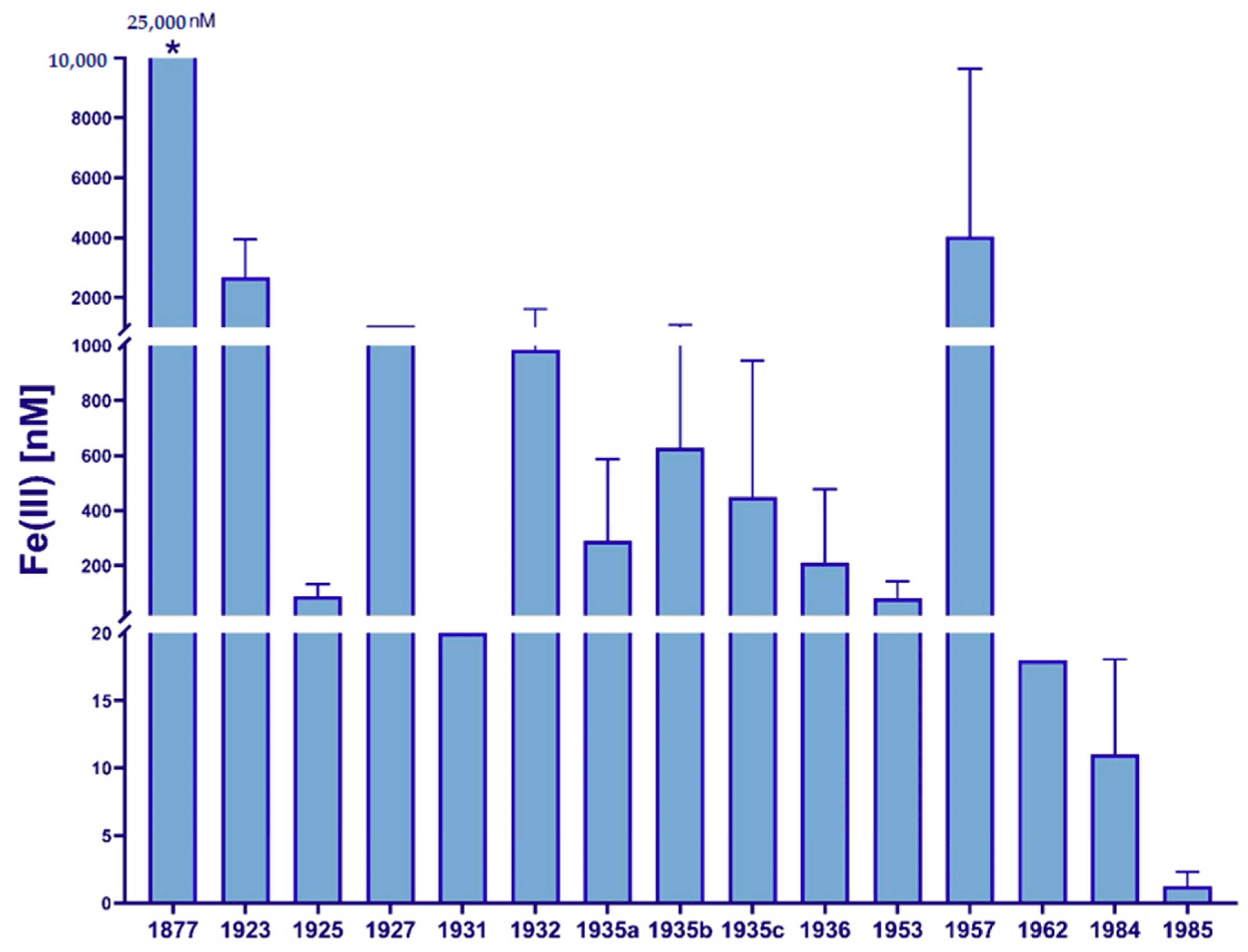
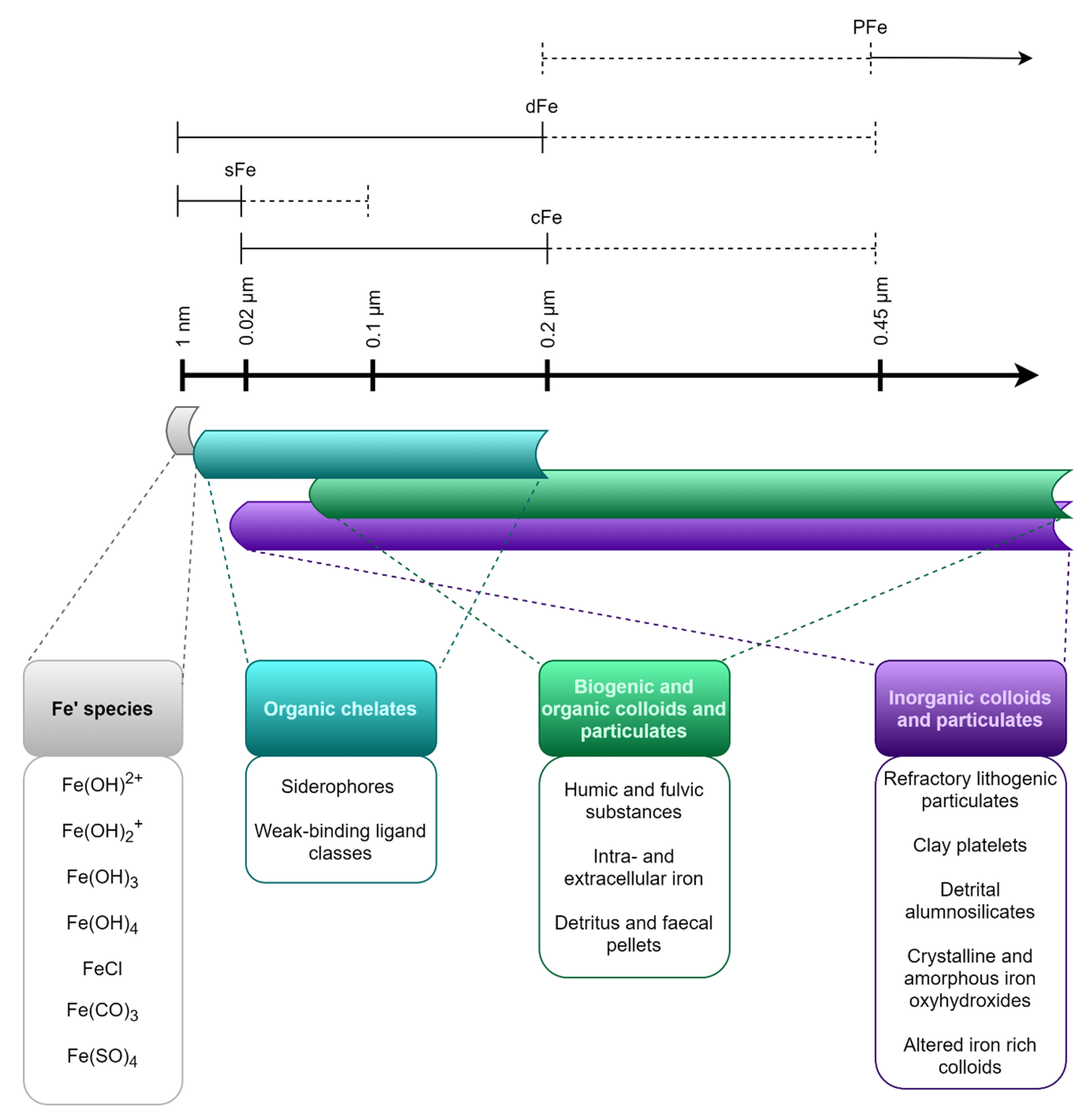
Size Fractions of Iron in Seawater
5. Challenges and Methodical Problems in Iron Limitation Studies
5.1. Physiological Challenges
5.2. Iron Related Challenges
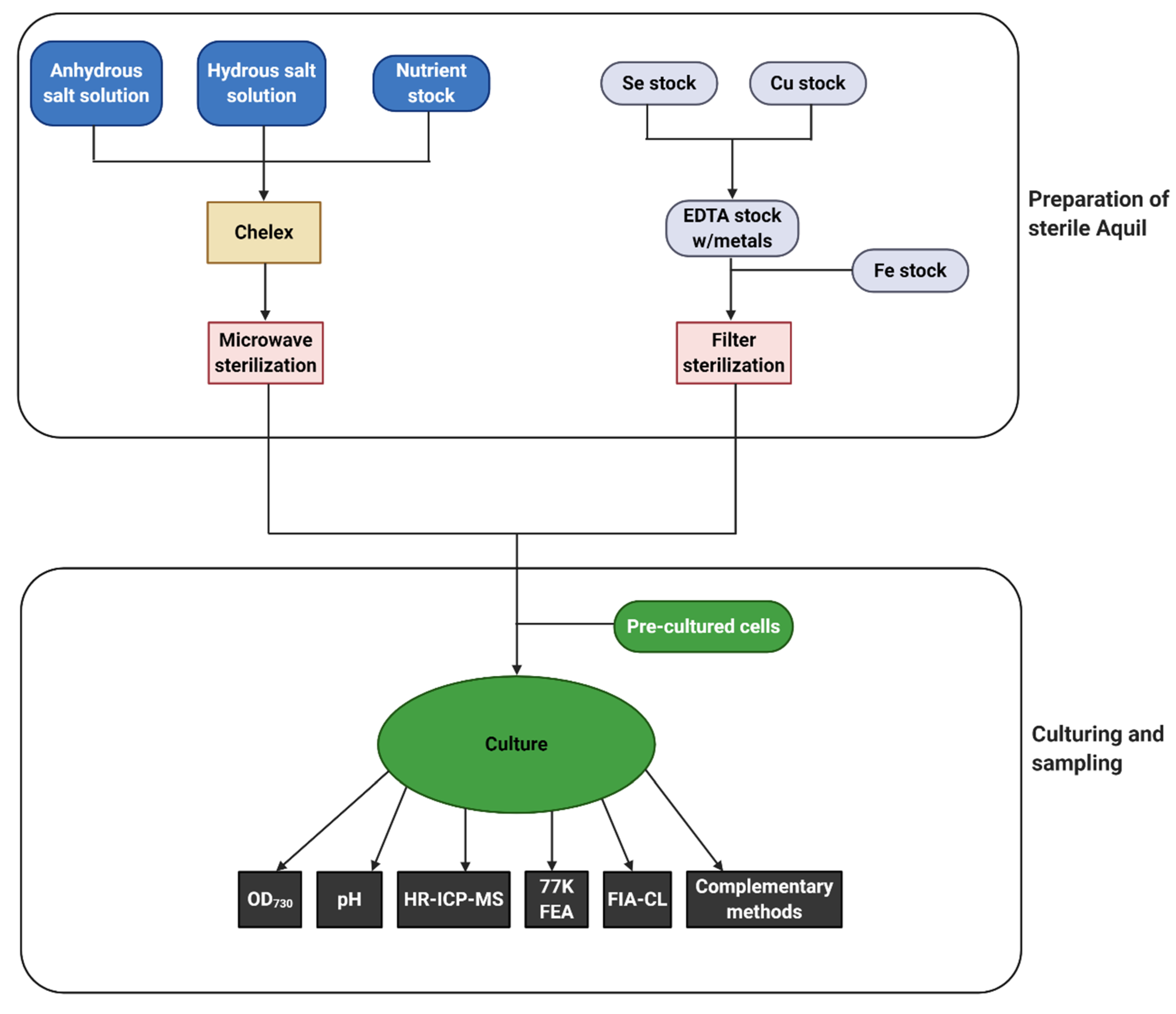
6. Methodology
6.1. Analysis of Fe(III) Fractions by HR-ICP-MS
6.2. Measurement of Fe(II)
6.3. Assessment of Photosynthetic Properties
6.4. Assessment of Cell Number and Features
6.5. Molecular Biological Approaches
6.6. Complementary Methods
6.6.1. Measuring Iron Uptake Rates by Radioactive Iron
6.6.2. Stable Isotopes
6.6.3. Analysis of the Iron Oxidation State and Coordination Environment
7. Future Trends
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Norman, L.; Cabanes, D.J.E.; Blanco-Ameijeiras, S.; Moisset, S.A.M.; Hassler, C.S. Iron biogeochemistry in aquatic systems: from source to bioavailability. Chimia 2014, 68, 764–771. [Google Scholar] [CrossRef] [PubMed]
- Boyd, P.W.; Ellwood, M.J. The biogeochemical cycle of iron in the ocean. Nat. Geosci. 2010, 3, 675–682. [Google Scholar] [CrossRef]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Fenchel, T. Marine plankton food-chains. Annu. Rev. Ecol. Syst. 1988, 19, 19–38. [Google Scholar] [CrossRef]
- Shi, D.L.; Xu, Y.; Hopkinson, B.M.; Morel, F.M.M. Effect of ocean acidification on iron availability to marine phytoplankton. Science 2010, 327, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Breitbarth, E.; Bellerby, R.J.; Neill, C.C.; Ardelan, M.V.; Meyerhofer, M.; Zollner, E.; Croot, P.L.; Riebesell, U. Ocean acidification affects iron speciation during a coastal seawater mesocosm experiment. Biogeosciences 2010, 7, 1065–1073. [Google Scholar] [CrossRef]
- Boyd, P.W. Physiology and iron modulate diverse responses of diatoms to a warming Southern Ocean. Nat. Clim. Chang. 2019, 9, 148–152. [Google Scholar] [CrossRef]
- Andrew, S.M.; Morell, H.T.; Strzepek, R.F.; Boyd, P.W.; Ellwood, M.J. Iron availability influences the tolerance of Southern Ocean phytoplankton to warming and elevated irradiance. Front. Mar. Sci. 2019, 6. [Google Scholar] [CrossRef]
- De Baar, H.J.W.; Boyd, P.W.; Coale, K.H.; Landry, M.R.; Tsuda, A.; Assmy, P.; Bakker, D.C.E.; Bozec, Y.; Barber, R.T.; Brzezinski, M.A.; et al. Synthesis of iron fertilization experiments: From the iron age in the age of enlightenment. J. Geophys. Res. Ocean. 2005, 110. [Google Scholar] [CrossRef]
- Henderson, G.M.; Anderson, R.F.; Adkins, J.; Andersson, P.; Boyle, E.A.; Cutter, G.; de Baar, H.; Eisenhauer, A.; Frank, M.; Francois, R.; et al. GEOTRACES—An international study of the global marine biogeochemical cycles of trace elements and their isotopes. Chem. Erde. Geochem. 2007, 67, 85–131. [Google Scholar] [CrossRef]
- Sohrin, Y.; Bruland, K.W. Global status of trace elements in the ocean. Trac. Trend Anal. Chem. 2011, 30, 1291–1307. [Google Scholar] [CrossRef]
- Anderson, R.F. GEOTRACES: Accelerating research on the marine biogeochemical cycles of trace elements and their isotopes. Annu. Rev. Mar. Sci. 2020, 12, 49–85. [Google Scholar] [CrossRef] [PubMed]
- Pollard, R.T.; Salter, I.; Sanders, R.J.; Lucas, M.I.; Moore, C.M.; Mills, R.A.; Statham, P.J.; Allen, J.T.; Baker, A.R.; Bakker, D.C.E.; et al. Southern Ocean deep-water carbon export enhanced by natural iron fertilization. Nature 2009, 457, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Ameijeiras, S.; Cosio, C.; Hassler, C.S. Long-term acclimation to iron limitation reveals new insights in metabolism regulation of synechococcus sp. PCC7002. Front. Mar. Sci. 2017, 4. [Google Scholar] [CrossRef]
- Blanco-Ameijeiras, S.; Moisset, S.A.M.; Trimborn, S.; Campbell, D.A.; Heiden, J.P.; Hassler, C.S. Elemental stoichiometry and photophysiology regulation of synechococcus sp. PCC7002 under increasing severity of chronic iron limitation. Plant. Cell Physiol. 2018, 59, 1803–1816. [Google Scholar] [CrossRef]
- Thompson, A.W.; Huang, K.; Saito, M.A.; Chisholm, S.W. Transcriptome response of high-and low-light-adapted Prochlorococcus strains to changing iron availability. ISME J. 2011, 5, 1580–1594. [Google Scholar] [CrossRef]
- Kranzler, C.; Lis, H.; Finkel, O.M.; Schmetterer, G.; Shaked, Y.; Keren, N. Coordinated transporter activity shapes high-affinity iron acquisition in cyanobacteria. ISME J. 2014, 8, 409–417. [Google Scholar] [CrossRef]
- Mackey, K.R.M.; Post, A.F.; McIlvin, M.R.; Cutter, G.A.; John, S.G.; Saito, M.A. Divergent responses of Atlantic coastal and oceanic synechococcus to iron limitation. Proc. Natl. Acad. Sci. USA 2015, 112, 9944. [Google Scholar] [CrossRef]
- GEOTRACES. Cruice and methods manual (Cookbook). Available online: https://www.geotraces.org/methods-cookbook/ (accessed on 29 September 2020).
- Achterberg, E.P.; Holland, T.W.; Bowie, A.R.; Fauzi, R.; Mantoura, C.; Worsfold, P.J. Determination of iron in seawater. Anal. Chim. Acta 2001, 442, 1–14. [Google Scholar] [CrossRef]
- Breitbarth, E.; Achterberg, E.P.; Ardelan, M.V.; Baker, A.R.; Bucciarelli, E.; Chever, F.; Croot, P.L.; Duggen, S.; Gledhill, M.; Hassellov, M.; et al. Iron biogeochemistry across marine systems—Progress from the past decade. Biogeosciences 2010, 7, 1075–1097. [Google Scholar] [CrossRef]
- Sunda, W.G.; Price, N.M.; Morel, F.M.M. Trace metal ion buffers and their use in culture studies. In Algal Culturing Techniques; Andersen, R.A., Ed.; Elsevier Academic Press: London, UK, 2005. [Google Scholar]
- Kranzler, C.; Rudolf, M.; Keren, N.; Schleiff, E. Iron in Cyanobacteria. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2013; Volume 65, pp. 57–105. [Google Scholar]
- Gran, H.H. On the conditions for the production of plankton in the sea. Cons. Perm. Internat. l’Explor. Mer. Rapp. Proces-Verb. 1931, 75, 37–46. [Google Scholar]
- Martin, J.H. Glacial-interglacial Co2 change: The iron hypothesis. Paleoceanography 1990, 5, 1–13. [Google Scholar] [CrossRef]
- Patterson, C.C.; Settle, D. The reduction of orders of magnitude frrors in lead analyses of biological materials and natural waters by evaluating and controlling the extent and sources of industrial lead contamination introduced during sample collecting, handling, and analysis. Accuracy Trace Anal. Sampl. Sample Handl. Anal. 1976, 321. [Google Scholar]
- Brand, L.E.; Sunda, W.G.; Guillard, R.R.L. Limitation of marine-phytoplankton reproductive rates by zinc, manganese, and iron. Limnol. Oceanogr. 1983, 28, 1182–1198. [Google Scholar] [CrossRef]
- De Baar, H.J.W. Von Liebig’s law of the minimum and plankton ecology (1899–1991). Prog. Oceanogr. 1994, 33, 347–386. [Google Scholar] [CrossRef]
- Schmidt, C. Title unknown (as cited by Lewis and Goldberg, 1954). Bull. Acad. Sci. St. Petersburg 1874, 9, 175–229. [Google Scholar]
- Schmidt, C. Title unknown (as cited by Lewis and Goldberg, 1954). Bull. Acad. Sci. St. Petersburg 1877, 10, 525–628. [Google Scholar]
- Vernadsky, W. La Matière Vivante et la Chimie de la Mer 1924. Available online: http://vernadsky.fr/documents.html (accessed on 26 July 2020).
- Lewis, G.; Goldberg, E.D. Iron in marine waters. J. Mar. Res. 1954, 13, 183–197. [Google Scholar]
- Orton, J.H. Title unknown (as cited by Thompson and Bremner, 1935). Mar. Biol. Assoc. Ser. 1923, 2, 6. [Google Scholar]
- Harvey, H. Oxidation in sea water. J. Mar. Biol Assoc. UK 1925, 13, 953–969. [Google Scholar] [CrossRef]
- Wattenberg, H. Die deutsche atlantische expedition auf “Meteor” 4. Ber. Zeitschr. Gesell. Erdkd. 1927, 5–6, 308. [Google Scholar]
- Braarud, T.; Klem, A. Hydrographical and chemical investigations in the sea off Möre. Skr. Hvalrådets Univ. Biol. Lab. Oslo 1931, 1, 1–88. [Google Scholar]
- Thompson, T.G.; Bremner, R.W.; Jamieson, I.M. Occurrence and determination of iron in sea water. Ind. Eng. Chem. Anal. Ed. 1932, 4, 288–290. [Google Scholar] [CrossRef]
- Cooper, L.H.N. Iron in the sea and in marine plankton. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1935, 118, 419–438. [Google Scholar]
- Thompson, T.G.; Bremner, R.W. The occurrence of iron in the waters of the north-east Pacific Ocean. ICES J. Mar. Sci. 1935, 10, 39–47. [Google Scholar] [CrossRef]
- Seiwell, G.E. Note on iron analyses of Atlantic coastal waters. Ecology 1935, 16, 663–664. [Google Scholar] [CrossRef]
- Rakestraw, N.W.; Mahncke, H.E.; Beach, E.F. Determination of iron in sea water. Ind. Eng. Chem. Anal. Ed. 1936, 8, 136–138. [Google Scholar] [CrossRef]
- Simons, L.H.; Monaghan, P.H.; Taggart, M.S. Aluminum and iron in atlantic and gulf of mexico surface waters. Anal. Chem. 1953, 25, 989–990. [Google Scholar] [CrossRef]
- Armstrong, F. The iron content of sea water. J. Mar. Biol Assoc. UK 1957, 36, 509–517. [Google Scholar] [CrossRef]
- Menzel, D.W.; Spaeth, J.P. Occurrence of iron in the Sargasso Sea off Bermuda. Limnol. Oceanogr. 1962, 7, 155–158. [Google Scholar] [CrossRef]
- Rao, D.S.; Yeats, P. Effect of iron on phytoplankton production in the Sargasso Sea. J. Exp. Mar. Biol Ecol. 1984, 81, 281–289. [Google Scholar] [CrossRef]
- Symes, J.L.; Kester, D.R. The distribution of iron in the northwest Atlantic. Mar. Chem. 1985, 17, 57–74. [Google Scholar] [CrossRef]
- Martin, J.H.; Fitzwater, S.E. Iron-deficiency limits phytoplankton growth in the Northeast Pacific Subarctic. Nature 1988, 331, 341–343. [Google Scholar] [CrossRef]
- Bruland, K.W.; Donat, J.R.; Hutchins, D.A. Interactive influences of bioactive trace metals on biological production in oceanic waters. Limnol. Oceanogr. 1991, 36, 1555–1577. [Google Scholar] [CrossRef]
- Hudson, R.J.M.; Morel, F.M.M. lron transport in marine phytoplankton: Kinetics of cellular and medium coordination reactions. Limnol. Oceanogr. 1990, 35, 1002–1020. [Google Scholar] [CrossRef]
- Martin, J.H.; Gordon, R.M. Northeast Pacific iron distributions in relation to phytoplankton productivity. Deep Sea Res. 1988, 35, 177–196. [Google Scholar] [CrossRef]
- Martin, J.H.; Coale, K.H.; Johnson, K.S.; Fitzwater, S.E.; Gordon, R.M.; Tanner, S.J.; Hunter, C.N.; Elrod, V.A.; Nowicki, J.L.; Coley, T.L.; et al. Testing the iron hypothesis in ecosystems of the equatorial Pacific-Ocean. Nature 1994, 371, 123–129. [Google Scholar] [CrossRef]
- Coale, K.H.; Johnson, K.S.; Fitzwater, S.E.; Blain, S.P.G.; Stanton, T.P.; Coley, T.L. IronEx-I, an in situ iron-enrichment experiment: Experimental design, implementation and results. Deep Sea Res. Pt. II 1998, 45, 919–945. [Google Scholar] [CrossRef]
- Bakker, D.C.E.; Bozec, Y.; Nightingale, P.D.; Goldson, L.; Messias, M.J.; de Baar, H.J.W.; Liddicoat, M.; Skjelvan, I.; Strass, V.; Watson, A.J. Iron and mixing affect biological carbon uptake in SOIREE and EisenEx, two Southern Ocean iron fertilisation experiments. Deep Sea Res. Pt. I 2005, 52, 1001–1019. [Google Scholar] [CrossRef][Green Version]
- Tagliabue, A.; Bowie, A.R.; Boyd, P.W.; Buck, K.N.; Johnson, K.S.; Saito, M.A. The integral role of iron in ocean biogeochemistry. Nature 2017, 543, 51–59. [Google Scholar] [CrossRef]
- Hernandez, J.A.; Curatti, L.; Aznar, C.P.; Perova, Z.; Britt, R.D.; Rubio, L.M. Metal trafficking for nitrogen fixation: NifQ donates molybdenum to NifEN/NifH for the biosynthesis of the nitrogenase FeMo-cofactor. Proc. Natl. Acad. Sci. USA 2008, 105, 11679–11684. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A. The iron and molybdenum use efficiencies of plant-growth with different energy, carbon and nitrogen-sources. New Phytol. 1988, 109, 279–287. [Google Scholar] [CrossRef]
- González Rodríguez, A.; Fillat, M.; Bes, M.-T.; Peleato, M.-L.; Sevilla, E. The challenge of iron stress. In Cyanobacteria; Books on Demand: Norderstedt, Germany, 2018. [Google Scholar] [CrossRef]
- Ivanov, A.G.; Krol, M.; Sveshnikov, D.; Selstam, E.; Sandström, S.; Koochek, M.; Park, Y.-I.; Vasil’ev, S.; Bruce, D.; Oquist, G.; et al. Iron deficiency in cyanobacteria causes monomerization of photosystem I trimers and reduces the capacity for state transitions and the effective absorption cross section of photosystem I in vivo. Plant. Physiol. 2006, 141, 1436–1445. [Google Scholar] [CrossRef] [PubMed]
- Boyd, P.W.; Crossley, A.C.; DiTullio, G.R.; Griffiths, F.B.; Hutchins, D.A.; Queguiner, B.; Sedwick, P.N.; Trull, T.W. Control of phytoplankton growth by iron supply and irradiance in the subantarctic Southern Ocean: Experimental results from the SAZ Project. J. Geophys. Res. Ocean. 2001, 106, 31573–31583. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Barber, R.T.; Smetacek, V. Biogeochemical controls and feedbacks on ocean primary production. Science 1998, 281, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Sarthou, G.; Baker, A.R.; Blain, S.; Achterberg, E.P.; Boye, M.; Bowie, A.R.; Croot, P.; Laan, P.; de Baar, H.J.W.; Jickells, T.D.; et al. Atmospheric iron deposition and sea-surface dissolved iron concentrations in the eastern Atlantic Ocean. Deep Sea Res. Pt. I 2003, 50, 1339–1352. [Google Scholar] [CrossRef][Green Version]
- Yucel, M.; Gartman, A.; Chan, C.S.; Luther, G.W. Hydrothermal vents as a kinetically stable source of iron-sulphide-bearing nanoparticles to the ocean. Nat. Geosci. 2011, 4, 367–371. [Google Scholar] [CrossRef]
- Ardelan, M.V.; Holm-Hansen, O.; Hewes, C.D.; Reiss, C.S.; Silva, N.S.; Dulaiova, H.; Steinnes, E.; Sakshaug, E. Natural iron enrichment around the Antarctic Peninsula in the Southern Ocean. Biogeosciences 2010, 7, 11–25. [Google Scholar] [CrossRef]
- Laufkotter, C.; Stern, A.A.; John, J.G.; Stock, C.A.; Dunne, J.P. Glacial iron sources stimulate the Southern Ocean carbon cycle. Geophys. Res. Lett. 2018, 45, 13377–13385. [Google Scholar] [CrossRef]
- Hawkings, J.R.; Benning, L.G.; Raiswell, R.; Kaulich, B.; Araki, T.; Abyaneh, M.; Stockdale, A.; Koch-Muller, M.; Wadham, J.L.; Tranter, M. Biolabile ferrous iron bearing nanoparticles in glacial sediments. Earth Planet. Sci. Lett. 2018, 493, 92–101. [Google Scholar] [CrossRef]
- Krachler, R.; Krachler, R.; Valda, A.; Keppler, B.K. Natural iron fertilization of the coastal ocean by “blackwater rivers”. Sci. Total Env. 2019, 656, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Dulaiova, H.; Ardelan, M.V.; Henderson, P.B.; Charette, M.A. Shelf-derived iron inputs drive biological productivity in the southern Drake Passage. Glob. Biogeochem. Cycles 2009, 23. [Google Scholar] [CrossRef]
- Sanchez, N.; Reiss, C.S.; Holm-Hansen, O.; Hewes, C.D.; Bizsel, K.C.; Ardelan, M.V. Weddell-Scotia confluence effect on the iron distribution in waters surrounding the South Shetland (Antarctic Peninsula) and South Orkney (Scotia Sea) islands during the austral summer in 2007 and 2008. Front. Mar. Sci. 2019, 6. [Google Scholar] [CrossRef]
- Rijkenberg, M.J.A.; Slagter, H.A.; van der Loeff, M.R.; van Ootien, J.; Gerringa, L.J.A. Dissolved Fe in the deep and upper arctic ocean with a focus on Fe limitation in the Nansen basin. Front. Mar. Sci. 2018, 5. [Google Scholar] [CrossRef]
- Mahowald, N.M.; Baker, A.R.; Bergametti, G.; Brooks, N.; Duce, R.A.; Jickells, T.D.; Kubilay, N.; Prospero, J.M.; Tegen, I. Atmospheric global dust cycle and iron inputs to the ocean. Glob. Biogeochem. Cycles 2005, 19. [Google Scholar] [CrossRef]
- Zhao, C.M.; Campbell, P.G.C.; Wilkinson, K.J. When are metal complexes bioavailable? Env. Chem. 2016, 13, 425–433. [Google Scholar] [CrossRef]
- Wells, M.L.; Trick, C.G. Controlling iron availability to phytoplankton in iron-replete coastal waters. Mar. Chem. 2004, 86, 1–13. [Google Scholar] [CrossRef]
- Gledhill, M.; Van Den Berg, C.M.G. Determination of complexation of iron (III) with natural organic complexing ligands in seawater using cathodic stripping voltammetry. Mar. Chem. 1994, 47, 41–54. [Google Scholar] [CrossRef]
- Rue, E.L.; Bruland, K.W. Complexation of iron (III) by natural organic-ligands in the central north pacific as determined by a new competitive ligand equilibration adsorptive cathodic stripping voltammetric method. Mar. Chem. 1995, 50, 117–138. [Google Scholar] [CrossRef]
- Wu, J.F.; Boyle, E.; Sunda, W.; Wen, L.S. Soluble and colloidal iron in the olgotrophic North Atlantic and North Pacific. Science 2001, 293, 847–849. [Google Scholar] [CrossRef]
- Ozturk, M.; Bizsel, N. Iron speciation and biogeochemistry in different nearshore waters. Mar. Chem. 2003, 83, 145–156. [Google Scholar] [CrossRef]
- Wilhelm, S.W.; Trick, C.G. Iron-limited growth of cyanobacteria—Multiple siderophore production is a common response. Limnol. Oceanogr. 1994, 39, 1979–1984. [Google Scholar] [CrossRef]
- Gledhill, M.; Buck, K.N. The organic complexation of iron in the marine environment: A review. Front. Microbiol 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Rich, H.W.; Morel, F.M.M. Availability of well-defined iron colloids to the marine diatom thalassiosira-weissflogii. Limnol. Oceanogr. 1990, 35, 652–662. [Google Scholar] [CrossRef]
- Wells, M.L.; Price, N.M.; Bruland, K.W. Iron chemistry in seawater and its relationship to phytoplankton—A workshop report. Mar. Chem. 1995, 48, 157–182. [Google Scholar] [CrossRef]
- Hopwood, M.J.; Birchill, A.J.; Gledhill, M.; Achterberg, E.P.; Klar, J.K.; Milne, A. A comparison between four analytical methods for the measurement of Fe (II) at nanomolar concentrations in coastal seawater. Front. Mar. Sci. 2017, 4. [Google Scholar] [CrossRef]
- Lis, H.; Kranzler, C.; Keren, N.; Shaked, Y. A comparative study of iron uptake rates and mechanisms amongst marine and fresh water cyanobacteria: Prevalence of reductive iron uptake. Life 2015, 5, 841–860. [Google Scholar] [CrossRef]
- Srai, S.K.S.; Bomford, A.; McArdle, H.J. Iron transport across cell membranes: Molecular understanding of duodenal and placental iron uptake. Best Pract. Res. Clin. Haematol. 2002, 15, 243–259. [Google Scholar] [CrossRef]
- Sunda, W.G. Bioavailability and bioaccumulation of iron in the sea. In The Biogeochemistry of Iron in Seawater; Turner, D.R., Hunter, K.A., Eds.; Wiley: New York, NY, USA, 2001; pp. 41–48. [Google Scholar]
- Millero, F.J.; Sotolongo, S.; Izaguirre, M. The oxidation-kinetics of Fe (II) in seawater. Geochim. Cosmochim. Acta 1987, 51, 793–801. [Google Scholar] [CrossRef]
- Santana-Casiano, J.M.; Gonzalez-Davila, A.; Millero, F.J. The role of Fe (II) species on the oxidation of Fe (II) in natural waters in the presence of O-2 and H2O2. Mar. Chem. 2006, 99, 70–82. [Google Scholar] [CrossRef]
- Flombaum, P.; Gallegos, J.L.; Gordillo, R.A.; Rincon, J.; Zabala, L.L.; Jiao, N.A.Z.; Karl, D.M.; Li, W.K.W.; Lomas, M.W.; Veneziano, D.; et al. Present and future global distributions of the marine cyanobacteria prochlorococcus and synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 9824–9829. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, F.; Straus, N.A. Iron deprivation in cyanobacteria. J. Appl. Phycol. 1994, 6, 199–210. [Google Scholar] [CrossRef]
- Singh, A.K.; McIntyre, L.M.; Sherman, L.A. Microarray analysis of the genome-wide response to iron deficiency and iron reconstitution in the cyanobacterium synechocystis sp PCC 6803. Plant. Physiol. 2003, 132, 1825–1839. [Google Scholar] [CrossRef] [PubMed]
- Bibby, T.S.; Nield, J.; Barber, J. Three-dimensional model and characterization of the iron stress-induced CP43 ′-photosystem I supercomplex isolated from the cyanobacterium synechocystis PCC 6803. J. Biol. Chem. 2001, 276, 43246–43252. [Google Scholar] [CrossRef] [PubMed]
- Sandmann, G. Consequences of iron-deficiency on photosynthetic and respiratory electron-transport in blue-green-algae. Photosynth. Res. 1985, 6, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Shcolnick, S.; Summerfield, T.C.; Reytman, L.; Sherman, L.A.; Keren, N. The mechanism of iron homeostasis in the unicellular cyanobacterium synechocystis sp PCC 6803 and its relationship to oxidative stress. Plant. Physiol. 2009, 150, 2045–2056. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, B.R.; John, S.G. The effect of iron limitation on cyanobacteria major nutrient and trace element stoichiometry. Limnol. Oceanogr. 2017, 62, 846–858. [Google Scholar] [CrossRef]
- Guikema, J.A.; Sherman, L.A. Organization and function of chlorophyll in membranes of cyanobacteria during iron starvation. Plant. Physiol. 1983, 73, 250–256. [Google Scholar] [CrossRef]
- Bibby, T.S.; Nield, J.; Barber, J. Iron deficiency induces the formation of an antenna ring around trimeric photosystem I in cyanobacteria. Nature 2001, 412, 743–745. [Google Scholar] [CrossRef]
- Katoh, H.; Hagino, N.; Grossman, A.R.; Ogawa, T. Genes essential to iron transport in the cyanobacterium synechocystis sp strain PCC 6803. J. Bacteriol. 2001, 183, 2779–2784. [Google Scholar] [CrossRef]
- Kranzler, C.; Lis, H.; Shaked, Y.; Keren, N. The role of reduction in iron uptake processes in a unicellular, planktonic cyanobacterium. Env. Microbiol. 2011, 13, 2990–2999. [Google Scholar] [CrossRef] [PubMed]
- Årstøl, E.; Hohmann-Marriott, F.M. Cyanobacterial siderophores—Physiology, structure, biosynthesis, and applications. Mar. Drugs 2019, 17, 281. [Google Scholar] [CrossRef]
- Lamb, J.J.; Hill, R.E.; Eaton-Rye, J.J.; Hohmann-Marriott, M.F. Functional role of PilA in iron acquisition in the cyanobacterium synechocystis sp PCC 6803. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Lamb, J.J.; Hohmann-Marriott, M.F. Manganese acquisition is facilitated by PilA in the cyanobacterium synechocystis sp PCC 6803. PLoS ONE 2017, 12. [Google Scholar] [CrossRef]
- Pennington, J.T.; Blum, M.; Chavez, F.P. Seawater sampling by an autonomous underwater vehicle: “Gulper” sample validation for nitrate, chlorophyll, phytoplankton, and primary production. Limnol. Oceanogr. Methods 2016, 14, 14–23. [Google Scholar] [CrossRef]
- Geißler, F.; Achterberg, E.P.; Beaton, A.D.; Hopwood, M.J.; Clarke, J.S.; Mutzberg, A.; Mowlem, M.C.; Connelly, D.P. Evaluation of a ferrozine based autonomous in situ lab-on-chip analyzer for dissolved iron species in coastal waters. Front. Mar. Sci. 2017, 4. [Google Scholar] [CrossRef]
- Kim, H.; Bishop, J.K.; Wood, T.J.; Fung, I.Y. Autonomous water sampling for long-term monitoring of trace metals in remote environments. Environ. Sci. Technol. 2012, 46, 11220–11226. [Google Scholar] [CrossRef]
- Planquette, H.; Sherrell, R.M. Sampling for particulate trace element determination using water sampling bottles: Methodology and comparison to in situ pumps. Limnol. Oceanogr. Methods 2012, 10, 367–388. [Google Scholar] [CrossRef]
- Measures, C.I.; Landing, W.; Brown, M.T.; Buck, C. A commercially available rosette system for trace metal–Clean sampling. Limnol. Oceanogr. Methods 2008, 6, 384–394. [Google Scholar] [CrossRef]
- Sakshaug, E.; Johnsen, G.H.; Kovacs, K.M. Ecosystem Barents Sea; Tapir Academic: Trondheim, Norway, 2009; 587p. [Google Scholar]
- Clarke, G.L.; Ewing, G.C.; Lorenzen, C.J. Spectra of backscattered light from the sea obtained from aircraft as a measure of chlorophyll concentration. Science 1970, 167, 1119. [Google Scholar] [CrossRef]
- Roesler, C.S.; Barnard, A.H. Optical proxy for phytoplankton biomass in the absence of photophysiology: Rethinking the absorption line height. Methods Oceanogr. 2013, 7, 79–94. [Google Scholar] [CrossRef]
- Descy, J.P.; Sarmento, H.; Higgins, H.W. Variability of phytoplankton pigment ratios across aquatic environments. Eur. J. Phycol. 2009, 44, 319–330. [Google Scholar] [CrossRef]
- Aminot, A.; Rey, F. Standard Procedure for the Determination of Chlorophyll a by Spectroscopic Methods; International Council for the Exploration of the Sea: Copenhagen, Denmark, 2002; Volume 28. [Google Scholar]
- Babin, M. Phytoplankton fluorescence: Theory, current literature and in situ measurement. In Real-Time Coastal Observing Systems for Marine Ecosystem Dynamics and Harmful Algal Blooms; Babin, M., Roesler, C., Cullen, J.J., Eds.; UNESCO: Paris, France, 2008; pp. 237–280. [Google Scholar]
- Boyd, P.W.; Collins, S.; Dupont, S.; Fabricius, K.; Gattuso, J.P.; Havenhand, J.; Hutchins, D.A.; Riebesell, U.; Rintoul, M.S.; Vichi, M. Experimental strategies to assess the biological ramifications of multiple drivers of global ocean change—A review. Glob. Chang. Biol. 2018, 24, 2239–2261. [Google Scholar] [CrossRef] [PubMed]
- Morel, F.M.M.; Rueter, J.G.; Anderson, D.M.; Guillard, R.R.L. Aquil—Chemically defined phytoplankton culture-medium for trace-metal studies. J. Phycol. 1979, 15, 135–141. [Google Scholar] [CrossRef]
- Von der Heyden, B.P.; Roychoudhury, A.N. A review of colloidal iron partitioning and distribution in the open ocean. Mar. Chem. 2015, 177, 9–19. [Google Scholar] [CrossRef]
- Bruland, K.W.; Rue, E.L. Iron: Analytical methods for the determination of concentrations and speciation. In The Biogeochemistry of Iron in Seawater; J. Wiley: Hoboken, NJ, USA, 2001; pp. 255–289. [Google Scholar]
- Croot, P.L.; Streu, P.; Baker, A.R. Short residence time for iron in surface seawater impacted by atmospheric dry deposition from Saharan dust events. Geophys. Res. Lett. 2004, 31. [Google Scholar] [CrossRef]
- Frew, R.D.; Hutchins, D.A.; Nodder, S.; Sanudo-Wilhelmy, S.; Tovar-Sanchez, A.; Leblanc, K.; Hare, C.E.; Boyd, P.W. Particulate iron dynamics during FeCycle in subantarctic waters southeast of New Zealand. Glob. Biogeochem. Cycles 2006, 20. [Google Scholar] [CrossRef]
- Johnson, K.S.; Gordon, R.M.; Coale, K.H. What controls dissolved iron concentrations in the world ocean? Mar. Chem. 1997, 57, 137–161. [Google Scholar] [CrossRef]
- Tovar-Sanchez, A.; Sanudo-Wilhelmy, S.A.; Garcia-Vargas, M.; Weaver, R.S.; Popels, L.C.; Hutchins, D.A. A trace metal clean reagent to remove surface-bound iron from marine phytoplankton. Mar. Chem. 2003, 82, 91–99. [Google Scholar] [CrossRef]
- Tang, D.G.; Morel, F.M.M. Distinguishing between cellular and Fe-oxide-associated trace elements in phytoplankton. Mar. Chem. 2006, 98, 18–30. [Google Scholar] [CrossRef]
- Hopwood, M.J.; Santana-Gonzáez, C.; Gallego-Urrea, J.; Sanchez, N.; Achterberg, E.P.; Ardelan, M.V.; Gledhill, M.; González-Dávila, M.; Hoffmann, L.; Leiknes, Ø.; et al. Fe(II) stability in seawater. Biogeosci. Discuss. 2018, 2018, 1–29. [Google Scholar] [CrossRef]
- Lueder, U.; Jorgensen, B.B.; Kappler, A.; Schmidt, C. Photochemistry of iron in aquatic environments. Env. Sci. Process. Impacts 2020, 22, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Jezberová, J.; Komárková, J. Morphometry and growth of three synechococcus-like picoplanktic cyanobacteria at different culture conditions. Hydrobiologia 2007, 578, 17–27. [Google Scholar] [CrossRef]
- Moore, L.R.; Coe, A.; Zinser, E.R.; Saito, M.A.; Sullivan, M.B.; Lindell, D.; Frois-Moniz, K.; Waterbury, J.; Chisholm, S.W. Culturing the marine cyanobacterium Prochlorococcus. Limnol. Oceanogr. Methods 2007, 5, 353–362. [Google Scholar] [CrossRef]
- Kaneko, T.; Sato, S.; Kotani, H.; Tanaka, A.; Asamizu, E.; Nakamura, Y.; Miyajima, N.; Hirosawa, M.; Sugiura, M.; Sasamoto, S.; et al. Sequence analysis of the genome of the unicellular cyanobacterium synechocystis sp. strain PCC6803. II. Sequence determination of the entire genome and assignment of potential protein-coding regions. DNA Res. 1996, 3, 109–136. [Google Scholar] [CrossRef]
- Kaneko, T.; Tanaka, A.; Sato, S.; Kotani, H.; Sazuka, T.; Miyajima, N.; Sugiura, M.; Tabata, S. Sequence analysis of the genome of the unicellular cyanobacterium synechocystis sp. strain PCC6803. I. Sequence features in the 1 Mb region from map positions 64% to 92% of the genome. DNA Res. 1995, 2, 153–166. [Google Scholar] [CrossRef]
- Morris, J.N.; Crawford, T.S.; Jeffs, A.; Stockwell, P.A.; Eaton-Rye, J.J.; Summerfield, T.C. Whole genome re-sequencing of two ‘wild-type’ strains of the model cyanobacterium synechocystis sp. PCC 6803. N. Zeal. J. Bot. 2014, 52, 36–47. [Google Scholar] [CrossRef]
- Allen, R.C.; Rittmann, B.E.; Curtiss, R. Axenic biofilm formation and aggregation by synechocystis sp strain pcc 6803 is induced by changes in nutrient concentration, and requires cell surface structures. bioRxiv 2018. [Google Scholar] [CrossRef]
- Keller, M.D.; Bellows, W.K.; Guillard, R.R.L. Microwave treatment for sterilization of phytoplankton culture media. J. Exp. Mar. Biol. Ecol. 1988, 117, 279–283. [Google Scholar] [CrossRef]
- Sarthou, G.; Bucciarelli, E.; Chever, F.; Hansard, S.P.; Gonzalez-Davila, M.; Santana-Casiano, J.M.; Planchon, F.; Speich, S. Labile Fe(II) concentrations in the Atlantic sector of the Southern Ocean along a transect from the subtropical domain to the Weddell Sea Gyre. Biogeosciences 2011, 8, 2461–2479. [Google Scholar] [CrossRef]
- Croot, P.L.; Laan, P. Continuous shipboard determination of Fe (II) in polar waters using flow injection analysis with chemiluminescence detection. Anal. Chim. Acta 2002, 466, 261–273. [Google Scholar] [CrossRef]
- González-Davila, M.; Santana-Casiano, J.M.; Millero, F.J. Oxidation of nanomolar levels of Fe(ii) with oxygen in natural waters. Geochim. Cosmochim. Acta 2005, 69, 83–93. [Google Scholar] [CrossRef]
- Morgan, B.; Lahav, O. The effect of pH on the kinetics of spontaneous Fe (II) oxidation by O2 in aqueous solution—Basic principles and a simple heuristic description. Chemosphere 2007, 68, 2080–2084. [Google Scholar] [CrossRef] [PubMed]
- Emmenegger, L.; King, D.W.; Sigg, L.; Sulzberger, B. Oxidation kinetics of Fe (II) in a Eutrophic Swiss Lake. Env. Sci. Technol. 1998, 32, 2990–2996. [Google Scholar] [CrossRef]
- Tagliabue, A.; Voelker, C. Towards accounting for dissolved iron speciation in global ocean models. Biogeosciences 2011, 8, 3025–3039. [Google Scholar] [CrossRef]
- Fan, S.-M. Photochemical and biochemical controls on reactive oxygen and iron speciation in the pelagic surface ocean. Mar. Chem. 2008, 109, 152–164. [Google Scholar] [CrossRef]
- Kleint, C.; Hawkes, J.A.; Sander, S.G.; Koschinsky, A. Voltammetric investigation of hydrothermal iron speciation. Front. Mar. Sci. 2016, 3. [Google Scholar] [CrossRef]
- Caprara, S.; Buck, K.N.; Gerringa, L.J.A.; Rijkenberg, M.J.A.; Monticelli, D. A compilation of iron speciation data for open oceanic waters. Front. Mar. Sci. 2016, 3. [Google Scholar] [CrossRef]
- Hutchins, D.A.; Witter, A.E.; Butler, A.; Luther, G.W. Competition among marine phytoplankton for different chelated iron species. Nature 1999, 400, 858–861. [Google Scholar] [CrossRef]
- Hassler, C.S.; Alasonati, E.; Mancuso Nichols, C.A.; Slaveykova, V.I. Exopolysaccharides produced by bacteria isolated from the pelagic Southern Ocean—Role in Fe binding, chemical reactivity, and bioavailability. Mar. Chem. 2011, 123, 88–98. [Google Scholar] [CrossRef]
- National Research Council Canada. Certified Reference Materials. National Research Council Canada; 2019. Available online: https://nrc.canada.ca/en/certifications-evaluations-standards/certified-reference-materials (accessed on 10 November 2020).
- Milne, A.; Landing, W.; Bizimis, M.; Morton, P. Determination of Mn, Fe, Co, Ni, Cu, Zn, Cd and Pb in seawater using high resolution magnetic sector inductively coupled mass spectrometry (HR-ICP-MS). Anal. Chim. Acta 2010, 665, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.A.; Noble, A.E.; Tagliabue, A.; Goepfert, T.J.; Lamborg, C.H.; Jenkins, W.J. Slow-spreading submarine ridges in the South Atlantic as a significant oceanic iron source. Nat. Geosci. 2013, 6, 775–779. [Google Scholar] [CrossRef]
- Vegueria, S.F.J.; Godoy, J.M.; de Campos, R.C.; Goncalves, R.A. Trace element determination in seawater by ICP-MS using online, offline and bath procedures of preconcentration and matrix elimination. Microchem. J. 2013, 106, 121–128. [Google Scholar] [CrossRef]
- Sondergaard, J.; Asmund, G.; Larsen, M.M. Trace elements determination in seawater by ICP-MS with on-line pre-concentration on a Chelex-100 column using a ‘standard’ instrument setup. Methodsx 2015, 2, 323–330. [Google Scholar] [CrossRef]
- Kuma, K.; Nishioka, J.; Matsunaga, K. Controls on iron (III) hydroxide solubility in seawater: The influence of pH and natural organic chelators. Limnol. Oceanogr. 1996, 41, 396–407. [Google Scholar] [CrossRef]
- Wurl, O. Practical Guidelines for the Analysis of Seawater; CRC Press: Boca Raton, FL, USA, 2009; 401p. [Google Scholar]
- Ozturk, M.; Steinnes, E.; Sakshaug, E. Iron speciation in the Trondheim fjord from the perspective of iron limitation for phytoplankton. Estuar. Coast. Shelf. Sci. 2002, 55, 197–212. [Google Scholar] [CrossRef]
- Calatayud, J.M.N. Flow Injection Analysis of Pharmaceuticals: Automation in the Laboratory; Taylor & Francis: London, UK; Bristol, UK, 1996; 394p. [Google Scholar]
- Decastro, M.D.L.; Valcarcel, M. Novel kinetic approaches to flow-injection analysis. Trac Trend Anal. Chem. 1989, 8, 172–177. [Google Scholar]
- Vedamati, J.; Goepfert, T.; Moffett, J.W. Iron speciation in the eastern tropical South Pacific oxygen minimum zone off Peru. Limnol. Oceanogr. 2014, 59, 1945–1957. [Google Scholar] [CrossRef]
- King, D.W.; Lounsbury, H.A.; Millero, F.J. Rates and mechanism of Fe (II) oxidation at nanomolar total iron concentrations. Env. Sci. Technol. 1995, 29, 818–824. [Google Scholar] [CrossRef]
- Johnson, K.S.; Elrod, V.; Fitzwater, S.; Plant, J.; Boyle, E.; Bergquist, B.; Bruland, K.; Aguilar-Islas, A.; Buck, K.; Lohan, M.; et al. Developing standards for dissolved iron in seawater. EosTrans. Am. Geophys. Union 2007, 88, 131–132. [Google Scholar] [CrossRef]
- Measures, C.I.; Yuan, J.; Resing, J.A. Determination of iron in seawater by flow injection analysis using in-line preconcentration and spectrophotometric detection. Mar. Chem. 1995, 50, 3–12. [Google Scholar] [CrossRef]
- Bowie, A.R.; Achterberg, E.P.; Sedwick, P.N.; Ussher, S.; Worsfold, P.J. Real-time monitoring of picomolar concentrations of iron (II) in marine waters using automated flow injection-chemiluminescence instrumentation. Env. Sci. Technol. 2002, 36, 4600–4607. [Google Scholar] [CrossRef] [PubMed]
- Bowie, A.R.; Sedwick, P.N.; Worsfold, P.J. Analytical intercomparison between flow injection-chemiluminescence and flow injection-spectrophotometry for the determination of picomolar concentrations of iron in seawater. Limnol. Oceanogr. Methods 2004, 2, 42–54. [Google Scholar] [CrossRef]
- Floor, G.H.; Clough, R.; Lohan, M.C.; Ussher, S.J.; Worsfold, P.J.; Quétel, C.R. Combined uncertainty estimation for the determination of the dissolved iron amount content in seawater using flow injection with chemiluminescence detection. Limnol. Oceanogr. Methods 2015, 13, 673–686. [Google Scholar] [CrossRef] [PubMed]
- Miró, M.; Estela, J.M.; Cerdà, V. Potentials of multisyringe flow injection analysis for chemiluminescence detection. Anal. Chim. Acta 2005, 541, 55–66. [Google Scholar] [CrossRef]
- Oliveira, H.M.; Grand, M.M.; Ruzicka, J.; Measures, C.I. Towards chemiluminescence detection in micro-sequential injection lab-on-valve format: A proof of concept based on the reaction between Fe (II) and luminol in seawater. Talanta 2015, 133, 107–111. [Google Scholar] [CrossRef]
- Hirata, S.; Yoshihara, H.; Aihara, M. Determination of iron (II) and total iron in environmental water samples by flow injection analysis with column preconcentration of chelating resin functionalized with N-hydroxyethylethylenediamine ligands and chemiluminescence detection. Talanta 1999, 49, 1059–1067. [Google Scholar] [CrossRef]
- King, D.W.; Lin, J.; Kester, D.R. Spectrophotometric determination of iron (II) in seawater at nanomolar concentrations. Anal. Chim. Acta 1991, 247, 125–132. [Google Scholar] [CrossRef]
- Blain, S.; Tréguer, P. Ironn (II) and iron (III) determination in sea water at the nanomolar level with selective on-line preconcentration and spectrophotometric determination. Anal. Chim. Acta 1995, 308, 425–432. [Google Scholar] [CrossRef]
- Stookey, L.L. Ferrozine—A new spectrophotometric reagent for iron. Anal. Chem. 1970, 42, 779–781. [Google Scholar] [CrossRef]
- Milani, A.; Floquet, C.; Connelly, D.; Mowlem, M.; Statham, P. Microfluidic technology for in-situ detection of iron at low concentration in seawater. In Proceedings of the EGU General Assembly 2012, Vienna, Austria, 22–27 April 2012; p. 4564. [Google Scholar]
- Milani, A.; Statham, P.J.; Mowlem, M.C.; Connelly, D.P. Development and application of a microfluidic in-situ analyzer for dissolved Fe and Mn in natural waters. Talanta 2015, 136, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Jaywant, S.A.; Arif, K.M. A comprehensive review of microfluidic water quality monitoring sensors. Sensors 2019, 19, 4781. [Google Scholar] [CrossRef] [PubMed]
- Huot, Y.; Babin, M. Overview of fluorescence protocols: theory, basic concepts, and practice. In Chlorophyll a Fluorescence in Aquatic Sciences. Methods and Applications; Springer: Dordrecht, The Netherlands, 2010; p. 334. [Google Scholar] [CrossRef]
- Gutiérrez-Rodríguez, A.; Latasa, M. Pigment-based measurements of phytoplankton rates. In Phytoplankton Pigments: Characterization, Chemotaxonomy and Applications in Oceanography; Llewellyn, C.A., Egeland, E.S., Johnsen, G., Roy, S., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 472–495. [Google Scholar] [CrossRef]
- Roy, S. Phytoplankton Pigments: Characterization, Chemotaxonomy, and Applications in Oceanography; Cambridge University Press: Cambridge, UK, 2011; pp. 163–164. [Google Scholar]
- Lee, D. Plant pigments and their manipulation. In Annual Plant Reviews, 2004th ed.; Davies K.M., Ed.; Blackwell Publishing: Oxford, UK; CRC Press: Boca Raton, FL, USA, 2005; Volume 12, 352p. Ann. Bot-Lond. 2005, 96, 1332–1333. [Google Scholar] [CrossRef]
- Pocock, T.; Król, M.; Huner, N. The determination and quantification of photosynthetic pigments by reverse phase high-performance liquid chromatography, thin-layer chromatography, and spectrophotometry. Methods Mol. Biol. (CliftonNJ) 2004, 274, 137–148. [Google Scholar] [CrossRef]
- Lamb, J.J.; Rokke, G.; Hohmann-Marriott, M.F. Chlorophyll fluorescence emission spectroscopy of oxygenic organisms at 77 K. Photosynthetica 2018, 56, 105–124. [Google Scholar] [CrossRef]
- Sinha, R.K.; Komenda, J.; Knoppova, J.; Sedlarova, M.; Pospisil, P. Small CAB-like proteins prevent formation of singlet oxygen in the damaged photosystem II complex of the cyanobacterium synechocystis sp PCC 6803. Plant. Cell Env. 2012, 35, 806–818. [Google Scholar] [CrossRef] [PubMed]
- Boardman, N.K.; Thorne, S.W.; Anderson, J.M. Fluorescence properties of particles obtained by digitonin fragmentation of spinach chloroplasts. Proc. Natl. Acad. Sci. USA 1966, 56, 586. [Google Scholar] [CrossRef]
- Lamb, J.; Forfang, K.; Hohmann-Marriott, M. A practical solution for 77 K fluorescence measurements based on LED excitation and CCD array detector. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Linhartova, M.; Bucinska, L.; Halada, P.; Jecmen, T.; Setlik, J.; Komenda, J.; Sobotka, R. Accumulation of the Type IV prepilin triggers degradation of SecY and YidC and inhibits synthesis of Photosystem II proteins in the cyanobacterium synechocystis PCC 6803. Mol. Microbiol. 2014, 93, 1207–1223. [Google Scholar] [CrossRef]
- Johnsen, G.; Sakshaug, E. Biooptical characteristics of PSII and PSI in 33 species (13 pigment groups) of marine phytoplankton, and the relevance for pulse-amplitude-modulated and fast-repetition-rate fluorometry1. J. Phycol. 2007, 43, 1236–1251. [Google Scholar] [CrossRef]
- Trask, B.J.; Vandenengh, G.J.; Elgershuizen, J.H.B.W. Analysis of phytoplankton by flow-cytometry. Cytometry 1982, 2, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Van den Engh, G.J.; Doggett, J.K.; Thompson, A.W.; Doblin, M.A.; Gimpel, C.N.G.; Karl, D.M. Dynamics of prochlorococcus and synechococcus at station aloha revealed through flow cytometry and high-resolution vertical sampling. Front. Mar. Sci. 2017, 4. [Google Scholar] [CrossRef]
- Berla, B.M.; Saha, R.; Immethun, C.M.; Maranas, C.D.; Moon, T.S.; Pakrasi, H.B. Synthetic biology of cyanobacteria: Unique challenges and opportunities. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, R.; Shen, G.; Bryant, D.A.; Golbeck, J.H. Regulatory Rroles for IscA and SufA in iron homeostasis and redox stress responses in the cyanobacterium synechococcus sp. strain PCC 7002. J. Bacteriol. 2006, 188, 3182. [Google Scholar] [CrossRef]
- Ludwig, M.; Chua, T.T.; Chew, C.Y.; Bryant, D.A. Fur-type transcriptional repressors and metal homeostasis in the cyanobacterium synechococcus sp. PCC 7002. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Falk, S.; Samson, G.; Bruce, D.; Huner, N.P.A.; Laudenbach, D.E. Functional analysis of the iron-stress induced CP 43′ polypeptide of PS II in the cyanobacterium synechococcus sp. PCC 7942. Photosynth. Res. 1995, 45, 51–60. [Google Scholar] [CrossRef]
- Markley, A.L.; Begemann, M.B.; Clarke, R.E.; Gordon, G.C.; Pfleger, B.F. Synthetic biology toolbox for controlling gene expression in the cyanobacterium synechococcus sp strain PCC 7002. ACS Synth. Biol. 2015, 4, 595–603. [Google Scholar] [CrossRef]
- Vogel, A.I.M.; Lale, R.; Hohmann-Marriott, M.F. Streamlining recombination-mediated genetic engineering by validating three neutral integration sites in synechococcus sp PCC 7002. J. Biol. Eng. 2017, 11. [Google Scholar] [CrossRef]
- Xu, Y.; Alvey, R.M.; Byrne, P.O.; Graham, J.E.; Shen, G.; Bryant, D.A. Expression of genes in cyanobacteria: adaptation of endogenous plasmids as platforms for high-level gene expression in synechococcus sp. PCC 7002. In Photosynthesis Research Protocols; Carpentier, R., Ed.; Humana Press: Totowa, NJ, USA, 2011; pp. 273–293. [Google Scholar] [CrossRef]
- Zess, E.K.; Begemann, M.B.; Pfleger, B.F. Construction of new synthetic biology tools for the control of gene expression in the cyanobacterium synechococcus sp strain PCC 7002. Biotechnol. Bioeng. 2016, 113, 424–432. [Google Scholar] [CrossRef]
- Cohen, M.F.; Wallis, J.G.; Campbell, E.L.; Meeks, J.C. Transposon mutagenesis of nostoc sp strain Atcc-29133, a filamentous cyanobacterium with multiple cellular-differentiation alternatives. Microbiol. SGM 1994, 140, 3233–3240. [Google Scholar] [CrossRef]
- Holtman, C.K.; Chen, Y.; Sandoval, P.; Gonzales, A.; Nalty, M.S.; Thomas, T.L.; Youderian, P.; Golden, S.S. High-throughput functional analysis of the synechococcus elongatus PCC 7942 genome. DNA Res. 2005, 12, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Ruffing, A.M.; Jensen, T.J.; Strickland, L.M. Genetic tools for advancement of synechococcus sp PCC 7002 as a cyanobacterial chassis. Microb. Cell Fact. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Wendt, K.E.; Ungerer, J.; Cobb, R.E.; Zhao, H.M.; Pakrasi, H.B. CRISPR/Cas9 mediated targeted mutagenesis of the fast growing cyanobacterium synechococcus elongatus UTEX 2973. Microb. Cell Fact. 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, M.; Bryant, D.A. Transcription profiling of the model cyanobacterium synechococcus sp strain PCC 7002 by Next-Gen (SOLiD (TM)) sequencing of cDNA. Front. Microbiol. 2011, 2. [Google Scholar] [CrossRef]
- Hernandez-Prieto, M.A.; Futschik, M.E. CyanoEXpress: A web database for exploration and visualisation of the integrated transcriptome of cyanobacterium synechocystis sp. PCC6803. Bioinformation 2012, 8, 634–638. [Google Scholar] [CrossRef]
- Martin, S.A.M.; Dehler, C.E.; Król, E. Transcriptomic responses in the fish intestine. Dev. Comp. Immunol. 2016, 64, 103–117. [Google Scholar] [CrossRef]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef]
- Kono, N.; Arakawa, K. Nanopore sequencing: Review of potential applications in functional genomics. Dev. Growth Differ. 2019, 61, 316–326. [Google Scholar] [CrossRef]
- Burlacot, A.; Li-Beisson, Y.; Peltier, G. Membrane inlet mass spectrometry at the crossroads of photosynthesis, biofuel, and climate research. Plant. Physiol. 2020, 183, 451. [Google Scholar] [CrossRef]
- Wilson, S.T.; Böttjer, D.; Church, M.J.; Karl, D.M. Comparative assessment of nitrogen fixation methodologies, conducted in the oligotrophic North Pacific Ocean. Appl. Env. Microb. 2012, 78, 6516. [Google Scholar] [CrossRef]
- Katoh, H.; Grossman, A.R.; Hagino, N.; Ogawa, T. A gene of synechocystis sp strain PCC 6803 encoding a novel iron transporter. J. Bacteriol. 2000, 182, 6523–6524. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.B.; Lou, W.J.; Du, H.Y.; Price, N.M.; Qiu, B.S. Sll1263, a unique cation diffusion facilitator protein that promotes iron uptake in the cyanobacterium synechocystis sp strain PCC 6803. Plant. Cell Physiol. 2012, 53, 1404–1417. [Google Scholar] [CrossRef] [PubMed]
- L’Annunziata, M. (Ed.) Table of radioactive isotopes. In A Handbook of Radioactive Analysis, 3rd ed.; Elsevier Inc: Amsterdam, The Netherlands, 2012. [Google Scholar]
- PerkinElmer, Inc. Iron-55 Handling Precautions. Available online: https://www.perkinelmer.com/lab-solutions/resources/docs/TCH_Iron55.pdf (accessed on 5 August 2020).
- Perkin Elmer. Radiometric Detection Guide. Available online: https://www.perkinelmer.com/lab-solutions/resources/docs/BRO_RadiometricDetectionGuide.pdf (accessed on 20 September 2020).
- Strzepek, R.F.; Price, N.M. Influence of irradiance and temperature on the iron content of the marine diatom Thalassiosira weissflogii (Bacillariophyceae). Mar. Ecol. Prog. Ser. 2000, 206, 107–117. [Google Scholar] [CrossRef]
- Hudson, R.J.M.; Morel, F.M.M. Distinguishing between extracellular and intracellular iron in marine-phytoplankton. Limnol. Oceanogr. 1989, 34, 1113–1120. [Google Scholar] [CrossRef]
- Ecker, D.J.; Emery, T. Iron uptake from ferrichrome-a and iron citrate in ustilago-sphaerogena. J. Bacteriol. 1983, 155, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Kammler, M.; Schon, C.; Hantke, K. Characterization of the ferrous iron uptake system of Escherichia Coli. J. Bacteriol. 1993, 175, 6212–6219. [Google Scholar] [CrossRef] [PubMed]
- Strzepek, R.F.; Maldonado, M.T.; Hunter, K.A.; Frew, R.D.; Boyd, P.W. Adaptive strategies by Southern Ocean phytoplankton to lessen iron limitation: Uptake of organically complexed iron and reduced cellular iron requirements. Limnol. Oceanogr. 2011, 56, 1983–2002. [Google Scholar] [CrossRef]
- Jiang, H.B.; Lou, W.J.; Ke, W.T.; Song, W.Y.; Price, N.M.; Qiu, B.S. New insights into iron acquisition by cyanobacteria: An essential role for ExbB-ExbD complex in inorganic iron uptake. ISME J. 2015, 9, 297–309. [Google Scholar] [CrossRef]
- Lis, H.; Shaked, Y.; Kranzler, C.; Keren, N.; Morel, F.M.M. Iron bioavailability to phytoplankton: An empirical approach. ISME J. 2015, 9, 1003–1013. [Google Scholar] [CrossRef]
- Barbeau, K.; Rue, E.L.; Trick, C.G.; Bruland, K.T.; Butler, A. Photochemical reactivity of siderophores produced by marine heterotrophic bacteria and cyanobacteria based on characteristic Fe (III) binding groups. Limnol. Oceanogr. 2003, 48, 1069–1078. [Google Scholar] [CrossRef]
- Wada, E.; Ohki, K.; Yoshikawa, S.; Parker, P.L.; Baalen, C.V.; Matsumoto, G.I.; Aita, M.N.; Saino, T. Ecological aspects of carbon and nitrogen isotope ratios of cyanobacteria. Plankton Benthos Res. 2012, 7, 135–145. [Google Scholar] [CrossRef][Green Version]
- Gu, B.; Alexander, V. Stable carbon isotope evidence for atmospheric CO (inf2) uptake by cyanobacterial surface scums in a Eutrophic lake. Appl. Env. Microb. 1996, 62, 1803. [Google Scholar] [CrossRef] [PubMed]
- Ferrier-Pagès, C.; Leal, M.C. Stable isotopes as tracers of trophic interactions in marine mutualistic symbioses. Ecol. Evol. 2019, 9, 723–740. [Google Scholar] [CrossRef] [PubMed]
- Amor, M.; Busigny, V.; Louvat, P.; Tharaud, M.; Gélabert, A.; Cartigny, P.; Carlut, J.; Isambert, A.; Durand-Dubief, M.; Ona-Nguema, G.; et al. Iron uptake and magnetite biomineralization in the magnetotactic bacterium Magnetospirillum magneticum strain AMB-1: An iron isotope study. Geochim. Cosmochim. Acta 2018, 232, 225–243. [Google Scholar] [CrossRef]
- Sun, R.; Wang, B. Iron isotope fractionation during uptake of ferrous ion by phytoplankton. Chem. Geol. 2018, 481, 65–73. [Google Scholar] [CrossRef]
- Mulholland, D.S.; Poitrasson, F.; Shirokova, L.S.; González, A.G.; Pokrovsky, O.S.; Boaventura, G.R.; Vieira, L.C. Iron isotope fractionation during Fe (II) and Fe (III) adsorption on cyanobacteria. Chem. Geol. 2015, 400, 24–33. [Google Scholar] [CrossRef]
- Gault-Ringold, M.; Adu, T.; Stirling, C.H.; Frew, R.D.; Hunter, K.A. Anomalous biogeochemical behavior of cadmium in subantarctic surface waters: Mechanistic constraints from cadmium isotopes. Earth Planet. Sci. Lett. 2012, 341–344, 94–103. [Google Scholar] [CrossRef]
- Conway, T.M.; Rosenberg, A.D.; Adkins, J.F.; John, S.G. A new method for precise determination of iron, zinc and cadmium stable isotope ratios in seawater by double-spike mass spectrometry. Anal. Chim. Acta 2013, 793, 44–52. [Google Scholar] [CrossRef]
- Conway, T.M.; John, S.G. Quantification of dissolved iron sources to the North Atlantic Ocean. Nature 2014, 511, 212–215. [Google Scholar] [CrossRef]
- Ellwood, M.J.; Hutchins, D.A.; Lohan, M.C.; Milne, A.; Nasemann, P.; Nodder, S.D.; Sander, S.G.; Strzepek, R.; Wilhelm, S.W.; Boyd, P.W. Iron stable isotopes track pelagic iron cycling during a subtropical phytoplankton bloom. Proc. Natl. Acad. Sci. USA 2015, 112, E15. [Google Scholar] [CrossRef]
- Peak, D.; Regier, T. Direct observation of tetrahedrally coordinated Fe (III) in ferrihydrite. Env. Sci. Technol. 2012, 46, 3163–3168. [Google Scholar] [CrossRef] [PubMed]
- Hocking, R.K.; George, S.D.; Raymond, K.N.; Hodgson, K.O.; Hedman, B.; Solomon, E.I. Fe L-Edge X-ray absorption spectroscopy determination of differential orbital covalency of siderophore model compounds: Electronic structure contributions to high stability constants. J. Am. Chem. Soc. 2010, 132, 4006–4015. [Google Scholar] [CrossRef] [PubMed]
- Miot, J.; Benzerara, K.; Morin, G.; Kappler, A.; Bernard, S.; Obst, M.; Ferard, C.; Skouri-Panet, F.; Guigner, J.M.; Posth, N.; et al. Iron biomineralization by anaerobic neutrophilic iron-oxidizing bacteria. Geochim. Cosmochim. Acta 2009, 73, 696–711. [Google Scholar] [CrossRef]
- Von der Heyden, B.P.; Roychoudhury, A.N.; Mtshali, T.N.; Tyliszczak, T.; Myneni, S.C.B. Chemically and geographically distinct solid-phase iron pools in the Southern Ocean. Science 2012, 338, 1199–1201. [Google Scholar] [CrossRef]
- Von der Heyden, B.P.; Hauser, E.J.; Mishra, B.; Martinez, G.A.; Bowie, A.R.; Tyliszczak, T.; Mtshali, T.N.; Roychoudhury, A.N.; Myneni, S.C.B. Ubiquitous presence of Fe(II) in aquatic colloids and its association with organic carbon. Environ. Sci. Technol. Lett. 2014, 1, 387–392. [Google Scholar] [CrossRef]
- Lam, P.J.; Ohnemus, D.C.; Marcus, M.A. The speciation of marine particulate iron adjacent to active and passive continental margins. Geochim. Cosmochim. Acta 2012, 80, 108–124. [Google Scholar] [CrossRef]
- Kretschmer, X.C.; Meitzner, G.; Gardea-Torresdey, J.L.; Webb, R. Determination of Cu environments in the cyanobacterium Anabaena flos-aquae by X-Ray absorption spectroscopy. Appl. Env. Microb. 2004, 70, 771. [Google Scholar] [CrossRef]
- IPCC. Summary for policymakers. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part. A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment. Report of the Intergovernmental Panel on Climate Change; IPCC: Cambridge, UK; New York, NY, USA, 2014; pp. 1–32. [Google Scholar]
- IPCC. Climate Change 2013: The Physical Science Basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Cambridge, UK; New York, NY, USA, 2013; p. 1535. [Google Scholar]
- Trimborn, S.; Brenneis, T.; Hoppe, C.J.M.; Laglera, L.M.; Norman, L.; Santos-Echeandia, J.; Volkner, C.; Wolf-Gladrow, D.; Hassler, C.S. Iron sources alter the response of Southern Ocean phytoplankton to ocean acidification. Mar. Ecol. Prog. Ser. 2017, 578, 35–50. [Google Scholar] [CrossRef]
- Gledhill, M.; Achterberg, E.P.; Li, K.Q.; Mohamed, K.N.; Rijkenberg, M.J.A. Influence of ocean acidification on the complexation of iron and copper by organic ligands in estuarine waters. Mar. Chem. 2015, 177, 421–433. [Google Scholar] [CrossRef]
- Boyd, P.W.; Collins, S.; Dupont, S.; Fabricius, K.; Gattuso, J.P.; Havenhand, J.; Biswas, H.; McGraw, C.M.; Riebesell, U.; Vichi, M.; et al. SCOR WG149 Handbook to support the SCOR Best Practice Guide for Multiple Drivers Marine Research; SCOR: Hobart, Tasmania, 2019. [Google Scholar]
- Stockdale, A.; Tipping, E.; Lofts, S.; Mortimer, R.J.G. Effect of ocean acidification on organic and inorganic speciation of trace metals. Env. Sci. Technol. 2016, 50, 1906–1913. [Google Scholar] [CrossRef]
- Boyd, P.W.; Rynearson, T.A.; Armstrong, E.A.; Fu, F.; Hayashi, K.; Hu, Z.; Hutchins, D.A.; Kudela, R.M.; Litchman, E.; Mulholland, M.R.; et al. Marine phytoplankton temperature versus growth responses from polar to tropical waters—Outcome of a scientific community-wide study. PLoS ONE 2013, 8, e63091. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hunnestad, A.V.; Vogel, A.I.M.; Armstrong, E.; Digernes, M.G.; Ardelan, M.V.; Hohmann-Marriott, M.F. From the Ocean to the Lab—Assessing Iron Limitation in Cyanobacteria: An Interface Paper. Microorganisms 2020, 8, 1889. https://doi.org/10.3390/microorganisms8121889
Hunnestad AV, Vogel AIM, Armstrong E, Digernes MG, Ardelan MV, Hohmann-Marriott MF. From the Ocean to the Lab—Assessing Iron Limitation in Cyanobacteria: An Interface Paper. Microorganisms. 2020; 8(12):1889. https://doi.org/10.3390/microorganisms8121889
Chicago/Turabian StyleHunnestad, Annie Vera, Anne Ilse Maria Vogel, Evelyn Armstrong, Maria Guadalupe Digernes, Murat Van Ardelan, and Martin Frank Hohmann-Marriott. 2020. "From the Ocean to the Lab—Assessing Iron Limitation in Cyanobacteria: An Interface Paper" Microorganisms 8, no. 12: 1889. https://doi.org/10.3390/microorganisms8121889
APA StyleHunnestad, A. V., Vogel, A. I. M., Armstrong, E., Digernes, M. G., Ardelan, M. V., & Hohmann-Marriott, M. F. (2020). From the Ocean to the Lab—Assessing Iron Limitation in Cyanobacteria: An Interface Paper. Microorganisms, 8(12), 1889. https://doi.org/10.3390/microorganisms8121889