Disinfectant, Soap or Probiotic Cleaning? Surface Microbiome Diversity and Biofilm Competitive Exclusion


Abstract
1. Introduction
2. Materials and Methods
2.1. Strain Acquisition, Characterization and Maintenance
2.2. Cleaning Regime and Surface Selection
2.3. Cleaning Regime Impact on Community Dynamics
2.3.1. Resident Surface Community Quantification
2.3.2. Resident Surface Community Diversity
2.3.3. Competitive Exclusion: Cell Survival
2.3.4. Competitive Exclusion: Biofilm Proliferation
2.4. Statistical Analyses
3. Results
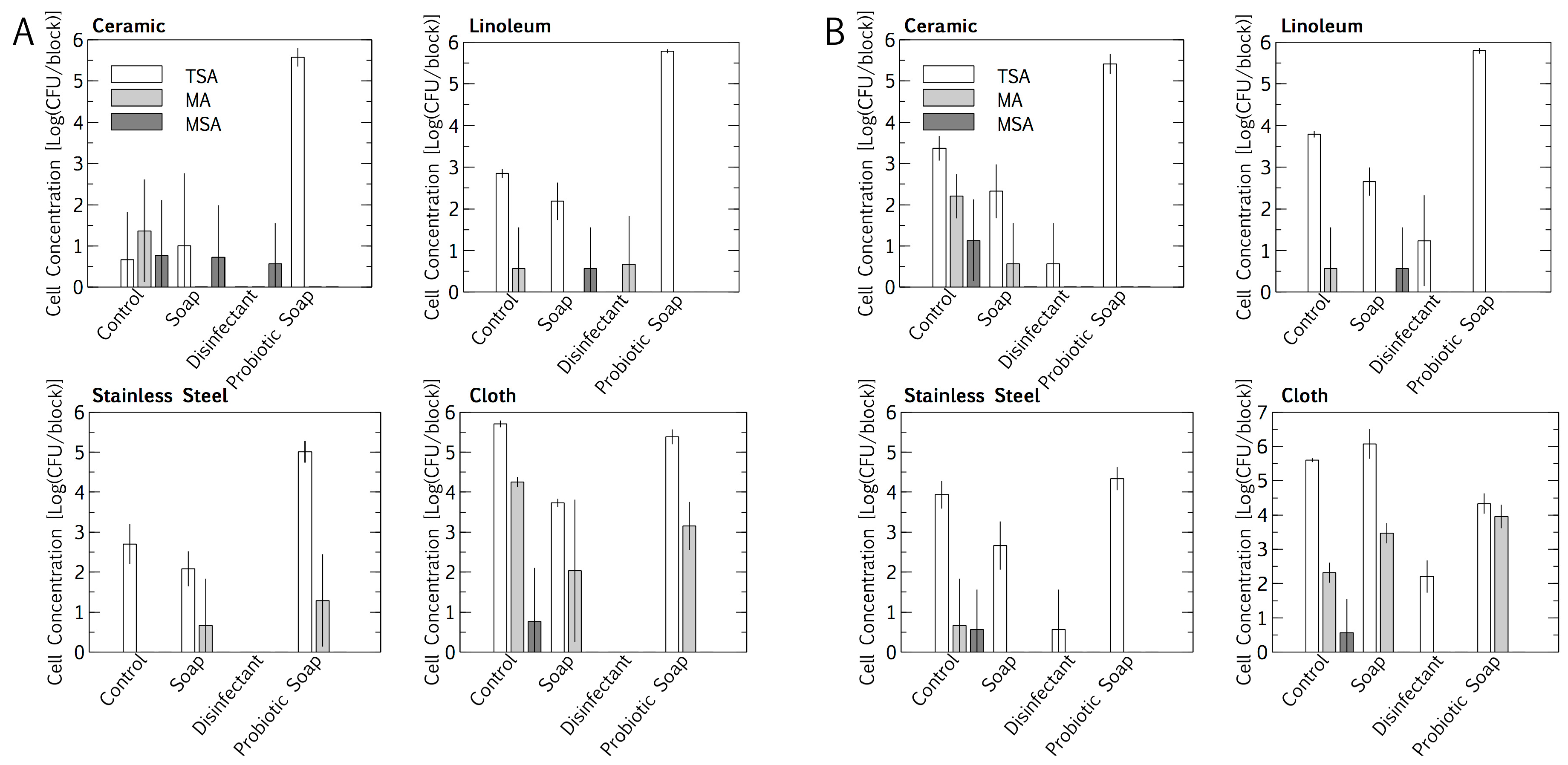
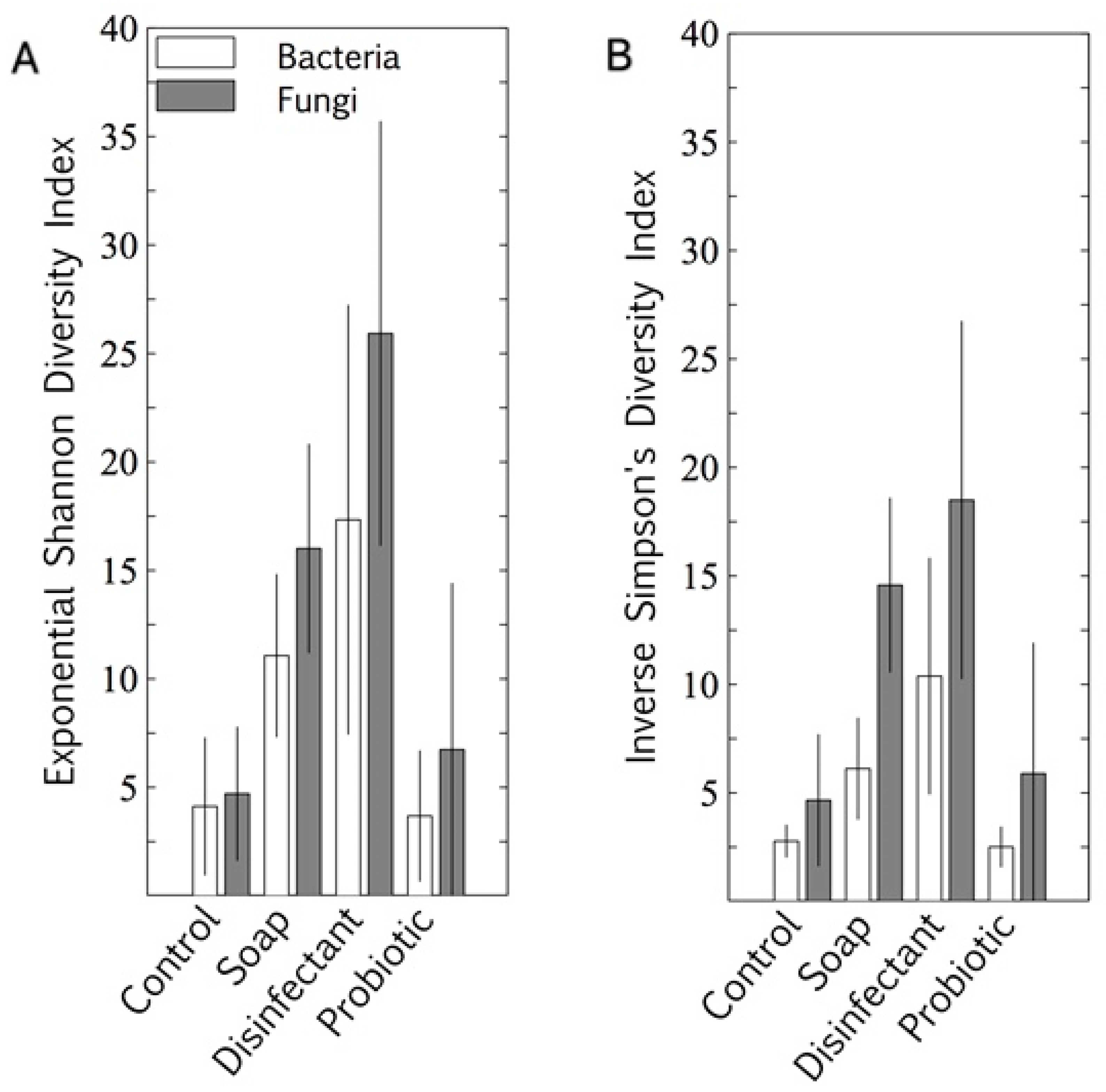
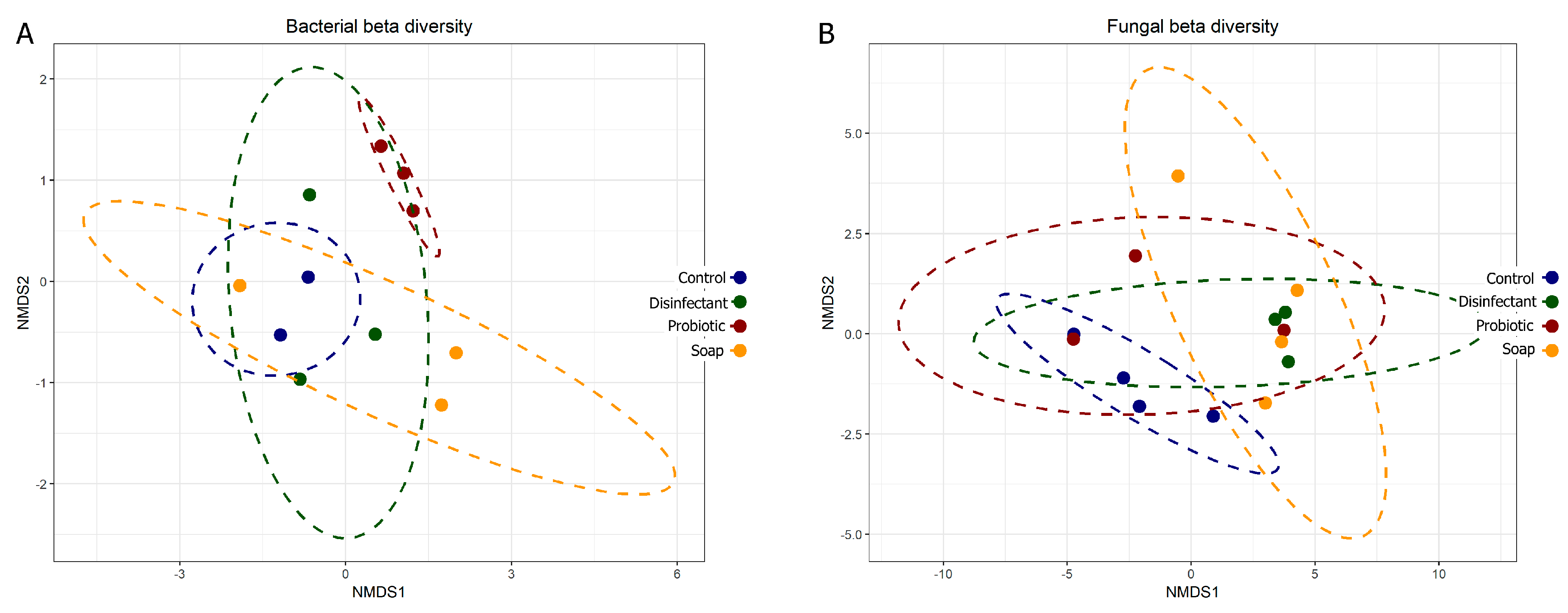
3.1. Resident Populations: Impact of Cleaning Regimes
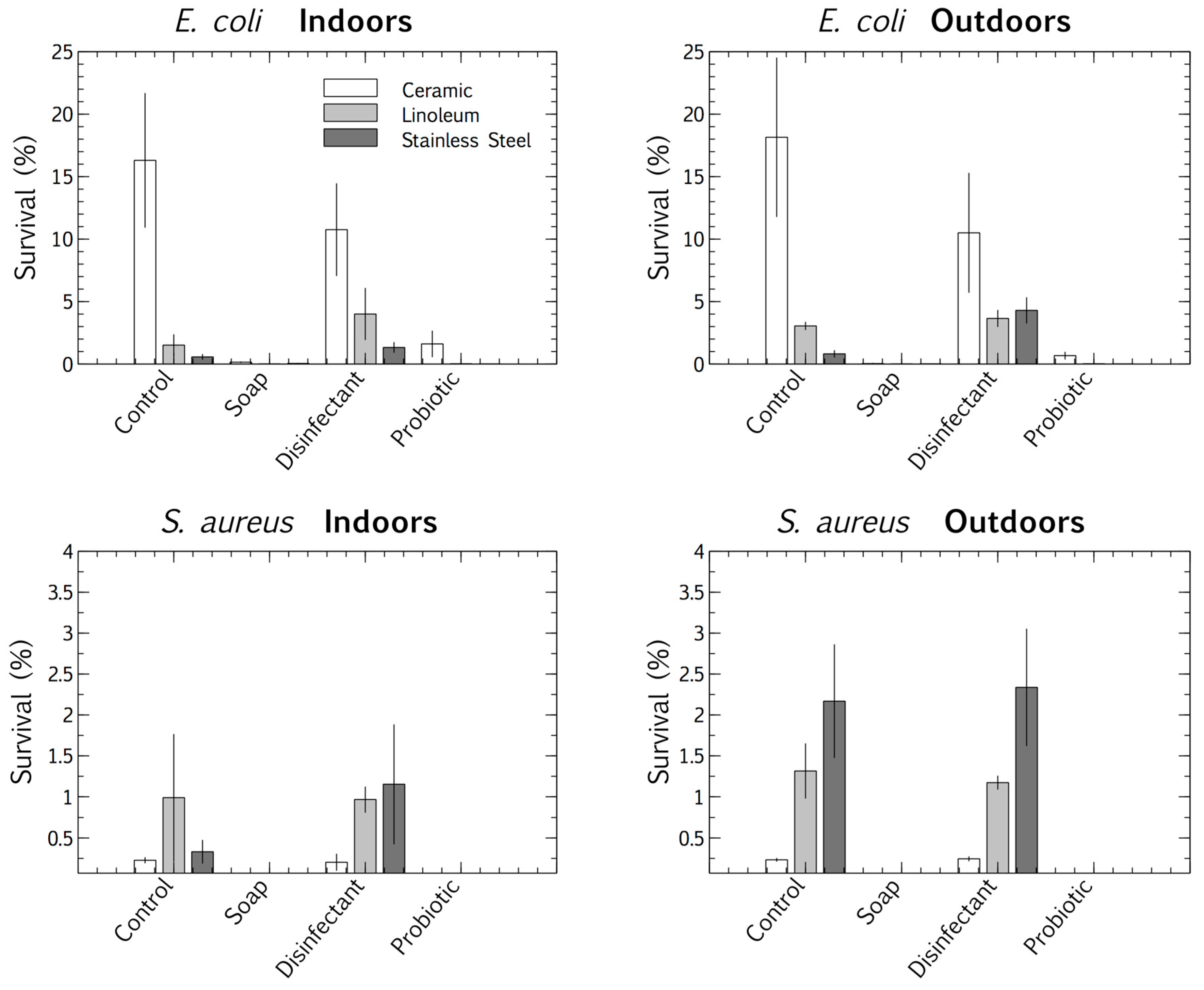
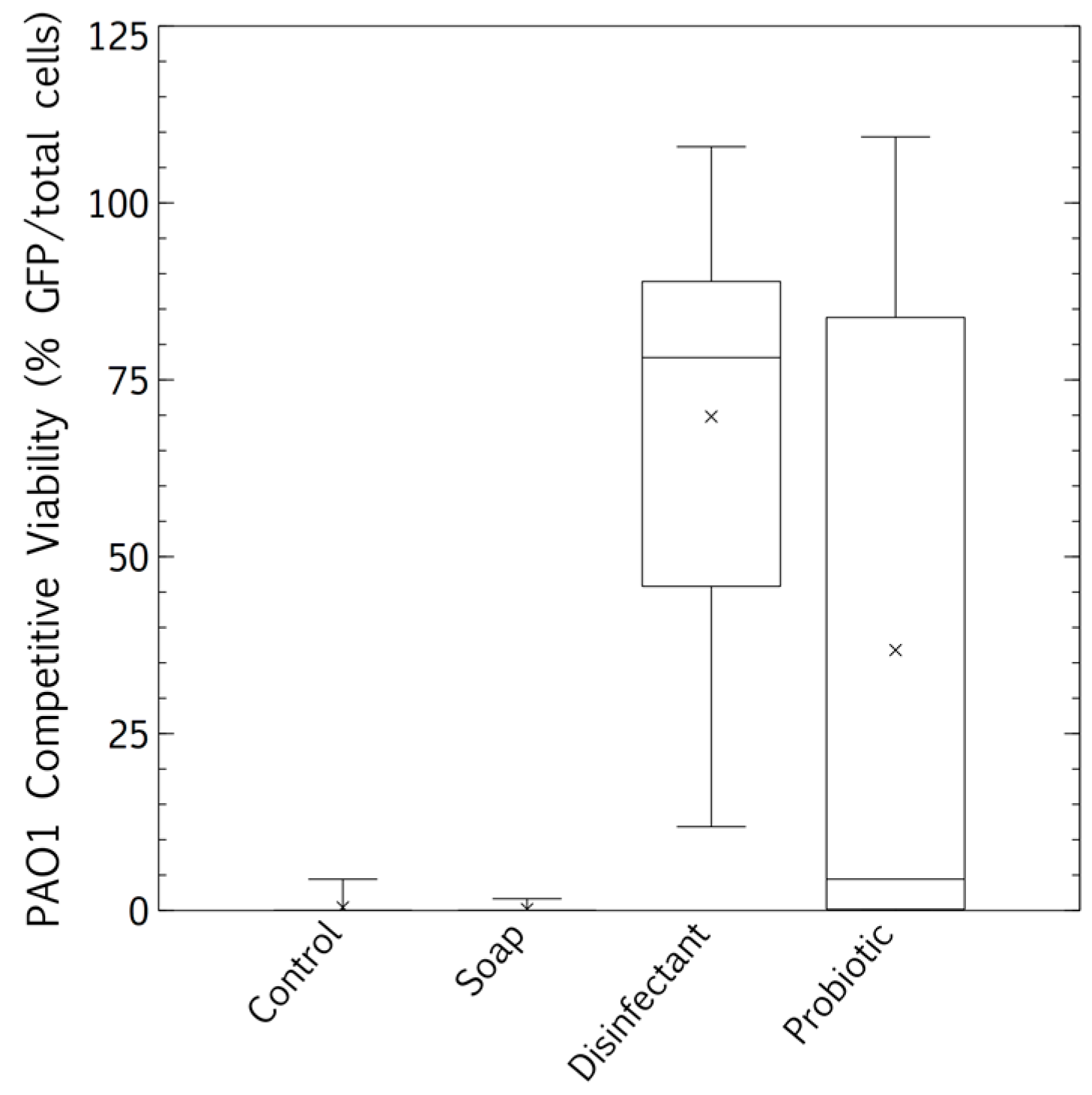
3.2. Pathogen Persistence: Impact of Cleaning Regimes
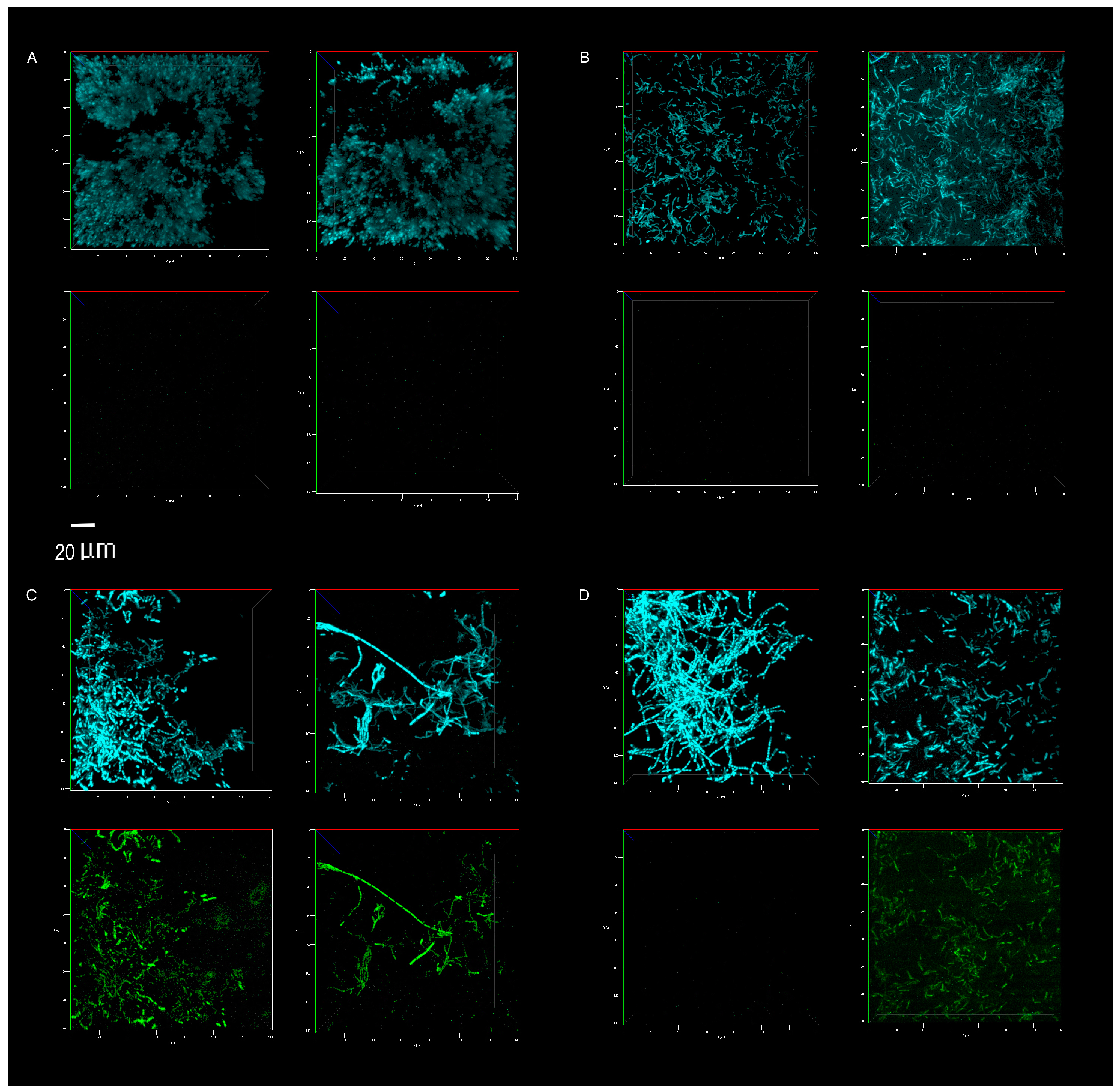
3.3. Biofilm Proliferation: Impact of Cleaning Regimes
4. Discussion
4.1. Placement and Materials
4.2. Resident Microbiome
4.3. Competitive Exclusion: Survival and Biofilm Growth
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. Methods 1. ARISA Sequencing and Data Processing
Appendix A.2. Methods 2. Confocal Microscopy Imaging and Image Processing
Appendix B. Surface and Microbiome Visualization: Scanning Electron Microscopy
Appendix B.1. Materials and Methods
Appendix B.2. Results

References
- Lax, S.; Gilbert, J.A. Hospital-associated microbiota and implications for nosocomial infections. Trends Mol. Med. 2015, 21, 427–432. [Google Scholar] [CrossRef]
- Weinstein, R.A.; Hota, B. Contamination, Disinfection, and Cross-Colonization: Are Hospital Surfaces Reservoirs for Nosocomial Infection? Clin. Infect. Dis. 2004, 39, 1182–1189. [Google Scholar] [CrossRef] [PubMed]
- Vandini, A.; Temmerman, R.; Frabetti, A.; Caselli, E.; Antonioli, P.; Balboni, P.G.; Mazzacane, S. Hard surface biocontrol in hospitals using microbial-based cleaning products. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Mann, E.E.; Manna, D.; Mettetal, M.R.; May, R.M.; Dannemiller, E.M.; Chung, K.K.; Reddy, S.T. Surface micropattern limits bacterial contamination. Antimicrob. Resist. Infect. Control. 2014, 3, 28. [Google Scholar] [CrossRef]
- Mora, M.; Mahnert, A.; Koskinen, K.; Pausan, M.R.; Oberauner-Wappis, L.; Krause, R.; Moissl-Eichinger, C. Microorganisms in confined habitats: Microbial monitoring and control of intensive care units, operating rooms, cleanrooms and the international space station. Front. Microbiol. 2016. [Google Scholar] [CrossRef]
- Wand, M.E.; Bock, L.J.; Bonney, L.C.; Sutton, J.M. Mechanisms of increased resistance to chlorhexidine and cross-resistance to colistin following exposure of Klebsiella pneumoniae clinical isolates to chlorhexidine. Antimicrob. Agents Chemother. 2017, 61, e01162-16. [Google Scholar] [CrossRef] [PubMed]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Salamat, M.K.F. Infection and Drug Resistance Dovepress Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018. [Google Scholar] [CrossRef]
- World Health Organization. Health Care-Associated Infections Fact Sheet. 2017. Available online: http://www.who.int/gpsc/country_work/gpsc_ccisc_fact_sheet_en.pdf (accessed on 12 April 2018).
- D’Accolti, M.; Soffritti, I.; Mazzacane, S.; Caselli, E. Fighting AMR in the healthcare environment: Microbiome-based sanitation approaches and monitoring tools. Int. J. Mol. Sci. 2019, 20, 1535. [Google Scholar] [CrossRef] [PubMed]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277. [Google Scholar]
- Tezel, U.; Pavlostathis, S.G. Quaternary ammonium disinfectants: Microbial adaptation, degradation and ecology. Curr. Opin. Biotechnol. 2015, 33, 296–304. [Google Scholar] [CrossRef]
- Dunne, S.S.; Ahonen, M.; Modic, M.; Crijns, F.R.; Keinänen-Toivola, M.M.; Meinke, R.; Dunne, C.P. Specialized cleaning associated with antimicrobial coatings for reduction of hospital-acquired infection: Opinion of the COST Action Network AMiCI (CA15114). J. Hosp. Infect. 2018, 99, 250–255. [Google Scholar] [CrossRef]
- Abt, M.C.; Pamer, E.G. Commensal bacteria mediated defenses against pathogens. Curr. Opin. Immunol. 2014, 16–22. [Google Scholar] [CrossRef]
- Hibbing, M.E.; Fuqua, C.; Parsek, M.R.; Peterson, S.B. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 15–25. [Google Scholar] [CrossRef]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Smid, E.J. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotech. 2017, 44, 94–102. [Google Scholar] [CrossRef]
- Finlay, B.B.; Arrieta, M.C. Let Them Eat Dirt: Saving Your Child from an Oversanitized World; Algonquin Books: New York, NY, USA, 2016. [Google Scholar]
- Sha, S.; Liang, J.; Chen, M.; Xu, B.; Liang, C.; Wei, N.; Wu, K. Systematic review: Faecal microbiota transplantation therapy for digestive and nondigestive disorders in adults and children. Aliment. Pharm. Therap. 2014, 39, 1003–1032. [Google Scholar] [CrossRef]
- Falagas, M.E.; Makris, G.C. Probiotic bacteria and biosurfactants for nosocomial infection control: A hypothesis. J. Hosp. Infect. 2009, 71, 301–306. [Google Scholar] [CrossRef]
- Kembel, S.W.; Jones, E.; Kline, J.; Northcutt, D.; Stenson, J.; Womack, A.M.; Green, J.L. Architectural design influences the diversity and structure of the built environment microbiome. ISME J. 2012, 6, 1469–1479. [Google Scholar] [CrossRef]
- Berg, G.; Mahnert, A.; Moissl-Eichinger, C. Beneficial effects of plant-associated microbes on indoor microbiomes and human health? Front. Microbiol. 2014. [Google Scholar] [CrossRef]
- Available online: http://www.healthierhospitals.org/ (accessed on 6 August 2019).
- Available online: https://noharm.org/ (accessed on 6 August 2019).
- Available online: https://practicegreenhealth.org/ (accessed on 6 August 2019).
- Parat, S.; Perdrix, A.; Fricker-Hidalgo, H.; Saude, I.; Grillot, R.; Baconnier, P. Multivariate analysis comparing microbial air content of an air-conditioned building and a naturally ventilated building over one year. Atmos. Environ. 1997, 31, 441–449. [Google Scholar] [CrossRef]
- Harrison, J.; Pickering, C.A.C.; Faragher, E.B.; Austwick, P.K.C.; Little, S.A.; Lawton, L. An investigation of the relationship between microbial and particulate indoor air pollution and the sick building syndrome. Resp. Med. 1992, 86, 225–235. [Google Scholar] [CrossRef]
- Ronan, E.; Yeung, C.W.; Hausner, M.; Wolfaardt, G.M. Interspecies interaction extends bacterial survival at solid-air interfaces. Biofouling 2013, 29, 1087–1096. [Google Scholar] [CrossRef]
- Cardinale, M.; Brusetti, L.; Quatrini, P.; Borin, S.; Puglia, A.M.; Rizzi, A.; Daffonchio, D. Comparison of different primer sets for use in automated ribosomal intergenic spacer analysis of complex bacterial communities. Appl. Environ. Microbiol. 2004, 70, 6147–6156. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press Inc.: New York, NY, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- De Waal, G.M.; Engelbrecht, L.; Davis, T.; De Villiers, W.J.; Kell, D.B.; Pretorius, E. Correlative Light-Electron Microscopy detects lipopolysaccharide and its association with fibrin fibres in Parkinson’s Disease, Alzheimer’s Disease and Type 2 Diabetes Mellitus. Sci. Rep. 2018, 8, 16798. [Google Scholar] [CrossRef]
- La Duc, M.T.; Dekas, A.; Osman, S.; Moissl, C.; Newcombe, D.; Venkateswaran, K. Isolation and characterization of bacteria capable of tolerating the extreme conditions of clean room environments. Appl. Environ. Microbiol. 2007, 73, 2600–2611. [Google Scholar] [CrossRef]
- Palmer, J.; Flint, S.; Brooks, J. Bacterial cell attachment, the beginning of a biofilm. J. Ind. Microbiol. Biotechnol. 2007, 34, 577–588. [Google Scholar] [CrossRef]
- Morgan, T.D.; Wilson, M. The effects of surface roughness and type of denture acrylic on biofilm formation by Streptococcus oralis in a constant depth film fermentor. J. Appl. Microbiol. 2001, 91, 47–53. [Google Scholar] [CrossRef]
- Gupta, M.; Bisesi, M.; Lee, J. Comparison of survivability of Staphylococcus aureus and spores of Aspergillus niger on commonly used floor materials. Am. J. Infect. Control 2017, 45, 717–722. [Google Scholar] [CrossRef]
- Bergen, L.K.; Meyer, M.; Høg, M.; Rubenhagen, B.; Andersen, L.P. Spread of bacteria on surfaces when cleaning with microfibre cloths. J. Hosp. Infect. 2009, 71, 132–137. [Google Scholar] [CrossRef]
- Moeller, R.; Stackebrandt, E.; Reitz, G.; Berger, T.; Rettberg, P.; Doherty, A.J.; Nicholson, W.L. Role of DNA repair by nonhomologous-end joining in Bacillus subtilis spore resistance to extreme dryness, mono-and polychromatic UV, and ionizing radiation. J. Bacteriol. 2007, 189, 3306–3311. [Google Scholar] [CrossRef] [PubMed]
- Baron, J.L.; Vikram, A.; Duda, S.; Stout, J.E.; Bibby, K. Shift in the microbial ecology of a hospital hot water system following the introduction of an on-site monochloramine disinfection system. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Pourhajibagher, M.; Bahador, A. An in vivo evaluation of microbial diversity before and after the photo-activated disinfection in primary endodontic infections: Traditional phenotypic and molecular approaches. Photodiag. Photodyn. Ther. 2018, 22, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Tringe, S.G.; Zhang, T.; Liu, X.; Yu, Y.; Lee, W.H.; Yap, J.; Rohwer, F. The airborne metagenome in an indoor urban environment. PLoS ONE 2008, 3. [Google Scholar] [CrossRef]
- Dillon, R.J.; Vennard, C.T.; Buckling, A.; Charnley, A.K. Diversity of locust gut bacteria protects against pathogen invasion. Ecol. Lett. 2005, 8, 1291–1298. [Google Scholar] [CrossRef]
- Available online: https://countercultureclean.com/products/baby-got-bac−2-pack (accessed on 6 August 2019).
- Available online: https://www.probioclean.com/faq/ (accessed on 6 August 2019).
- Available online: https://mrsmartins.co.za/product/mrs-martins-probiotic-surface-soap−500mL-aluminium/ (accessed on 6 August 2019).
- Available online: https://patents.google.com/patent/CA2983075A1/en (accessed on 6 August 2019).
- Available online: https://www.probac.co.za/pages/the-probac-story (accessed on 6 August 2019).
- Caselli, E.; Brusaferro, S.; Coccagna, M.; Arnoldo, L.; Berloco, F.; Antonioli, P.; Conte, A. Reducing healthcare-associated infections incidence by a probiotic-based sanitation system: A multicentre, prospective, intervention study. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Caselli, E.; Antonioli, P.; Mazzacane, S. Safety of probiotics used for hospital environmental sanitation. J. Hosp. Infect. 2016, 94, 193. [Google Scholar] [CrossRef]
- Caselli, E.; Arnoldo, L.; Rognoni, C.; D’Accolti, M.; Soffritti, I.; Lanzoni, L.; Mazzacane, S. Impact of a probiotic-based hospital sanitation on antimicrobial resistance and HAI-associated antimicrobial consumption and costs: A multicenter study. Dovepress 2019, 12, 501–510. [Google Scholar] [CrossRef]
- Joubert, L.M.; Ferreira, J.A.; Stevens, D.A.; Nazik, H.; Cegelski, L. Visualization of Aspergillus fumigatus biofilms with Scanning Electron Microscopy and Variable Pressure-Scanning Electron Microscopy: A comparison of processing techniques. J. Microbiol. Methods. 2017, 132, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Slabbert, E.; Van Heerden, C.J.; Jacobs, K. Optimisation of automated ribosomal intergenic spacer analysis for the estimation of microbial diversity in fynbos soil. S. Afr. J. Sci. 2010, 106, 1–4. [Google Scholar] [CrossRef][Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 10 December 2019).
- Available online: https://imagej.nih.gov/ij/ (accessed on 10 December 2019).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Product | Dilution | Procedure |
|---|---|---|---|
| Control | Tap water (Cold tap) | - | Rinse cloth, wring well, wipe once. |
| Plain soap | EarthSap biodegradable soap. Active ingredients: saponified vegetable extract, essential oils, natural gum | 250 mL·500 mL−1 | Wipe with cloth soaked in soap, rinse cloth thoroughly, wipe (×3). |
| Disinfectant | Jik bleach. Active ingredient: 3.5% m/v sodium hypochlorite | 50 mL·500 mL−1 | Wipe with disinfectant, soak for 5 min. Rinse cloth thoroughly, wipe (×3). |
| Probiotic cleaner | Bacterrorist non-toxic all-purpose cleaner (8.6 × 107 colony forming units (cfu)·mL−1 Bacillus spores, analyzed in-study) | Undiluted | Spray directly onto surface, wipe with damp cloth. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stone, W.; Tolmay, J.; Tucker, K.; Wolfaardt, G.M. Disinfectant, Soap or Probiotic Cleaning? Surface Microbiome Diversity and Biofilm Competitive Exclusion. Microorganisms 2020, 8, 1726. https://doi.org/10.3390/microorganisms8111726
Stone W, Tolmay J, Tucker K, Wolfaardt GM. Disinfectant, Soap or Probiotic Cleaning? Surface Microbiome Diversity and Biofilm Competitive Exclusion. Microorganisms. 2020; 8(11):1726. https://doi.org/10.3390/microorganisms8111726
Chicago/Turabian StyleStone, Wendy, Janke Tolmay, Keira Tucker, and Gideon M. Wolfaardt. 2020. "Disinfectant, Soap or Probiotic Cleaning? Surface Microbiome Diversity and Biofilm Competitive Exclusion" Microorganisms 8, no. 11: 1726. https://doi.org/10.3390/microorganisms8111726
APA StyleStone, W., Tolmay, J., Tucker, K., & Wolfaardt, G. M. (2020). Disinfectant, Soap or Probiotic Cleaning? Surface Microbiome Diversity and Biofilm Competitive Exclusion. Microorganisms, 8(11), 1726. https://doi.org/10.3390/microorganisms8111726