Abstract
Nosocomial diseases represent a huge health and economic burden. A significant portion is associated with the use of medical devices, with 80% of these infections being caused by a bacterial biofilm. The insertion of a foreign material usually elicits inflammation, which can result in hampered antimicrobial capacity of the host immunity due to the effort of immune cells being directed to degrade the material. The ineffective clearance by immune cells is a perfect opportunity for bacteria to attach and form a biofilm. In this study, we analyzed the antibiofilm capacity of three naturally derived biofilm inhibitors when combined with immune cells in order to assess their applicability in implantable titanium devices and low-density polyethylene (LDPE) endotracheal tubes. To this end, we used a system based on the coculture of HL-60 cells differentiated into polymorphonuclear leukocytes (PMNs) and Staphylococcus aureus (laboratory and clinical strains) on titanium, as well as LDPE surfaces. Out of the three inhibitors, the one coded DHA1 showed the highest potential to be incorporated into implantable devices, as it displayed a combined activity with the immune cells, preventing bacterial attachment on the titanium and LDPE. The other two inhibitors seemed to also be good candidates for incorporation into LDPE endotracheal tubes.
1. Introduction
Over 2.6 million new cases of healthcare-associated infections are annually reported just in the European Union [1], and over 33,000 result in death [2] due to the increasing number of antimicrobial-resistance cases [3]. At least 25% of these infections are associated with the use of medical devices, and 80% of them are estimated to be caused by bacterial biofilms [4,5]. Biofilms are defined as a community of microorganisms encased within a self-produced matrix that adheres to biological or nonbiological surfaces [6,7]. They are currently regarded as the most important nonspecific mechanism of antimicrobial resistance [8,9].
During the worldwide crisis of SARS-CoV-2, as well as in many other pathological conditions, mechanical ventilation is used to assist or replace spontaneous breathing as a life-saving procedure in intensive care. However, the use of endotracheal intubation also poses major risks in prolonged ventilation. The endotracheal tube provides an ideal opportunity for bacteria to form biofilms on both the outer and luminal surface of the tube, increasing the risk of pulmonary infection by 6 to 10 times [10,11,12], with Staphylococcus spp. and Pseudomonas aeruginosa among the most frequent colonizing agents [13]. Colonization by microorganisms and the subsequent formation of a biofilm can happen within hours [13], but these kinds of devices are relatively easy to replace. On the other hand, infection of orthopedic implants is particularly problematic, as these devices remain in the body, often causing chronic and/or recurring infections mediated by biofilms. These infections also frequently require removal of the infected implant, thereby causing implant failure [14,15,16]. Given the rising number of implantations, the absolute number of complications is inevitably increasing at the same pace, causing not only distress for the patients but also an increasing economic burden [15,16].
The most common causative agents of infection in orthopedic implants are Gram-positive cocci of the genus Staphylococcus, e.g., Staphylococcus aureus and Staphylococcus epidermidis [17]. In the absence of a foreign body, contaminations caused by these opportunistic pathogens are usually cleared by the immune system. In contrast, the placement of an implant per se represents a risk factor for the development of a chronic infection. This is due to the fact that the surgical procedure causes tissue damage resulting in the local generation of damage-associated molecular patterns (DAMPs), endogenous danger molecules that are released from damaged or dying cells and activate the innate immune system by interacting with pattern recognition receptors (PRRs) [18]. This is sensed by host neutrophils, which migrate to the injured tissue sites, activating defense mechanisms, such as the generation of oxygen-derived and nitrogen-derived reactive species as well as phagocytosis, to unsuccessfully attempt to clear the foreign material. These events lead to immune cell exhaustion and death, and tissue damage caused by the triggered inflammation eventually leads to a niche of immune suppression around the implant [19]. Under these specific conditions, the clearance of planktonic bacteria by immune cells becomes impaired [14], which predisposes the implant to microbial colonization and biofilm-mediated infection.
However, it is possible that not only host immune cells and bacterial cells can be present at the moment of implantation, but also the cells of the tissue where the material is being implanted. The “race for the surface” concept describes the competition between bacteria and the host cells of the tissue where the device is implanted to colonize the surface of the material [20]. The rapid integration of the material into the host tissue is a key component in the success of an implant, as the colonization of the surface by the cells of the host not only ensures correct integration, but also prevents bacterial colonization [21]. However, if bacterial adhesion occurs first, the host defense system is unable to prevent the colonization and subsequent biofilm formation [22].
One of the current challenges to prevent and resolve biofilm-mediated infections is the limited repertoire of compounds that are able to act on them at sufficiently low concentrations [23]. Because of this, there is intense ongoing research focused on the search for new biofilm inhibitors by means of chemical screenings. However, for compounds to be truly effective when used for the protection of biomaterials in translational applications, they must be tested in meaningful experimental models. Based on this, we previously optimized an in vitro system based on the coculture of SaOS-2, osteogenic cells, and S. aureus (laboratory and clinical strains) on a titanium surface (Reigada; et al. [24]), and studied the effect of three naturally derived biofilm inhibitors (Figure 1). Two of them were dehydroabietic acid (DHA) derivatives, namely, N-(abiet-8,11,13-trien-18-oyl)cyclohexyl-L-alanine and N-(abiet-8,11,13-trien-18-oyl)-D-tryptophan, coded DHA1 (Figure 1a) and DHA2 (Figure 1b), respectively. The third one was a flavan derivative, 6-chloro-4-(6-chloro-7-hydroxy-2,4,4-trimethylchroman-2-yl)benzene-1,3-diol, coded FLA1 (Figure 1c). All of them were previously reported by our group and demonstrated to display activity in preventing biofilm formation, as well as disrupting preformed S. aureus biofilms on 96-well plates [25,26], but the testing in the coculture model with osteogenic cells provided with new insights into their applicability as part of anti-infective implantable devices.
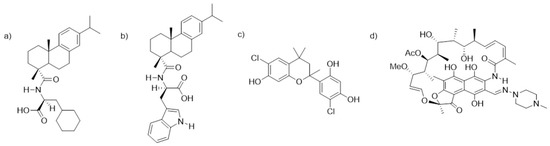
Figure 1.
Chemical structures of the two dehydroabietic acid (DHA) derivatives, (a) N-(abiet-8,11,13-trien-18-oyl)cyclohexyl-L-alanine, (b) N-(abiet-8,11,13-trien-18-oyl)-D-tryptophan, coded DHA1 and DHA2, the flavan derivative, (c) 6-chloro-4-(6-chloro-7-hydroxy-2,4,4-trimethylchroman-2-yl)benzene-1,3-diol, coded FLA1, and (d) rifampicin, coded RIF.
The coculture model developed previously [24] involving SaOS-2 cells and S. aureus strains offered an in vitro environment that was closer in terms of host–bacteria interactions and substrate materials to the in vivo scenario of the implanted device. Moreover, in terms of antimicrobial evaluation, it provided information not only regarding the antibiofilm activity but also about the effects on tissue integration. However, this model did not assess the effect of the antimicrobials on immune cells. As mentioned earlier, chronic inflammation not only lowers the antimicrobial efficacy, but also complicates the integration of the implant material as a consequence of maintained inflammation and tissue damage. Therefore, it is essential to assess the effect that antimicrobials might have in the presence of immune cells, particularly for those intended to be used in medical devices. Endotracheal tubes significantly differ from orthopedic implants in material, function, and implantation procedure, but they both cause impairment of the host immune antimicrobial capacity.
Because of this, in this contribution, we move one step forward toward emulating in vivo conditions encountered by medical devices in an in vitro setting. In this case, we introduce immune cells, specifically neutrophils, in a coculture model with bacterial cells. Neutrophils were selected as immune cells as they are the first cell types to migrate toward damaged tissue cells [27]. Our aim was to develop cocultures of S. aureus and differentiated HL-60 cells, grown on titanium coupons or low-density polyethylene (LDPE) tubes, to simulate biofilm formation on orthopedic implants or endotracheal tubes, respectively. As a proof-of-concept, we also study the possible antimicrobial effects of naturally derived antibiofilm inhibitors DHA1, DHA2, and FLA1, and their possible applicability as part of medical devices.
2. Materials and Methods
2.1. Compounds
The dehydroabietic acid derivatives coded DHA1 and DHA2 (previously coded 11 and 9b, respectively) were synthesized according to [25]. Their spectral data were identical to those reported in [28]. The flavan derivative coded FLA1 (previously coded 291 in [26]), was purchased from TimTec (product code: ST075672, www.timtec.net). These compounds were selected given their low minimum inhibitory concentrations (MICs) and the low concentrations needed in order to prevent S. aureus biofilm formation (Table S1). The control antibiotic rifampicin was purchased from Sigma-Aldrich (CAS number 13292-46-1) and coded RIF. Molecular structures are shown in Figure 1.
2.2. Bacterial Strains
Bacterial studies were performed using the laboratory strain S. aureus ATCC 25923 (American Type Culture Collection, Manassas, VA, USA) and one clinical strain isolated from hip prostheses and osteosynthesis implants at the Hospital Fundación Jiménez Díaz (Madrid, Spain) [29] (coded S. aureus P2).
2.3. HL-60 Cell Culture and Differentiation
HL-60 (ATCC CCL-240) cells were grown and maintained in Roswell Park Memorial Institute (RPMI) 1640 Medium (R8758, Sigma Aldrich, St. Louis, MO, USA), supplemented with 20% (v/v) heat-inactivated fetal bovine serum (FBS) (Sigma Aldrich, St. Louis, MO, USA) and 1% (v/v) penicillin/streptomycin (Sigma Aldrich, St. Louis, MO, USA). Cells were maintained in suspension at a concentration between 105–106 cells/mL in 72 cm2 culture flasks (VWR, Radnor, PA, USA) at 37 °C in 5% CO2 in a humidified incubator. N,N–Dimethylformamide (DMF)(Sigma Aldrich, St. Louis, MO, USA) was utilized in order to differentiate the cells into polymorphonuclear-like cells [30]. In order to carry out the differentiation, cells were cultured for 6 days in the maintenance medium at a concentration of 100 mM DMF. The success of the differentiation was assessed visually after Giemsa staining using a Leica DMLS microscope (Leica, Wetzlar, Germany). Differentiated cells were neutrophil-like, with a multilobar nucleus and a fairly clear cytoplasm.
2.4. Biofilm Prevention Efficacy of Differentiated HL-60 Cells against Different Bacterial Concentrations of S. aureus ATCC 25923
2.4.1. Bacterial Inoculum Preparation
Pure colonies (2–3) of S. aureus ATCC 25923 were added to 5 mL of tryptic soy broth (TSB, Neogen®, Lansing, MI, USA) and incubated overnight at 37 °C, 120 rpm. After incubation, 10 µL of the preculture was added to 5 mL fresh TSB and incubated for 3–4 h (37 °C, 120 rpm), in order to obtain a midlogarithmic growth phase of bacteria. Bacterial cultures were washed twice with sterile phosphate-buffered saline (PBS; 140 mM NaCl, pH 7.4) and the concentration was adjusted to 2 × 108 in RPMI 1640 based on the optical density of the suspension at 600 nm. From this stock, serial dilutions were made in RPMI 1640 between 2 × 108 and 2 × 102 CFU/mL.
2.4.2. Immune Cell (HL-60) Preparation
Twenty-four hours prior to starting the experiments, the media of the differentiated HL-60 cells were refreshed with nonsupplemented RPMI 1640 in order to remove possible traces of antibiotics from the maintenance media. Differentiated HL-60 cells (after 6 days exposure to a concentration of 100 mM DMF) were counted with a Countess™ II automated cell counter (Thermo Scientific, Waltham, MA, USA) and adjusted to a concentration of 2 × 105 cells/mL. In order to test the influence of activating the cells with phorbol 12-myristate 13-acetate (PMA) (Sigma Aldrich, St. Louis, MO, USA) on the antimicrobial capacity, half of the suspension of differentiated HL-60 cells was incubated for 20 min in RPMI 1640 supplemented with 25 nM PMA. PMA-activated and nonactivated HL-60 cells were separately added (100 µL) to flat-bottomed, 96-well microplates (Nunclon Δ surface, Nunc, Roskilde, Denmark).
2.4.3. Coculture of S. aureus ATCC 25923 and HL-60 Cells
S. aureus ATCC 25923 suspensions at the different concentrations (100 µL) were added to the wells of the 96-well microplate already containing 100 µL of one of the two different HL-60 cell suspensions (activated or nonactivated). In the bacterial control wells, 100 µL of the S. aureus ATCC 25923 suspension at different concentrations were added to wells containing 100 µL of RPMI 1640 alone. The wells corresponding to the HL-60 cell control consisting of a suspension of HL-60 cells at a concentration 105 cells/mL in RPMI 1640 were used to observe the cell morphology after 18 h of incubation, but no quantitative viability test was carried out. The 96-well microplates were incubated for 18 h at 37 °C and 5% CO2 in a humidified incubator.
2.4.4. Biofilm Quantification in 96-Well Microplates
After S. aureus ATCC 25923 biofilms were grown, the media were carefully removed and each well was washed twice with 125 µL sterile PBS, followed by the addition of 150 µL of TSB. Each 96-well microplate was closed with a plastic seal and parafilm and placed in a plastic bag, which was sealed with heat in order to prevent leakage. The plate was sonicated for 10 min at 35 kHz in an Ultrasonic Cleaner 3800 water bath sonicator (Branson Ultrasonics, Danbury, CT, USA) at 25 °C. This procedure did not affect the viability of the staphylococci (Figure S1).
S. aureus ATCC 25923 was then resuspended in RPMI 1640 using 3 pipetting cycles (up/down). Samples (10 µL) from each tested condition were transfer to 90 µL of TSB and serial dilutions were made from 10−1 up to 10−7. Aliquots (10 µL) of all dilutions were transferred to sheep blood agar plates (Amsterdam UMC, Amsterdam, The Netherlands) and incubated at 37 °C overnight. Viable plate colonies were counted the next day.
2.5. Influence of Opsonizing S. aureus ATCC 25923 on the Efficacy of HL-60 Cells in Preventing Bacterial Attachment on Titanium Coupons
S. aureus ATCC 25923 suspensions were prepared as described in Section 2.4.1. The inoculum was adjusted to 2 × 107 CFU/mL and opsonized using 50% (v/v) pooled human serum in PBS (pooled from 15 healthy blood donors) by incubating at 37 °C for 30 min with gentle agitation, washing twice with PBS, and resuspending in RPMI 1640 [31]. As a control, a nonopsonized suspension of S. aureus ATCC 25923 was used. The two different suspensions (opsonized and nonopsonized), were then added to sterilized titanium coupons (0.4 cm height, 1.27 cm diameter, BioSurface Technologies Corp., Bozeman, MT, USA).
On the other hand, HL-60 cells were prepared as described in Section 2.4.2 and added to the titanium coupons, to which the bacterial suspension was previously added. The final volume covering each coupon was 1 mL, made up of 500 µL of a suspension of 2 × 107 CFU/mL S. aureus ATCC 25923, and 500 µL of a suspension of 2 × 105 HL-60 cells/mL in RPMI 1640.
2.6. Effect of the Antimicrobial Compounds on the Prevention of S. aureus ATCC 25923 and S. aureus P2 Adhesion in Coculture with Differentiated HL-60 Cells on Titanium Coupons
2.6.1. Culture of Staphylococci and HL-60 Cells
Bacterial inocula of S. aureus ATCC 25923 and S. aureus P2 were prepared as described in Section 2.4.1. The concentration was adjusted to 2 × 107 CFU/mL in RPMI 1640. For the differentiated HL-60 cells, the media were refreshed with RPMI 1640 24 h before the experiments started in order to remove possible traces of antibiotics from the maintenance media. On the day of the experiment, cells were counted with a Countess™ II automated cell counter (Thermo Scientific, Waltham, MA, USA) and adjusted to a concentration of 2 × 105 cells/mL in RPMI 1640. Each suspension (bacterial and HL-60 cells suspensions, 500 µL of each) was added to sterilized titanium coupons that were inserted in the different wells of a 24-well plate (Nunclon Δ surface, Nunc, Roskilde, Denmark) and to which the different tested compounds or the control antibiotics were added at a concentration of 50 µM (0.25% DMSO).
The 24-well plates containing the titanium coupons were maintained in cocultures with a solution containing 107 CFU of S. aureus ATCC 25923 or S. aureus P2 and 105 HL-60 cells in a total volume of 1 mL of RPMI 1640 for 24 h. Titanium coupons with added RIF (50 μM, 0.25% DMSO) were used as positive antibiotic controls. As coculture controls, titanium coupons without the addition of the tested antimicrobial compounds or control antibiotics were exposed simultaneously to both cellular systems (S. aureus and HL-60 cells) at the concentrations previously described. In addition, bacterial controls (exposed or not to the antimicrobial compounds in the absence of differentiated HL-60 cells) were also included.
2.6.2. Bacterial Adherence on Titanium Coupons
Titanium coupons were gently washed with TSB to remove remaining adhering planktonic cells and transferred to Falcon tubes containing 1 mL of 0.5% (w/v) Tween® 20-TSB solution. Next, the tubes were sonicated in an Ultrasonic Cleaner 3800 water bath sonicator (Branson Ultrasonics, Danbury, CT, USA) at 25 °C for 5 min at 35 kHz. The tubes were vortexed for 20 s prior to and after the sonication step. Serial dilutions of the resulting bacterial suspensions were made from 10-1 up to 10−7 and plated on tryptic soy agar (Neogen®, Lansing, MI, USA) plates.
2.7. Effect of the Antimicrobial Compounds on the Prevention of S. aureus ATCC 25923 Adhesion in Coculture with Differentiated HL-60 Cells on LDPE Tubes
The assay was carried out as described in Section 2.6 but instead of titanium coupons the materials added to the 24-well plates were 1 cm long sections of a sterilized fine bore LDPE tubing (Smiths Medical ASD, Minneapolis, MN, USA).
2.8. Scanning Electron Microscopy (SEM)
In order to visualize the effect of the antimicrobial compounds on the prevention of S. aureus ATCC 25923 adhesion in coculture with immune cells on titanium coupons, S. aureus ATCC 25923 was cocultured with differentiated HL-60 cells on titanium coupons, as described in Section 2.6. Prior to SEM, samples were fixed in 4% (v/v) paraformaldehyde and 1% (v/v) glutaraldehyde (Merck, Kenilworth, NJ, USA) overnight at room temperature. Samples were rinsed twice with distilled water, with the duration of each cycle being 10 min. The dehydration procedure consisted of 2 cycles of incubation for 15 min in 50% (v/v) ethanol, 2 cycles of incubation for 20 min in 70% (v/v) ethanol, 2 cycles of incubation for 30 min in 80% (v/v) ethanol, 2 cycles of incubation for 30 min in 90% (v/v) ethanol, and 2 cycles of incubation for 30 min in 100% (v/v) ethanol. In order to reduce the sample surface tension, samples were immersed in hexamethyldisilazane (Polysciences Inc., Warrington, FL, USA) overnight and air-dried. Before imaging, samples were mounted on aluminum SEM stubs and sputter-coated with a 4 nm platinum–palladium layer using a Leica EM ACE600 sputter coater (Leica Microsystems, Wetzlar, Germany). Images were acquired at 2 kV using a Zeiss Sigma 300 SEM (Zeiss, Oberkochen, Germany) at the Electron Microscopy Center Amsterdam (Amsterdam UMC).
Of each coupon, 8–10 fields were viewed and photographed at magnifications of 250×, 500×, 1000×, and 3000×. Representative images are shown in the results.
2.9. Statistical Analysis
The quantitative data are reported as the mean and standard deviation (SD) of at least three independent experiments. Data were analyzed using GraphPad Prism 8 for Windows. For statistical comparisons, Tukey’s multiple comparison test and Welch’s unpaired t-test were applied, and p < 0.05 was always considered as statistically significant.
3. Results
3.1. Effect of PMA Activation of Differentiated HL-60 Cells on Prevention of S. aureus ATCC 25923 Biofilm Formation
Before performing the experiments with the titanium coupons and the antimicrobial compounds, the initial concentration of S. aureus ATCC 25923 was determined where the bacteria were able to form a biofilm in absence of the HL-60 cells, but were prevented from forming a biofilm when cocultured with 105 HL-60 cells. At the same time, it was assessed whether the activation of the HL-60 cells with PMA enhanced their bacterial clearance capability (Figure 2). At an initial S. aureus ATCC 25923 concentration of 108 CFU/mL, HL-60 cells did not significantly affect the bacterial attachment in 96-well microplates. A reduction on the adhered S. aureus ATCC 25923 viable cell counts was observed at an S. aureus ATCC 25923 concentration of 107 CFU/mL and below (p < 0.001 in all cases when comparing the bacterial control with bacteria cocultured with HL-60 cells, and p = 0.001 for cells activated with PMA and a starting inoculum of 103 CFU/Ml), with the exception of the lowest bacterial concentration tested, i.e., 102 CFU/mL, where no difference was found. Both PMA-activated and nonactivated HL-60 cells (gray and green columns, respectively) showed similarly reduced numbers of adherent S. aureus ATCC 25923 viable cell counts at 24 h, so it was concluded that activation with PMA does not significantly enhance S. aureus ATCC 25923 clearance.
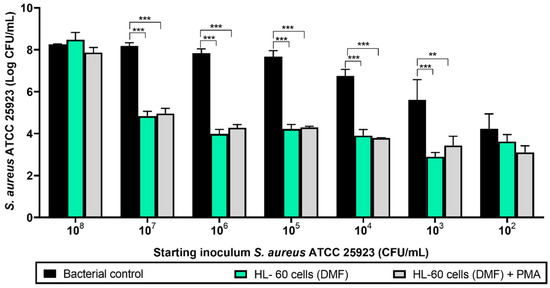
Figure 2.
Viable counts of 24-hour-old biofilms formed by different concentrations of S. aureus ATCC 25923 cocultured with HL-60 cells on 96-well microplates. Black columns represent the bacterial control (viable attached cells in the absence of HL-60 cells). Green columns show the coculture of S. aureus ATCC 25923 with HL-60 cells differentiated with N,N-dimethylformamide (DMF). Gray columns show the coculture of S. aureus ATCC 25923 with HL-60 cells differentiated with DMF and activated with phorbol 12-myristate 13-acetate (PMA). Values are means and SD of three independent experiments (*** p < 0.001; ** p < 0.01).
Based upon these results, the optimal initial S. aureus ATCC 25923 concentration was found to be 107 CFU/mL, which was then selected for the rest of the experiments. Since PMA activation of the HL-60 cells did not influence their phagocytic activity, no PMA stimulation was performed in subsequent experiments.
3.2. Influence of Opsonizing S. aureus ATCC 25923 on the Efficacy of HL-60 Cells in Preventing Bacterial Attachment on Titanium Coupons
One of the most relevant mechanisms of the host defense against Staphylococcus spp. is phagocytosis. Given that opsonization of S. aureus is important for neutrophils to be able to clear planktonic S. aureus [32], the phagocytic efficacy of HL-60 cells, as well as the impact of opsonization of S. aureus ATCC 25923, in preventing S. aureus ATCC 25923 biofilm formation on titanium coupons was simultaneously explored (Figure 3). The left part of Figure 3 shows how the preventive capability of HL-60 cells was preserved when tested on titanium surfaces (p < 0.001, when comparing S. aureus + HL-60 (gray column) with the bacterial control (black column)). This bacterial clearance activity was also observed when S. aureus ATCC 25923 was opsonized (p = 0.007), but no significant differences were found when comparing the effects of the HL-60 cells on opsonized versus nonopsonized S. aureus ATCC 25923 (p = 0.260). The antibacterial effects of PMA-activated HL-60 cells against opsonized S. aureus ATCC 25923 was additionally explored, but no differences were found with the antimicrobial activity of nonactivated HL-60 against nonopsonized S. aureus ATCC 25923 (Figure S2). Because opsonization did not enhance the bacterial clearance capacity of HL-60, the use of opsonized S. aureus during the rest of the experiments was decided against.
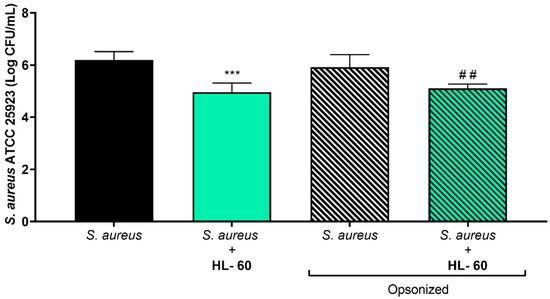
Figure 3.
Viable counts of adhered opsonized and nonopsonized S. aureus ATCC 25923 on titanium coupons after coculture with HL-60 cells for 24 h. The left half of the graph includes the nonopsonized S. aureus ATCC 25923 biofilm formation, in absence (black column) or cocultured with HL-60 cells (green column). The right half includes the opsonized S. aureus ATCC 25923 biofilm formation, in absence (white/black column) or cocultured with HL-60 cells (green/black column). “*” indicates statistical differences with the nonopsonized S. aureus ATCC 25923 in monoculture, while “#” represents statistical differences with the opsonized S. aureus ATCC 25923 in monoculture with Welch’s unpaired t-test (*** p < 0.001; ## p < 0.01). Values are means and SD of three independent experiments.
3.3. Effect of the Antimicrobial Compounds on the Prevention of S. aureus Adhesion in Coculture with Differentiated HL-60 Cells on Titanium Coupons
The effects of the three biofilm inhibitors DHA1, DHA2, and FLA1 and one control antibiotic, RIF, on the prevention of S. aureus attachment on titanium coupons were investigated using two strains, the laboratory strain ATCC 25923 (Figure 4a) and the clinical isolate P2 (Figure 4b), either in the absence (gray bars) or presence of HL-60 cells (green bars). In the absence of HL-60 cells, all of the tested antimicrobial compounds, as well as the control antibiotic, significantly reduced the attachment of S. aureus ATCC 25923 (p < 0.001, p = 0.040, p < 0.001, and p < 0.001, for DHA1, DHA2, FLA1, and RIF versus the control, respectively). In the case of the clinical strain (Figure 4b), all the antimicrobial compounds, except DHA2, also showed antimicrobial activity in the absence of HL-60 cells (p = 0.0067, p < 0.001, and p = 0.0087 versus control for DHA1, FLA1, and RIF, respectively).
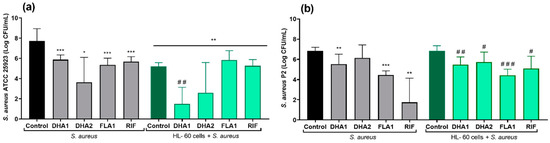
Figure 4.
Viable counts of adhered S. aureus ATCC 25923 (a) and S. aureus P2 (b) on titanium coupons exposed to different antimicrobial compounds (tested at 50 µM) after 24 h of incubation. The gray bars show the attached viable bacteria when exposed just to the antimicrobial compounds, while the green bars show the results when S. aureus strains were cocultured with HL-60 cells. “*” indicates statistical differences with the control in monoculture while “#” represents statistical differences with the control in cocultured controls with Welch’s unpaired t-test (* p < 0.05; ** p < 0.01; *** p < 0.001)/ # p < 0.05; ## p < 0.01; ### p< 0.001). Values are means and SD of three independent experiments.
The right part of Figure 4a (green bars) corresponds to the same experiments performed in the presence of HL-60 cells. Under such conditions, there was also a significant reduction of the attached viable S. aureus ATCC 25923 when compared with the material incubated without HL-60 cells (green control bar versus black control bar, p < 0.001). Moreover, this reduction was further increased by compound DHA1 (p = 0.025 when comparing the coculture control column with the DHA1 column in the HL-60 cells + S. aureus ATCC 25923 section of Figure 4, and p < 0.001 when comparing the DHA1 gray column with the DHA1 green column). The same tendency was observed for DHA2, but no statistical differences were found when comparing this to the control (dark green control: HL-60 cells + S. aureus ATCC 25923 section of Figure 4). Despite the fact that FLA1 and the model antibiotic (RIF) successfully prevented the adhesion of S. aureus ATCC 25923, they did not cause a further increase in the bacterial clearance activity of the HL-60 cells. In contrast, the mere presence of HL-60 cells did not result in a significant reduction of S. aureus P2 attachment (Figure 4b), since no differences were found between the S. aureus P2 control in monoculture and in coculture with HL-60 cells. In this case, no differences were found between the antibacterial effects of the compounds in monoculture when compared with the same treatment in coculture with HL-60 cells (DHA1, DHA2, FLA1, and RIF gray columns versus their corresponding green columns).
Using SEM imaging, it was visually confirmed that DHA1 reduced the number of S. aureus ATCC 25923 adherent to the titanium surface (Figure 5). In fact, almost no cocci were observed on the DHA1-treated titanium across the entire coupon. In addition, the presence of HL-60 cells also reduced the bacterial attachment, which was further enhanced by the treatment of DHA1. Adherent HL-60 cells were observed, as seen in the last row of images.

Figure 5.
Representative images acquired by SEM. From left to right the same section of the titanium coupon is shown with different magnification, 500, 1000 and 3000×. Upper row of images, bacterial control, i.e., coupons incubated with S. aureus ATCC 25923 only. Second row, titanium coupons incubated with S. aureus ATCC 25923 and treated with DHA1. Third row, cocultured control, titanium coupons incubated with S. aureus ATCC 25923 and HL-60 cells simultaneously. Fourth row, titanium coupons cocultured with S. aureus ATCC 25923 and HL-60 and treated with DHA1.
3.4. Effect of the Antimicrobial Compounds on the Prevention of S. aureus ATCC 25923 Adhesion in Coculture with Differentiated HL-60 Cells on LDPE tubes
The adhesive capacity of S. aureus, as well as the antimicrobial effect of different compounds, is known to significantly differ depending on the material [33]. For this reason, the applicability of the compounds as part of endotracheal tubes was tested on a clinically relevant material, LDPE. Figure 6 shows the effects of the tested compounds and the control antibiotic (RIF) on the prevention of S. aureus ATCC 25923 attachment on LDPE tubes. The left part of the figure shows that all the antimicrobial compounds significantly reduced the numbers of attached viable S. aureus ATCC 25923 cells, in the absence of HL-60 (p < 0.001, p = 0.0035, p < 0.001, and p < 0.001, when comparing, respectively, DHA1, DHA2, FLA1, and RIF with the control). Adding HL-60 cells resulted in significant prevention of S. aureus ATCC 25923 attachment to LDPE tubes (p < 0.001 when comparing the coculture control (dark green column) with the bacterial control (black column)). In this case, all the antimicrobial compounds looked to be able to further potentiate this bacterial clearance capability (p < 0.001, p = 0.023, p = 0.002, and p < 0.001, when comparing, respectively, DHA1, DHA2, FLA1, and RIF light green columns with the control dark green column). However, similarly to what was observed on the titanium model, it was only the DHA1 treatment that further potentiated the action of HL-60 cells against S. aureus ATCC 25923 (p = 0.015, when comparing the DHA1 gray column with the DHA1 green column). No differences were found between the viable cells (CFU/mL) attached on the LDPE exposed to S. aureus ATCC 25923 and treated with DHA2, FLA1, and RIF in monoculture and those exposed to both S. aureus ATCC 25923 and HL-60 with the same treatments (gray columns versus green columns). Full bacterial clearance was detected in LDPE tubes in the presence of DHA1 and RIF (i.e., no viable bacterial counts measured), where cocultures of S. aureus ATCC 25923 with differentiated HL-60 cells were formed. These findings further highlight the relevance of DHA1 as an antimicrobial candidate for incorporation into medical devices.
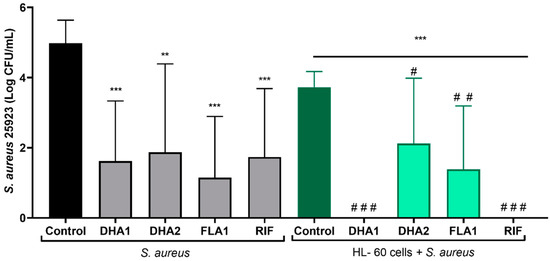
Figure 6.
Viable counts of adhered S. aureus ATCC 25923 on LDPE tubes exposed to different antimicrobial compounds (tested at 50 µM) after 24 h of incubation. The gray bars show the attached viable bacteria when exposed just to the antimicrobial compounds, while the green bars show the results when S. aureus strains were cocultured with HL-60 cells. “*” indicates statistical differences with the control in monoculture, while “#” represents statistical differences with the control in cocultured controls with Welch’s unpaired t-test (** p < 0.01; *** p < 0.001)/(# p < 0.05; ## p < 0.01; ### p < 0.001). Values are means and SD of three independent experiments.
4. Discussion
In this study, we explored the potential of incorporation of three previously identified naturally derived biofilm inhibitors into medical devices, particularly for titanium implantable devices and LDPE endotracheal tubes. From the three tested antimicrobial compounds, in line with previous findings [24], DHA1 appeared to be the best candidate for incorporation as part of implantable medical devices. All of the compounds proved to be interesting candidates to include into anti-infective endotracheal tubes, but it was DHA1 that again showed itself to be the most promising candidate, as it was the only one that significantly increased the bacterial clearance capacity of HL-60 cells against S. aureus ATCC 25923.
For compounds to be truly effective when used for protection of biomaterials in translational applications, they must be tested in meaningful experimental models. The insertion of any device provokes an acute inflammatory response that may cause ineffectiveness of innate immune cells such as neutrophils in cleaning planktonic bacteria, since these cells are directed to degrade the material. For this reason, it is vitally important to study the effects of antimicrobial compounds on neutrophils to assess their suitability as part of medical devices.
Before this investigation, no published reports existed on the effect of the two DHA derivatives (DHA1 and DHA2) on neutrophils, however, the parent compound (dehydroabietic acid, DHA) was previously reported to have slight toxicity toward this cell type [34]. This prior knowledge further justified the need for an assessment of the effects of DHA1 and DHA2 on the bacterial clearance capacity of neutrophils. Similarly, no data existed on effects of FLA1 on neutrophils, but other flavan derivatives were studied. Out of 10 different flavan-3-ol derivatives tested on human neutrophils, only two presented a slight toxic effect toward the neutrophils, but all of them reduced reactive oxygen species (ROS) and interleukin-8 production [35]. On the other hand, flavan-3-ol derivatives extracted from Bistorta officinalis (Delarbre) were reported to inhibit tumor necrosis factor-α (TNF-α) release from neutrophils [36]. These earlier findings were promising in terms of applying the flavan derivative FLA1 as part of medical devices, since its antimicrobial capacity in combination with its anti-inflammatory effects could result in prevention of infection while providing ideal cues toward material integration and resolution of inflammation.
In this study, we utilized HL-60 cells differentiated to polymorphonuclear-like cells in order to study the effects of the antimicrobial compounds in the presence of neutrophils. Alternatively, freshly extracted neutrophils could also have been used, but in such case differences would be encountered between individual donors in terms of reproducibility, the total number of cells that can be harvested, their short lifespan, or the disturbances in their physiology due to isolation procedures [37,38].
The three tested antimicrobial compounds and the control antibiotic (RIF) reduced S. aureus ATCC 25923 adhesion to titanium in the absence of HL-60 at the tested concentration of 50 µM. In the case of the clinical S. aureus strain, all compounds, except DHA2, significantly reduced S. aureus P2 biofilm formation on titanium in the absence of HL-60. The prevention of biofilm formation by DHA1 and FLA1 was as expected, as the compounds were used at concentrations higher than their MIC values [25,26]. Compound DHA2 showed some prevention activity, despite being tested at a concentration slightly below its MIC (i.e., 60 µM) [26]. As with most antimicrobials, cytotoxicity is a concern, and DHA2 was shown to reduce viability of HL cells (originating from the human respiratory tract) at a concentration of 100 µM [25].
The mere presence of HL-60 cells significantly reduced the bacterial attachment of S. aureus ATCC 25923. In contrast, this effect was not observed for the clinical S. aureus strain P2. This was also observed in our previous study, with the results obtained with the laboratory strain S. aureus ATCC 25923 significantly differing from the ones obtained with S. aureus P2 under the same experimental conditions. The latter has a key relevance in assessing the applicability of biofilm inhibitors as part of titanium implantable devices, as it was isolated from patients with orthopedic device-related infections. For this reason, the additional measurement of the preventive capacity of the biofilm inhibitors against the clinical S. aureus P2 strain is of great relevance.
Compound DHA1 was shown to further potentiate S. aureus ATCC 25923 clearance caused by HL-60, and this effect was further confirmed by SEM. This compound also managed to effectively prevent the adherence of the clinical strain (S. aureus P2), but in this case it was difficult to establish if the reduction in bacterial attachment caused by DHA1 (as well as FLA1 and RIF) was due to a combined antimicrobial effect with the HL-60 cells or if it was due to their intrinsic antimicrobial capacity, as their effects in monoculture were equal to those observed in coculture with the HL-60 cells. These results emphasized the importance of not limiting the in vitro experimentation to laboratory strains, especially in cases aimed at finding compounds effective against medical device-associated infections, as the results obtained with laboratory strains may overestimate the efficacy of the compound.
These results demonstrate that DHA1 seems to further increase S. aureus ATCC 25923 clearance by HL-60, while none of the other compounds negatively affect the antimicrobial effect of these immune cells. This is also of high relevance, as adverse effects on the immune response could be detrimental, and even increase the risk of infection. As an example, Croes et al. [39] biofunctionalized the surface of titanium implants with chitosan-based coatings that were incorporated with different concentrations of silver nanoparticles. Despite the good antimicrobial results obtained in the in vitro tests, these coatings did not demonstrate antibacterial effects in vivo. Due to the toxicity of the silver nanoparticles on the immune cells, these coatings aggravated infection-mediated bone remodeling, including increased osteoclast formation and inflammation-induced new bone formation.
Similar results were obtained in LDPE tubes, with the bacterial clearance capacity of the HL-60 cells against S. aureus ATCC 25923 also observed, and DHA1 further potentiated this activity. The prevention of S. aureus adherence on the surface of endotracheal tubes may have potential to significantly reduce the rates of ventilator-associated pneumonia caused by these bacteria [40]. Additionally, by preventing the attachment of S. aureus, the attachment of P. aeruginosa may also be hampered, as several infection models demonstrated how early colonization by S. aureus facilitated subsequent P. aeruginosa colonization [41,42]. The development of a dual-species biofilm is expected to not only strongly worsen the pathology but significantly complicate the treatment [43].
In this study, and in concordance with our previous findings, DHA1 was identified as the best candidate to be incorporated into implantable devices. This is because, in addition to its intrinsic antibiofilm capacity against both laboratory and clinical strains of S. aureus, it also seems to be able to enhance S. aureus ATCC 25923 clearance by HL-60 cells. Further mechanistic studies should be performed in order to elucidate if DHA1 has a direct effect on the antimicrobial activity of PMNs. In the near future, we plan to assess the effects of DHA1 on phagocytosis, ROS production, and formation of neutrophil extracellular traps (NETs).
Given the promising results that DHA1 showed, both in our previous publications and in the current one, this biofilm inhibitor is involved in plans to be integrated as part of a titanium coating by means of 3D printing, with the coating formulation consisting of DHA1-loaded poly(lactic-co-glycolic acid) micro particles that suspended in a gelatine–methacrylate gel inkjet-printed onto titanium coupons [44]. The printing procedure is already validated and the prototype materials are currently being tested. In the case of endotracheal tubes, our current results suggest that all of the tested antimicrobial compounds would be beneficial for the prevention of S. aureus adhesion and subsequent biofilm formation, but DHA1 appears to be the best candidate.
To the best of our knowledge, this is one of the first studies showing a positive effect of novel antimicrobials on the antibiofilm-clearing capacity of immune cells. This is of particular relevance because it does not only provide new alternatives to fight against the immense burden of bacterial biofilms, but it also sets the basis for a new in vitro system to accelerate the drug discovery process, thereby enabling better selection of antimicrobial incorporation into medical devices.
5. Conclusions
We showed the suitability of a coculture of S. aureus and differentiated HL-60 cells as an in vitro assay to assess the applicability of antimicrobial compounds for the protection of medical devices. As a proof-of-concept, we tested three antimicrobials, concluding that the DHA derivative DHA1 is the best candidate for incorporation into implantable devices, as it does not only prevent biofilm formation on titanium but also seems to enhance the antibacterial capability of immune cells. On the other hand, according to our results, all of the antimicrobial compounds studied here, i.e., the two DHA derivatives, DHA1 and DHA2, and the flavan derivative, FLA1, can tentatively be regarded as promising candidates to form part of anti-infective endotracheal tubes.
Supplementary Materials
Supplementary materials can be found at https://www.mdpi.com/2076-2607/8/11/1757/s1.
Author Contributions
Conceptualization, I.R., A.F., and S.A.J.Z.; methodology, J.Z.P., C.G.-P., M.R., and I.R.; software, I.R.; validation, I.R. and A.F.; formal analysis, I.R.; investigation, I.R.; resources, J.Y.-K., J.Z.P., K.S., A.F., and S.A.J.Z.; data curation, I.R.; writing—original draft preparation, I.R.; writing—review and editing, A.F., M.R., S.A.J.Z., C.G.-P., and J.Y.-K.; visualization, I.R., and A.F.; supervision, A.F., K.S., and S.A.J.Z.; project administration, A.F.; funding acquisition, A.F., S.A.J.Z., and J.Y.-K. All authors read and agreed to the published version of the manuscript.
Funding
This project has received funding from the European Union’s Horizon 2020 research and innovation program under the Marie Skłodowska—Curie grant agreement No. 722467 (PRINT-AID consortium). We also acknowledge the funding received from the Jane and Aatos Erkko Foundation.
Acknowledgments
We thank Teemu J. Kinnari and Ramón Pérez-Tanoira for their support during experimentation with the clinical strain. We thank Firas Hamdan for his help obtaining the human serum.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Friedrich, A.W. Control of hospital acquired infections and antimicrobial resistance in Europe: The way to go. Wien. Med. Wochenschr. 2019, 169, 25–30. [Google Scholar] [CrossRef]
- Cassini, A.; Högberg, L.D.; Plachouras, D.; Quattrocchi, A.; Hoxha, A.; Simonsen, G.S.; Colomb-Cotinat, M.; Kretzschmar, M.E.; Devleesschauwer, B.; Cecchini, M.; et al. Attributable deaths and disability-adjusted life-years caused by infections with antibiotic-resistant bacteria in the EU and the European Economic Area in 2015: A population-level modelling analysis. Lancet Infect. Dis. 2019, 19, 56–66. [Google Scholar] [CrossRef]
- Ferri, M.; Ranucci, E.; Romagnoli, P.; Giaccone, V. Antimicrobial resistance: A global emerging threat to public health systems. Crit. Rev. Food Sci. Nutr. 2017, 57, 2857–2876. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.L.; Suleman, L.; Vuotto, C.; Donelli, G. Healthcare-associated infections, medical devices and biofilms: Risk, tolerance and control. J. Med. Microbiol. 2015, 64, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.A.; Baig, F.K.; Mehboob, R. Nosocomial infections: Epidemiology, prevention, control and surveillance. Asian Pac. J. Trop. Biomed. 2017, 7, 478–482. [Google Scholar] [CrossRef]
- Paharik, A.E.; Horswill, A.R. The Staphylococcal Biofilm: Adhesins, Regulation, and Host Response. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the Natural environment to infectious diseases. Nat. Rev. Microbiol 2004, 2, 95. [Google Scholar] [CrossRef]
- Kumar, A.; Alam, A.; Rani, M.; Ehtesham, N.Z.; Hasnain, S.E. Biofilms: Survival and defense strategy for pathogens. Int. J. Med. Microbiol. 2017, 307, 481–489. [Google Scholar] [CrossRef]
- Singh, S.; Singh, S.K.; Chowdhury, I.; Singh, R. Understanding the Mechanism of Bacterial Biofilms Resistance to Antimicrobial Agents. Open Microbiol. J. 2017, 11, 53–62. [Google Scholar] [CrossRef]
- Ferreira Tde, O.; Koto, R.Y.; Leite, G.F.; Klautau, G.B.; Nigro, S.; Silva, C.B.; Souza, A.P.; Mimica, M.J.; Cesar, R.G.; Salles, M.J. Microbial investigation of biofilms recovered from endotracheal tubes using sonication in intensive care unit pediatric patients. Braz. J. Infect. Dis. Off. Publ. Braz. Soc. Infect. Dis. 2016, 20, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Bardes, J.M.; Waters, C.; Motlagh, H.; Wilson, A. The Prevalence of Oral Flora in the Biofilm Microbiota of the Endotracheal Tube. Am. Surg. 2016, 82, 403–406. [Google Scholar] [CrossRef]
- Li, H.; Song, C.; Liu, D.; Ai, Q.; Yu, J. Molecular analysis of biofilms on the surface of neonatal endotracheal tubes based on 16S rRNA PCR-DGGE and species-specific PCR. Int. J. Clin. Exp. Med. 2015, 8, 11075–11084. [Google Scholar] [PubMed]
- Vandecandelaere, I.; Matthijs, N.; Van Nieuwerburgh, F.; Deforce, D.; Vosters, P.; De Bus, L.; Nelis, H.J.; Depuydt, P.; Coenye, T. Assessment of microbial diversity in biofilms recovered from endotracheal tubes using culture dependent and independent approaches. PLoS ONE 2012, 7, e38401. [Google Scholar] [CrossRef] [PubMed]
- Arciola, C.R.; Campoccia, D.; Montanaro, L. Implant infections: Adhesion, biofilm formation and immune evasion. Nat. Rev. Microbiol. 2018, 16, 397–409. [Google Scholar] [CrossRef]
- Kurtz, S.M.; Lau, E.; Watson, H.; Schmier, J.K.; Parvizi, J. Economic burden of periprosthetic joint infection in the United States. J. Arthroplast. 2012, 27, 61–65.e1. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, M.G.; Meaike, J.D.; Izaddoost, S.A. Orthopedic Prosthetic Infections: Diagnosis and Orthopedic Salvage. Semin. Plast. Surg. 2016, 30, 66–72. [Google Scholar] [CrossRef]
- Arciola, C.R.; Campoccia, D.; Ehrlich, G.D.; Montanaro, L. Biofilm-based implant infections in orthopaedics. Adv. Exp. Med. Biol. 2015, 830, 29–46. [Google Scholar] [CrossRef]
- Roh, J.S.; Sohn, D.H. Damage-Associated Molecular Patterns in Inflammatory Diseases. Immune Netw. 2018, 18, e27. [Google Scholar] [CrossRef]
- Franz, S.; Rammelt, S.; Scharnweber, D.; Simon, J.C. Immune responses to implants—A review of the implications for the design of immunomodulatory biomaterials. Biomaterials 2011, 32, 6692–6709. [Google Scholar] [CrossRef]
- Gristina, A.G.; Naylor, P.T.; Myrvik, Q. The Race for the Surface: Microbes, Tissue Cells, and Biomaterials. In Molecular Mechanisms of Microbial Adhesion; Springer: New York, NY, USA, 1989; pp. 177–211. [Google Scholar]
- Perez-Tanoira, R.; Han, X.; Soininen, A.; Aarnisalo, A.A.; Tiainen, V.M.; Eklund, K.K.; Esteban, J.; Kinnari, T.J. Competitive colonization of prosthetic surfaces by Staphylococcus aureus and human cells. J. Biomed. Mater. Res. Part A 2017, 105, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Stones, D.H.; Krachler, A.M. Against the tide: The role of bacterial adhesion in host colonization. Biochem. Soc. Trans. 2016, 44, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Jaskiewicz, M.; Janczura, A.; Nowicka, J.; Kamysz, W. Methods Used for the Eradication of Staphylococcal Biofilms. Antibiotics 2019, 8, 174. [Google Scholar] [CrossRef]
- Reigada, I.; Perez-Tanoira, R.; Patel, J.Z.; Savijoki, K.; Yli-Kauhaluoma, J.; Kinnari, T.J.; Fallarero, A. Strategies to Prevent Biofilm Infections on Biomaterials: Effect of Novel Naturally-Derived Biofilm Inhibitors on a Competitive Colonization Model of Titanium by Staphylococcus aureus and SaOS-2 Cells. Microorganisms 2020, 8, 345. [Google Scholar] [CrossRef] [PubMed]
- Manner, S.; Vahermo, M.; Skogman, M.E.; Krogerus, S.; Vuorela, P.M.; Yli-Kauhaluoma, J.; Fallarero, A.; Moreira, V.M. New derivatives of dehydroabietic acid target planktonic and biofilm bacteria in Staphylococcus aureus and effectively disrupt bacterial membrane integrity. Eur. J. Med. Chem. 2015, 102, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Manner, S.; Skogman, M.; Goeres, D.; Vuorela, P.; Fallarero, A. Systematic Exploration of Natural and Synthetic Flavonoids for the Inhibition of Staphylococcus aureus Biofilms. Int. J. Mol. Sci. 2013, 14, 19434–19451. [Google Scholar] [CrossRef]
- Jhunjhunwala, S. Neutrophils at the Biological–Material Interface. ACS Biomater. Sci. Eng. 2018, 4, 1128–1136. [Google Scholar] [CrossRef]
- Hochbaum, A.I.; Kolodkin-Gal, I.; Foulston, L.; Kolter, R.; Aizenberg, J.; Losick, R. Inhibitory effects of D-amino acids on Staphylococcus aureus biofilm development. J. Bacteriol. 2011, 193, 5616–5622. [Google Scholar] [CrossRef]
- Esteban, J.; Gomez-Barrena, E.; Cordero, J.; Martin-de-Hijas, N.Z.; Kinnari, T.J.; Fernandez-Roblas, R. Evaluation of quantitative analysis of cultures from sonicated retrieved orthopedic implants in diagnosis of orthopedic infection. J. Clin. Microbiol. 2008, 46, 488–492. [Google Scholar] [CrossRef]
- Collins, S.J.; Ruscetti, F.W.; Gallagher, R.E.; Gallo, R.C. Terminal differentiation of human promyelocytic leukemia cells induced by dimethyl sulfoxide and other polar compounds. Proc. Natl. Acad. Sci. USA 1978, 75, 2458–2462. [Google Scholar] [CrossRef]
- Lu, T.; Porter, A.R.; Kennedy, A.D.; Kobayashi, S.D.; DeLeo, F.R. Phagocytosis and killing of Staphylococcus aureus by human neutrophils. J. Innate Immun. 2014, 6, 639–649. [Google Scholar] [CrossRef]
- van Kessel, K.P.M.; Bestebroer, J.; van Strijp, J.A.G. Neutrophil-Mediated Phagocytosis of Staphylococcus aureus. Front. Immunol. 2014, 5, 467. [Google Scholar] [CrossRef] [PubMed]
- Hiltunen, A.K.; Savijoki, K.; Nyman, T.A.; Miettinen, I.; Ihalainen, P.; Peltonen, J.; Fallarero, A. Structural and Functional Dynamics of Staphylococcus aureus Biofilms and Biofilm Matrix Proteins on Different Clinical Materials. Microorganisms 2019, 7, 584. [Google Scholar] [CrossRef]
- Sunzel, B.; Söderberg, T.A.; Reuterving, C.O.; Hallmans, G.; Holm, S.E.; Hänström, L. Neutralizing effect of zinc oxide on dehydroabietic acid-induced toxicity on human polymorphonuclear leukocytes. Biol. Trace Elem. 1991, 30, 257–266. [Google Scholar] [CrossRef]
- Czerwińska, M.E.; Dudek, M.K.; Pawłowska, K.A.; Pruś, A.; Ziaja, M.; Granica, S. The influence of procyanidins isolated from small-leaved lime flowers (Tilia cordata Mill.) on human neutrophils. Fitoterapia 2018, 127, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Pawłowska, K.A.; Hałasa, R.; Dudek, M.K.; Majdan, M.; Jankowska, K.; Granica, S. Antibacterial and anti-inflammatory activity of bistort (Bistorta officinalis) aqueous extract and its major components. Justification of the usage of the medicinal plant material as a traditional topical agent. J. Ethnopharmacol. 2020, 260, 113077. [Google Scholar] [CrossRef]
- Manda-Handzlik, A.; Bystrzycka, W.; Wachowska, M.; Sieczkowska, S.; Stelmaszczyk-Emmel, A.; Demkow, U.; Ciepiela, O. The influence of agents differentiating HL-60 cells toward granulocyte-like cells on their ability to release neutrophil extracellular traps. Immunol. Cell Biol. 2018, 96, 413–425. [Google Scholar] [CrossRef]
- Yaseen, R.; Blodkamp, S.; Luthje, P.; Reuner, F.; Vollger, L.; Naim, H.Y.; von Kockritz-Blickwede, M. Antimicrobial activity of HL-60 cells compared to primary blood-derived neutrophils against Staphylococcus aureus. J. Negat. Results Biomed. 2017, 16, 7. [Google Scholar] [CrossRef] [PubMed]
- Croes, M.; Bakhshandeh, S.; van Hengel, I.A.J.; Lietaert, K.; van Kessel, K.P.M.; Pouran, B.; van der Wal, B.C.H.; Vogely, H.C.; Van Hecke, W.; Fluit, A.C.; et al. Antibacterial and immunogenic behavior of silver coatings on additively manufactured porous titanium. Acta Biomater. 2018, 81, 315–327. [Google Scholar] [CrossRef]
- Seitz, A.P.; Schumacher, F.; Baker, J.; Soddemann, M.; Wilker, B.; Caldwell, C.C.; Gobble, R.M.; Kamler, M.; Becker, K.A.; Beck, S.; et al. Sphingosine-coating of plastic surfaces prevents ventilator-associated pneumonia. J. Mol. Med. 2019, 97, 1195–1211. [Google Scholar] [CrossRef]
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Lung infections associated with cystic fibrosis. Clin. Microbiol. Rev. 2002, 15, 194–222. [Google Scholar] [CrossRef]
- Alves, P.M.; Al-Badi, E.; Withycombe, C.; Jones, P.M.; Purdy, K.J.; Maddocks, S.E. Interaction between Staphylococcus aureus and Pseudomonas aeruginosa is beneficial for colonisation and pathogenicity in a mixed biofilm. Pathog. Dis. 2018, 76. [Google Scholar] [CrossRef]
- Beaudoin, T.; Yau, Y.C.W.; Stapleton, P.J.; Gong, Y.; Wang, P.W.; Guttman, D.S.; Waters, V. Staphylococcus aureus interaction with Pseudomonas aeruginosa biofilm enhances tobramycin resistance. NPJ Biofilms Microbiomes 2017, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chu, L.; Yang, S.; Zhang, H.; Qin, L.; Guillaume, O.; Eglin, D.; Richards, R.G.; Tang, T. Dual-functional 3D-printed composite scaffold for inhibiting bacterial infection and promoting bone regeneration in infected bone defect models. Acta Biomater. 2018, 79, 265–275. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).