Defining Coral Bleaching as a Microbial Dysbiosis within the Coral Holobiont


 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
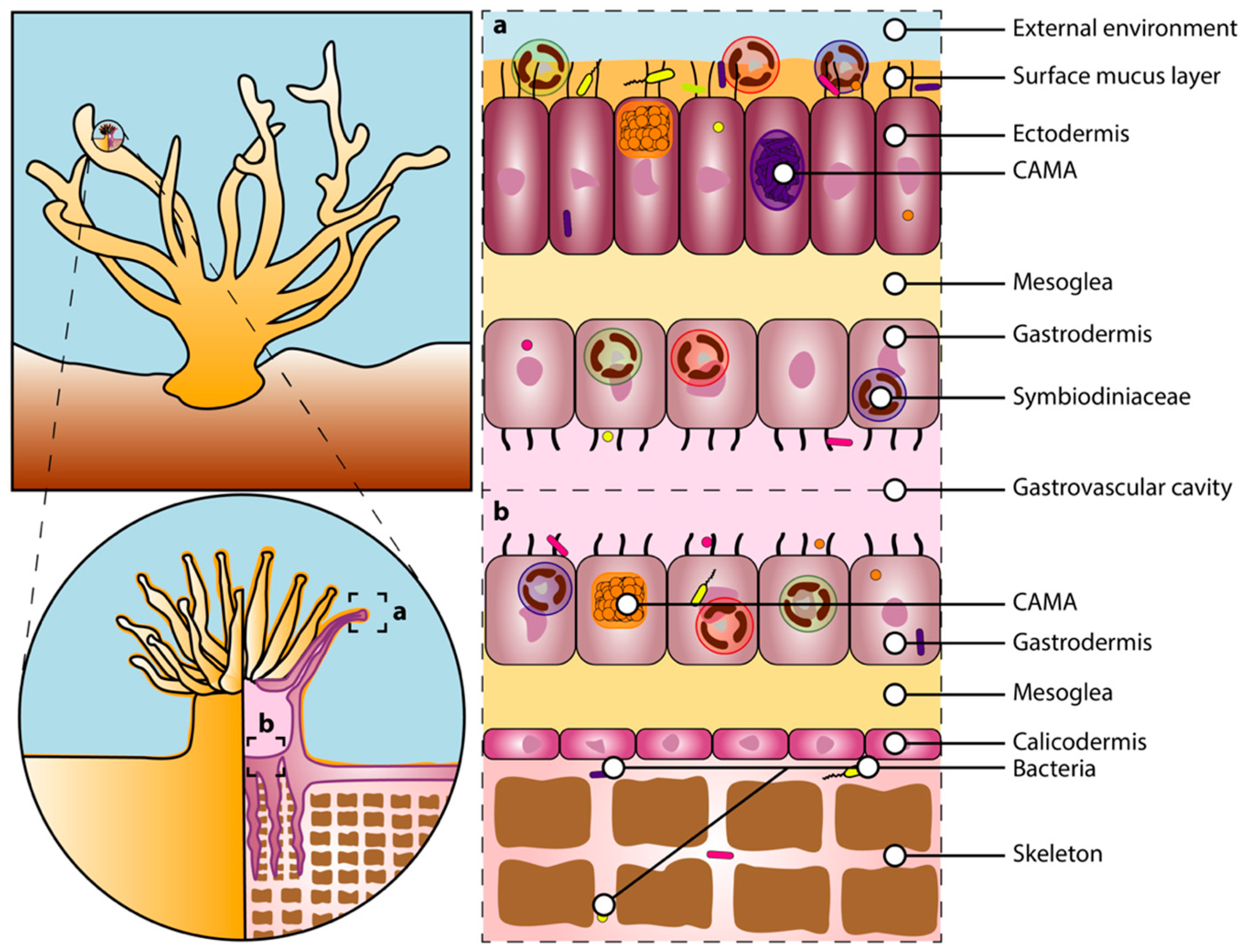
2. The Coral Holobiont
2.1. Corals
2.2. Symbiodiniaceae
2.3. The Microbiota
2.3.1. Bacteria
2.3.2. Viruses
2.3.3. Other Microbial Members
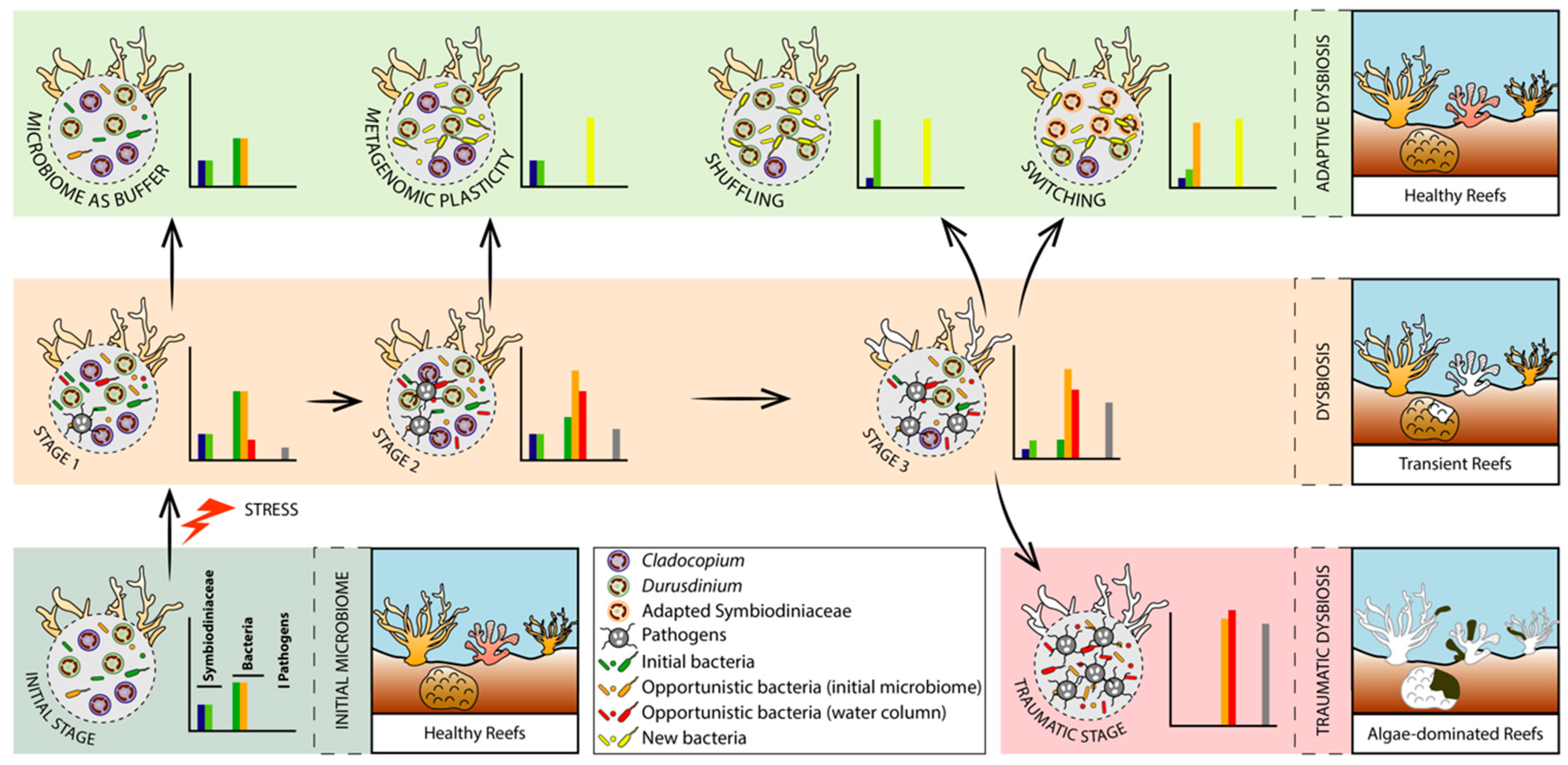
3. Coral Bleaching: From Adaptive to Traumatic Holobiont Dysbiosis
- (1)
- Adaptive dysbiosis without Symbiodiniaceae community restructuring;
- (2)
- Adaptive dysbiosis with Symbiodiniaceae community restructuring with or without transient bleaching;
- (3)
- Maladaptive/traumatic dysbiosis with irreversible loss of Symbiodiniaceae and invasion of opportunistic microbes leading to holobiont death.
3.1. Adaptive Dysbiosis without Symbiodiniaceae Community Restructuring
3.2. Adaptive Dysbiosis with Symbiodiniaceae Community Restructuring with or without Transient Bleaching
3.3. Madaptive/Traumatic Dysbiosis with loss of Symbiodiniaceae and Invasion of Opportunistic Microbes Leading to Holobiont Death
4. Microbiome Manipulations
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Moberg, F.; Folke, C. Ecological goods and services of coral reef ecosystems. Ecol. Econ. 1999, 29, 215–233. [Google Scholar] [CrossRef]
- Spalding, M.; Spalding, M.D.; Ravilious, C.; Green, E.P. World Atlas of Coral Reefs; University of California Press: Berkeley, CA, USA, 2001. [Google Scholar]
- Chen, P.-Y.; Chen, C.-C.; Chu, L.; McCarl, B. Evaluating the economic damage of climate change on global coral reefs. Glob. Environ. Chang. 2015, 30, 12–20. [Google Scholar] [CrossRef]
- Odum, H.T.; Odum, E.P. Trophic structure and productivity of a windward coral reef community on Eniwetok Atoll. Ecol. Monogr. 1955, 25, 291–320. [Google Scholar] [CrossRef]
- Hatcher, B.G. Coral reef primary productivity. A hierarchy of pattern and process. Trends Ecol. Evol. 1990, 5, 149–155. [Google Scholar] [CrossRef]
- Atkinson, M.; Falter, J.; Black, K.; Shimmield, G. Biogeochemistry of marine systems. In Coral Reefs; Blackwell: Oxford, UK, 2003; pp. 40–64. [Google Scholar]
- Mcallister, D.E. What is the status of the world’s coral reef fishes. Sea Wind 1991, 5, 14–18. [Google Scholar]
- Knowlton, N.; Brainard, R.E.; Fisher, R.; Moews, M.; Plaisance, L.; Caley, M.J. Coral reef biodiversity. In Life in the World’s Oceans: Diversity Distribution and Abundance; Wiley-Blackwell: Chichester, UK, 2010; pp. 65–74. [Google Scholar]
- White, A.T.; Vogt, H.P.; Arin, T. Philippine coral reefs under threat: The economic losses caused by reef destruction. Mar. Pollut. Bull. 2000, 40, 598–605. [Google Scholar] [CrossRef]
- UN Environment; ISU; ICRI; Trucost. The Coral Reef Economy: The Business Case for Investment in the Protection, Preservation and Enhancement of Coral Reef Health. 2018; 36p, Available online: https://wedocs.unep.org/handle/20.500.11822/26694 (accessed on 18 October 2020).
- Spalding, M.; Burke, L.; Wood, S.A.; Ashpole, J.; Hutchison, J.; Zu Ermgassen, P. Mapping the global value and distribution of coral reef tourism. Mar. Policy 2017, 82, 104–113. [Google Scholar] [CrossRef]
- Ferrario, F.; Beck, M.W.; Storlazzi, C.D.; Micheli, F.; Shepard, C.C.; Airoldi, L. The effectiveness of coral reefs for coastal hazard risk reduction and adaptation. Nat. Commun. 2014, 5, 3794. [Google Scholar] [CrossRef]
- Sipkema, D.; Franssen, M.C.; Osinga, R.; Tramper, J.; Wijffels, R.H. Marine sponges as pharmacy. Mar. Biotechnol. 2005, 7, 142. [Google Scholar] [CrossRef]
- Senthilkumar, K.; Ramajayam, G.; Venkatesan, J.; Kim, S.-K.; Ahn, B.-C. Marine sponge-derived antiangiogenic compounds for cancer therapeutics. In Marine Sponges: Chemicobiological and Biomedical Applications; Springer: New Delhi, India, 2016; pp. 305–314. [Google Scholar]
- do Nascimento-Neto, L.G.; Cabral, M.G.; Carneiro, R.F.; Silva, Z.; Arruda, F.V.; Nagano, C.S.; Fernandes, A.R.; Sampaio, A.H.; Teixeira, E.H.; Videira, P.A. Halilectin-3, a lectin from the marine sponge Haliclona caerulea, induces apoptosis and autophagy in human breast cancer MCF7 cells through caspase-9 pathway and LC3-II protein expression. Anti-Cancer Agents Med. Chem. (Former. Curr. Med. Chem. Anti-Cancer Agents) 2018, 18, 521–528. [Google Scholar] [CrossRef]
- Graham, N.; Nash, K. The importance of structural complexity in coral reef ecosystems. Coral Reefs 2013, 32, 315–326. [Google Scholar] [CrossRef]
- Wild, C.; Huettel, M.; Klueter, A.; Kremb, S.G.; Rasheed, M.Y.; Jørgensen, B.B. Coral mucus functions as an energy carrier and particle trap in the reef ecosystem. Nature 2004, 428, 66–70. [Google Scholar] [CrossRef] [PubMed]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbionts. Curr. Biol. 2018, 28, 2570–2580. [Google Scholar] [CrossRef] [PubMed]
- Allemand, D.; Furla, P. How does an animal behave like a plant? Physiological and molecular adaptations of zooxanthellae and their hosts to symbiosis. C. R. Biol. 2018, 341, 276–280. [Google Scholar] [CrossRef]
- Kimes, N.E.; Van Nostrand, J.D.; Weil, E.; Zhou, J.; Morris, P.J. Microbial functional structure of Montastraea faveolata, an important caribbean reef-building coral, differs between healthy and yellow-band diseased colonies. Environ. Microbiol. 2010, 12, 541–556. [Google Scholar] [CrossRef]
- Rohwer, F.; Seguritan, V.; Azam, F.; Knowlton, N. Diversity and distribution of coral-associated bacteria. Mar. Ecol. Prog. Ser. 2002, 243, 1–10. [Google Scholar] [CrossRef]
- Rosenberg, E.; Koren, O.; Reshef, L.; Efrony, R.; Zilber-Rosenberg, I. The role of microorganisms in coral health, disease and evolution. Nat. Rev. Microbiol. 2007, 5, 355. [Google Scholar] [CrossRef]
- Boulotte, N.M.; Dalton, S.J.; Carroll, A.G.; Harrison, P.L.; Putnam, H.M.; Peplow, L.M.; Van Oppen, M.J. Exploring the Symbiodinium rare biosphere provides evidence for symbiont switching in reef-building corals. ISME J. 2016, 10, 2693–2701. [Google Scholar] [CrossRef]
- Muscatine, L.; Porter, J.; Kaplan, I. Resource partitioning by reef corals as determined from stable isotope composition. Mar. Biol. 1989, 100, 185–193. [Google Scholar] [CrossRef]
- Davy, S.K.; Allemand, D.; Weis, V.M. Cell biology of cnidarian-dinoflagellate symbiosis. Microbiol. Mol. Biol. Rev. 2012, 76, 229–261. [Google Scholar] [CrossRef]
- Lema, K.A.; Willis, B.L.; Bourne, D.G. Corals form characteristic associations with symbiotic nitrogen-fixing bacteria. Appl. Environ. Microbiol. 2012, 78, 3136–3144. [Google Scholar] [CrossRef] [PubMed]
- Rädecker, N.; Pogoreutz, C.; Voolstra, C.R.; Wiedenmann, J.; Wild, C. Nitrogen cycling in corals: The key to understanding holobiont functioning? Trends Microbiol. 2015, 23, 490–497. [Google Scholar] [CrossRef]
- González, J.M.; Kiene, R.P.; Moran, M.A. Transformation of sulfur compounds by an abundant lineage of marine bacteria in the α-subclass of the class proteobacteria. Appl. Environ. Microbiol. 1999, 65, 3810–3819. [Google Scholar] [CrossRef] [PubMed]
- Raina, J.-B.; Tapiolas, D.; Willis, B.L.; Bourne, D.G. Coral-associated bacteria and their role in the biogeochemical cycling of sulfur. Appl. Environ. Microbiol. 2009, 75, 3492–3501. [Google Scholar] [CrossRef] [PubMed]
- Tandon, K.; Lu, C.-Y.; Chiang, P.-W.; Wada, N.; Yang, S.-H.; Chan, Y.-F.; Chen, P.-Y.; Chang, H.-Y.; Chiou, Y.-J.; Chou, M.-S. Comparative genomics: Dominant coral-bacterium Endozoicomonas acroporae metabolizes dimethylsulfoniopropionate (DMSP). ISME J. 2020, 14, 1290–1303. [Google Scholar] [CrossRef]
- Bourne, D.G.; Morrow, K.M.; Webster, N.S. Insights into the coral microbiome: Underpinning the health and resilience of reef ecosystems. Annu. Rev. Microbiol. 2016, 70, 317–340. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Agreda, A.; Gates, R.D.; Ainsworth, T.D. Defining the core microbiome in corals’ microbial soup. Trends Microbiol. 2017, 25, 125–140. [Google Scholar] [CrossRef]
- Zilber-Rosenberg, I.; Rosenberg, E. Role of microorganisms in the evolution of animals and plants: The hologenome theory of evolution. FEMS Microbiol. Rev. 2008, 32, 723–735. [Google Scholar] [CrossRef]
- Ainsworth, T.D.; Thurber, R.V.; Gates, R.D. The future of coral reefs: A microbial perspective. Trends Ecol. Evol. 2010, 25, 233–240. [Google Scholar] [CrossRef]
- Thurber, R.V.; Willner-Hall, D.; Rodriguez-Mueller, B.; Desnues, C.; Edwards, R.A.; Angly, F.; Dinsdale, E.; Kelly, L.; Rohwer, F. Metagenomic analysis of stressed coral holobionts. Environ. Microbiol. 2009, 11, 2148–2163. [Google Scholar] [CrossRef]
- Ainsworth, T.D.; Heron, S.F.; Ortiz, J.C.; Mumby, P.J.; Grech, A.; Ogawa, D.; Eakin, C.M.; Leggat, W. Climate change disables coral bleaching protection on the Great Barrier Reef. Science 2016, 352, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Eakin, C.; Liu, G.; Gomez, A.; De La Cour, J.; Heron, S.; Skirving, W.; Geiger, E.; Marsh, B.; Tirak, K.; Strong, A. Unprecedented three years of global coral bleaching 2014-17. In State of the Climate in 2017. Bull. Am. Meteorol. Soc. 2018, 99, S74–S75. [Google Scholar]
- Hoegh-Guldberg, O. Climate change, coral bleaching and the future of the world’s coral reefs. Mar. Freshw. Res. 1999, 50, 839–866. [Google Scholar] [CrossRef]
- Gates, R.D.; Baghdasarian, G.; Muscatine, L. Temperature stress causes host cell detachment in symbiotic cnidarians: Implications for coral bleaching. Biol. Bull. 1992, 182, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Lesser, M.P. Coral bleaching: Causes and mechanisms. In Coral Reefs: An Ecosystem in Transition; Springer: Berlin, Germany, 2011; pp. 405–419. [Google Scholar]
- Bieri, T.; Onishi, M.; Xiang, T.; Grossman, A.R.; Pringle, J.R. Relative contributions of various cellular mechanisms to loss of algae during cnidarian bleaching. PLoS ONE 2016, 11, e0152693. [Google Scholar] [CrossRef]
- Nielsen, D.A.; Petrou, K.; Gates, R.D. Coral bleaching from a single cell perspective. ISME J. 2018, 12, 1558–1567. [Google Scholar] [CrossRef]
- Glynn, P.W. Extensive ‘bleaching’and death of reef corals on the Pacific coast of Panama. Environ. Conserv. 1983, 10, 149–154. [Google Scholar] [CrossRef]
- Heron, S.F.; Maynard, J.A.; Van Hooidonk, R.; Eakin, C.M. Warming trends and bleaching stress of the world’s coral reefs 1985–2012. Sci. Rep. 2016, 6, 38402. [Google Scholar] [CrossRef]
- Hughes, T.P.; Anderson, K.D.; Connolly, S.R.; Heron, S.F.; Kerry, J.T.; Lough, J.M.; Baird, A.H.; Baum, J.K.; Berumen, M.L.; Bridge, T.C. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 2018, 359, 80–83. [Google Scholar] [CrossRef]
- Oliver, J.K.; Berkelmans, R.; Eakin, C.M. Coral bleaching in space and time. In Coral Bleaching; Springer: Berlin, Germany, 2018; pp. 27–49. [Google Scholar]
- Drury, C. Resilience in reef-building corals: The ecological and evolutionary importance of the host response to thermal stress. Mol. Ecol. 2020, 29, 448–465. [Google Scholar] [CrossRef]
- Elias, S. Loss of coral reefs. In Encyclopedia of the Anthropocene; Elsevier: Amsterdam, The Netherlands, 2018; pp. 245–258. [Google Scholar]
- Hoegh-Guldberg, O.; Kennedy, E.V.; Beyer, H.L.; McClennen, C.; Possingham, H.P. Securing a long-term future for coral reefs. Trends Ecol. Evol. 2018, 33, 936–944. [Google Scholar] [CrossRef]
- Hédouin, L.; Rouzé, H.; Berthe, C.; Perez-Rosales, G.; Martinez, E.; Chancerelle, Y.; Galand, P.E.; Lerouvreur, F.; Nugues, M.M.; Pochon, X.; et al. Contrasting patterns of mortality in polynesian coral reefs following the third global coral bleaching event in 2016. Coral Reefs 2020, 39, 939–952. [Google Scholar] [CrossRef]
- Morrow, K.; Muller, E.; Lesser, M. How does the coral microbiome cause, respond to, or modulate the bleaching process? In Coral Bleaching; Springer: Berlin, Germany, 2018; pp. 153–188. [Google Scholar]
- McFall-Ngai, M.; Hadfield, M.G.; Bosch, T.C.; Carey, H.V.; Domazet-Lošo, T.; Douglas, A.E.; Dubilier, N.; Eberl, G.; Fukami, T.; Gilbert, S.F. Animals in a bacterial world, a new imperative for the life sciences. Proc. Natl. Acad. Sci. USA 2013, 110, 3229–3236. [Google Scholar] [CrossRef]
- Krediet, C.J.; Ritchie, K.B.; Paul, V.J.; Teplitski, M. Coral-associated micro-organisms and their roles in promoting coral health and thwarting diseases. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122328. [Google Scholar] [CrossRef]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.M.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef]
- Holmes, E.; Li, J.V.; Marchesi, J.R.; Nicholson, J.K. Gut microbiota composition and activity in relation to host metabolic phenotype and disease risk. Cell Metab. 2012, 16, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M. The gut microbiota and host health: A new clinical frontier. Gut 2015, 65, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Gardner, S.G.; Camp, E.F.; Smith, D.J.; Kahlke, T.; Osman, E.O.; Gendron, G.; Hume, B.C.; Pogoreutz, C.; Voolstra, C.R.; Suggett, D.J. Coral microbiome diversity reflects mass coral bleaching susceptibility during the 2016 El Niño heat wave. Ecol. Evol. 2019, 9, 938–956. [Google Scholar] [CrossRef] [PubMed]
- Rosado, P.M.; Leite, D.C.; Duarte, G.A.; Chaloub, R.M.; Jospin, G.; da Rocha, U.N.; Saraiva, J.P.; Dini-Andreote, F.; Eisen, J.A.; Bourne, D.G. Marine probiotics: Increasing coral resistance to bleaching through microbiome manipulation. ISME J. 2019, 13, 921. [Google Scholar] [CrossRef]
- Bourne, D.G.; Garren, M.; Work, T.M.; Rosenberg, E.; Smith, G.W.; Harvell, C.D. Microbial disease and the coral holobiont. Trends Microbiol. 2009, 17, 554–562. [Google Scholar] [CrossRef]
- Kitahara, M.V.; Fukami, H.; Benzoni, F.; Huang, D. The new systematics of Scleractinia: Integrating molecular and morphological evidence. In The Cnidaria, Past, Present and Future; Springer: Berlin, Germany, 2016; pp. 41–59. [Google Scholar]
- Harrison, P.L. Sexual reproduction of scleractinian corals. In Coral Reefs: An Ecosystem in Transition; Springer: Berlin, Germany, 2011; pp. 59–85. [Google Scholar]
- Harrison, P.L.; Booth, D.J. Coral reefs: Naturally dynamic and increasingly disturbed ecosystems. In Marine Ecology; Oxford University Press: Melbourne, Australia, 2007; pp. 316–377. [Google Scholar]
- Blackall, L.L.; Wilson, B.; van Oppen, M.J. Coral—The world’s most diverse symbiotic ecosystem. Mol. Ecol. 2015, 24, 5330–5347. [Google Scholar] [CrossRef] [PubMed]
- Jokiel, P.L.; Jury, C.P.; Kuffner, I.B. Coral calcification and ocean acidification. In Coral Reefs at the Crossroads; Springer: Berlin, Germany, 2016; pp. 7–45. [Google Scholar]
- Comeau, S.; Tambutté, E.; Carpenter, R.; Edmunds, P.; Evensen, N.; Allemand, D.; Ferrier-Pagès, C.; Tambutté, S.; Venn, A. Coral calcifying fluid pH is modulated by seawater carbonate chemistry not solely seawater pH. Proc. R. Soc. B Biol. Sci. 2017, 284, 20161669. [Google Scholar] [CrossRef]
- Le Tissier, M.D.A. The nature of the skeleton and skeletogenic tissues in the Cnidaria. Hydrobiologia 1991, 216, 397–402. [Google Scholar] [CrossRef]
- Brown, B.; Bythell, J. Perspectives on mucus secretion in reef corals. Mar. Ecol. Prog. Ser. 2005, 296, 291–309. [Google Scholar] [CrossRef]
- Sweet, M.; Croquer, A.; Bythell, J. Bacterial assemblages differ between compartments within the coral holobiont. Coral Reefs 2011, 30, 39–52. [Google Scholar] [CrossRef]
- Apprill, A.; Weber, L.G.; Santoro, A.E. Distinguishing between microbial habitats unravels ecological complexity in coral microbiomes. Msystems 2016, 1. [Google Scholar] [CrossRef]
- Brown, B.; Le Tissier, M.; Bythell, J. Mechanisms of bleaching deduced from histological studies of reef corals sampled during a natural bleaching event. Mar. Biol. 1995, 122, 655–663. [Google Scholar] [CrossRef]
- Weis, V.M. Cellular mechanisms of cnidarian bleaching: Stress causes the collapse of symbiosis. J. Exp. Biol. 2008, 211, 3059–3066. [Google Scholar] [CrossRef]
- Pratte, Z.A.; Richardson, L.L. Impacts of temperature increase and acidification on thickness of the surface mucopolysaccharide layer of the caribbean coral Diploria spp. Coral Reefs 2014, 33, 487–496. [Google Scholar] [CrossRef]
- Lee, S.; Davy, S.K.; Tang, S.-L.; Kench, P.S. Mucus sugar content shapes the bacterial community structure in thermally stressed Acropora muricata. Front. Microbiol. 2016, 7, 371. [Google Scholar] [CrossRef]
- Cooper, E.L. Evolution of immune systems from self/not self to danger to artificial immune systems (AIS). Phys. Life Rev. 2010, 7, 55–78. [Google Scholar] [CrossRef]
- Putnam, N.H.; Srivastava, M.; Hellsten, U.; Dirks, B.; Chapman, J.; Salamov, A.; Terry, A.; Shapiro, H.; Lindquist, E.; Kapitonov, V.V. Sea anemone genome reveals ancestral eumetazoan gene repertoire and genomic organization. Science 2007, 317, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Bosch, T.C.; Miller, D.J. Negotiations between early evolving animals and symbionts. In The Holobiont Imperative; Springer: Berlin, Germany, 2016; pp. 57–65. [Google Scholar]
- Palmer, C.; Traylor-Knowles, N. Towards an integrated network of coral immune mechanisms. Proc. R. Soc. B Biol. Sci. 2012, 279, 4106–4114. [Google Scholar] [CrossRef] [PubMed]
- Bosch, T.C.; Augustin, R.; Anton-Erxleben, F.; Fraune, S.; Hemmrich, G.; Zill, H.; Rosenstiel, P.; Jacobs, G.; Schreiber, S.; Leippe, M. Uncovering the evolutionary history of innate immunity: The simple metazoan hydra uses epithelial cells for host defence. Dev. Comp. Immunol. 2009, 33, 559–569. [Google Scholar] [CrossRef]
- Hernandez-Agreda, A.; Leggat, W.; Ainsworth, T.D. A place for taxonomic profiling in the study of the coral prokaryotic microbiome. Fems Microbiol. Lett. 2019, 366, fnz063. [Google Scholar] [CrossRef]
- Barshis, D.J.; Ladner, J.T.; Oliver, T.A.; Seneca, F.O.; Traylor-Knowles, N.; Palumbi, S.R. Genomic basis for coral resilience to climate change. Proc. Natl. Acad. Sci. USA 2013, 110, 1387–1392. [Google Scholar] [CrossRef]
- Palmer, C.V.; McGinty, E.S.; Cummings, D.J.; Smith, S.M.; Bartels, E.; Mydlarz, L.D. Patterns of coral ecological immunology: Variation in the responses of caribbean corals to elevated temperature and a pathogen elicitor. J. Exp. Biol. 2011, 214, 4240–4249. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Wang, Y.B.; Chen, C.S.; Lin, C.Y.; Chen, S.H. Compartment-specific transcriptomics in a reef-building coral exposed to elevated temperatures. Mol. Ecol. 2014, 23, 5816–5830. [Google Scholar] [CrossRef]
- Mayfield, A.B.; Wang, Y.B.; Chen, C.S.; Chen, S.H.; Lin, C.Y. Dual-compartmental transcriptomic proteomic analysis of a marine endosymbiosis exposed to environmental change. Mol. Ecol. 2016, 25, 5944–5958. [Google Scholar] [CrossRef] [PubMed]
- Mayfield, A.B.; Chen, Y.-J.; Lu, C.-Y.; Chen, C.-S. The proteomic response of the reef coral Pocillopora acuta to experimentally elevated temperatures. PLoS ONE 2018, 13, e0192001. [Google Scholar] [CrossRef]
- Heyward, A.; Negri, A. Turbulence, cleavage, and the naked embryo: A case for coral clones. Science 2012, 335, 1064. [Google Scholar] [CrossRef]
- Dixon, G.B.; Davies, S.W.; Aglyamova, G.V.; Meyer, E.; Bay, L.K.; Matz, M.V. Genomic determinants of coral heat tolerance across latitudes. Science 2015, 348, 1460–1462. [Google Scholar] [CrossRef]
- Matz, M.V.; Treml, E.A.; Aglyamova, G.V.; Bay, L.K. Potential and limits for rapid genetic adaptation to warming in a Great Barrier Reef coral. PLoS Genet. 2018, 14, e1007220. [Google Scholar] [CrossRef]
- Hughes, T.P.; Ayre, D.; Connell, J.H. The evolutionary ecology of corals. Trends Ecol. Evol. 1992, 7, 292–295. [Google Scholar] [CrossRef]
- Baird, A.; Maynard, J.A. Coral adaptation in the face of climate change. Science 2008, 320, 315–316. [Google Scholar] [CrossRef]
- Peng, S.E.; Chen, W.N.U.; Chen, H.K.; Lu, C.Y.; Mayfield, A.B.; Fang, L.S.; Chen, C.S. Lipid bodies in coral–dinoflagellate endosymbiosis: Proteomic and ultrastructural studies. Proteomics 2011, 11, 3540–3555. [Google Scholar] [CrossRef]
- Mayfield, A.; Fan, T.-Y.; Chen, C.-S. Physiological acclimation to elevated temperature in a reef-building coral from an upwelling environment. Coral Reefs 2013, 32, 909–921. [Google Scholar] [CrossRef]
- Wada, N.; Ishimochi, M.; Matsui, T.; Pollock, F.J.; Tang, S.-L.; Ainsworth, T.D.; Willis, B.L.; Mano, N.; Bourne, D.G. Characterization of coral-associated microbial aggregates (CAMAs) within tissues of the coral Acropora hyacinthus. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Baird, A.H.; Guest, J.R.; Willis, B.L. Systematic and biogeographical patterns in the reproductive biology of scleractinian corals. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 551–571. [Google Scholar] [CrossRef]
- Padilla-Gamiño, J.L.; Pochon, X.; Bird, C.; Concepcion, G.T.; Gates, R.D. From parent to gamete: Vertical transmission of Symbiodinium (Dinophyceae) ITS2 sequence assemblages in the reef building coral Montipora capitata. PLoS ONE 2012, 7, e38440. [Google Scholar] [CrossRef] [PubMed]
- Sharp, K.H.; Distel, D.; Paul, V.J. Diversity and dynamics of bacterial communities in early life stages of the caribbean coral Porites astreoides. ISME J. 2012, 6, 790–801. [Google Scholar] [CrossRef] [PubMed]
- Leite, D.C.; Leão, P.; Garrido, A.G.; Lins, U.; Santos, H.F.; Pires, D.O.; Castro, C.B.; van Elsas, J.D.; Zilberberg, C.; Rosado, A.S. Broadcast spawning coral Mussismilia hispida can vertically transfer its associated bacterial core. Front. Microbiol. 2017, 8, 176. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Cai, L.; Yuan, T.; Tian, R.; Tong, H.; Zhang, W.; Jiang, L.; Guo, M.; Liu, S.; Qian, P.Y. Microbiome dynamics in early life stages of the scleractinian coral Acropora gemmifera in response to elevated PCO2. Environ. Microbiol. 2017, 19, 3342–3352. [Google Scholar] [CrossRef]
- Epstein, H.E.; Smith, H.A.; Torda, G.; van Oppen, M.J. Microbiome engineering: Enhancing climate resilience in corals. Front. Ecol. Environ. 2019, 17, 100–108. [Google Scholar] [CrossRef]
- Torda, G.; Donelson, J.M.; Aranda, M.; Barshis, D.J.; Bay, L.; Berumen, M.L.; Bourne, D.G.; Cantin, N.; Foret, S.; Matz, M. Rapid adaptive responses to climate change in corals. Nat. Clim. Chang. 2017, 7, 627–636. [Google Scholar] [CrossRef]
- Wakefield, T.S.; Kempf, S.C. Development of host-and symbiont-specific monoclonal antibodies and confirmation of the origin of the symbiosome membrane in a cnidarian–dinoflagellate symbiosis. Biol. Bull. 2001, 200, 127–143. [Google Scholar] [CrossRef] [PubMed]
- Stat, M.; Carter, D.; Hoegh-Guldberg, O. The evolutionary history of Symbiodinium and scleractinian hosts—symbiosis, diversity, and the effect of climate change. Perspect. Plant Ecol. Evol. Syst. 2006, 8, 23–43. [Google Scholar] [CrossRef]
- Burriesci, M.S.; Raab, T.K.; Pringle, J.R. Evidence that glucose is the major transferred metabolite in dinoflagellate–cnidarian symbiosis. J. Exp. Biol. 2012, 215, 3467–3477. [Google Scholar] [CrossRef]
- Banin, E.; Vassilakos, D.; Orr, E.; Martinez, R.J.; Rosenberg, E. Superoxide dismutase is a virulence factor produced by the coral bleaching pathogen Vibrio shiloi. Curr. Microbiol. 2003, 46, 0418–0422. [Google Scholar] [CrossRef]
- Rinkevich, B.; Loya, Y. Does light enhance calcification in hermatypic corals? Mar. Biol. 1984, 80, 1–6. [Google Scholar] [CrossRef]
- Al-Horani, F.; Al-Moghrabi, S.; De Beer, D. The mechanism of calcification and its relation to photosynthesis and respiration in the scleractinian coral Galaxea fascicularis. Mar. Biol. 2003, 142, 419–426. [Google Scholar] [CrossRef]
- Gutner-Hoch, E.; Schneider, K.; Stolarski, J.; Domart-Coulon, I.; Yam, R.; Meibom, A.; Shemesh, A.; Levy, O. Evidence for rhythmicity pacemaker in the calcification process of scleractinian coral. Sci. Rep. 2016, 6, 20191. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Nakamura, T.; Tanaka, Y.; Suzuki, A.; Yokoyama, Y.; Kawahata, H.; Sakai, K.; Gussone, N. A simple role of coral-algal symbiosis in coral calcification based on multiple geochemical tracers. Geochim. Et Cosmochim. Acta 2018, 235, 76–88. [Google Scholar] [CrossRef]
- Pochon, X.; Lajeunesse, T.C.; Pawlowski, J. Biogeographic partitioning and host specialization among foraminiferan dinoflagellate symbionts (Symbiodinium; Dinophyta). Mar. Biol. 2004, 146, 17–27. [Google Scholar] [CrossRef]
- Pochon, X.; Gates, R.D. A new Symbiodinium clade (Dinophyceae) from Soritid foraminifera in Hawai’i. Mol. Phylogenetics Evol. 2010, 56, 492–497. [Google Scholar] [CrossRef]
- Ziegler, M.; Arif, C.; Voolstra, C.R. Symbiodiniaceae diversity in Red Sea coral reefs & coral bleaching. In Coral Reefs of the Red Sea; Springer: Berlin, Germany, 2019; pp. 69–89. [Google Scholar]
- Coffroth, M.A. Genetic diversity of symbiotic dinoflagellates in the genus Symbiodinium. Protist 2005, 156, 19–34. [Google Scholar] [CrossRef]
- Rowan, R.; Knowlton, N. Intraspecific diversity and ecological zonation in coral-algal symbiosis. Proc. Natl. Acad. Sci. USA 1995, 92, 2850–2853. [Google Scholar] [CrossRef]
- Stat, M.; Pochon, X.; Cowie, R.O.; Gates, R.D. Specificity in communities of Symbiodinium in corals from Johnston Atoll. Mar. Ecol. Prog. Ser. 2009, 386, 83–96. [Google Scholar] [CrossRef]
- Bongaerts, P.; Frade, P.R.; Ogier, J.J.; Hay, K.B.; Van Bleijswijk, J.; Englebert, N.; Vermeij, M.J.; Bak, R.P.; Visser, P.M.; Hoegh-Guldberg, O. Sharing the slope: Depth partitioning of agariciid corals and associated Symbiodinium across shallow and mesophotic habitats (2–60 m) on a caribbean reef. BMC Evol. Biol. 2013, 13, 205. [Google Scholar] [CrossRef]
- Frade, P.; De Jongh, F.; Vermeulen, F.; Van Bleijswijk, J.; Bak, R. Variation in symbiont distribution between closely related coral species over large depth ranges. Mol. Ecol. 2008, 17, 691–703. [Google Scholar] [CrossRef]
- Ziegler, M.; Roder, C.; Büchel, C.; Voolstra, C.R. Niche acclimatization in Red Sea corals is dependent on flexibility of host-symbiont association. Mar. Ecol. Prog. Ser. 2015, 533, 149–161. [Google Scholar] [CrossRef]
- Iglesias-Prieto, R.; Beltran, V.; Lajeunesse, T.C.; Reyes-Bonilla, H.; Thome, P. Different algal symbionts explain the vertical distribution of dominant reef corals in the Eastern Pacific. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2004, 271, 1757–1763. [Google Scholar] [CrossRef] [PubMed]
- Reich, H.G.; Robertson, D.L.; Goodbody-Gringley, G. Do the shuffle: Changes in Symbiodinium consortia throughout juvenile coral development. PLoS ONE 2017, 12, e0171768. [Google Scholar] [CrossRef]
- Stat, M.; Gates, R.D. Clade D Symbiodinium in scleractinian corals: A “nugget” of hope, a selfish opportunist, an ominous sign, or all of the above? J. Mar. Biol. 2011, 2011, 1–9. [Google Scholar] [CrossRef]
- Eckert, R.J.; Reaume, A.M.; Sturm, A.B.; Studivan, M.S.; Voss, J.D. Depth influences Symbiodiniaceae associations among Montastraea cavernosa corals on the Belize Barrier Reef. Front. Microbiol. 2020, 11, 518. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Thornhill, D.J.; Cox, E.F.; Stanton, F.G.; Fitt, W.K.; Schmidt, G.W. High diversity and host specificity observed among symbiotic dinoflagellates in reef coral communities from Hawaii. Coral Reefs 2004, 23, 596–603. [Google Scholar] [CrossRef]
- Quigley, K.M.; Willis, B.L.; Bay, L.K. Heritability of the Symbiodinium community in vertically-and horizontally-transmitting broadcast spawning corals. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A. Host benefit and the evolution of specialization in symbiosis. Heredity 1998, 81, 599–603. [Google Scholar] [CrossRef]
- Grupstra, C.G.; Coma, R.; Ribes, M.; Leydet, K.P.; Parkinson, J.E.; McDonald, K.; Catlla, M.; Voolstra, C.R.; Hellberg, M.E.; Coffroth, M.A. Evidence for coral range expansion accompanied by reduced diversity of Symbiodinium genotypes. Coral Reefs 2017, 36, 981–985. [Google Scholar] [CrossRef]
- Decelle, J.; Carradec, Q.; Pochon, X.; Henry, N.; Romac, S.; Mahé, F.; Dunthorn, M.; Kourlaiev, A.; Voolstra, C.R.; Wincker, P. Worldwide occurrence and activity of the reef-building coral symbiont Symbiodinium in the open ocean. Curr. Biol. 2018, 28, 3625–3633.e3. [Google Scholar] [CrossRef]
- Sampayo, E.; Ridgway, T.; Bongaerts, P.; Hoegh-Guldberg, O. Bleaching susceptibility and mortality of corals are determined by fine-scale differences in symbiont type. Proc. Natl. Acad. Sci. USA 2008, 105, 10444–10449. [Google Scholar] [CrossRef]
- Thornhill, D.; Howells, E.; Wham, D.; Steury, T.; Santos, S. Population genetics of reef coral endosymbionts (Symbiodinium, Dinophyceae). Mol. Ecol. 2017, 26, 2640–2659. [Google Scholar] [CrossRef] [PubMed]
- Morris, L.A.; Voolstra, C.R.; Quigley, K.M.; Bourne, D.G.; Bay, L.K. Nutrient availability and metabolism affect the stability of coral–Symbiodiniaceae symbioses. Trends Microbiol. 2019, 27, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Amend, A.S.; Barshis, D.J.; Oliver, T.A. Coral-associated marine fungi form novel lineages and heterogeneous assemblages. ISME J. 2012, 6, 1291. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, M.; Seneca, F.O.; Yum, L.K.; Palumbi, S.R.; Voolstra, C.R. Bacterial community dynamics are linked to patterns of coral heat tolerance. Nat. Commun. 2017, 8, 14213. [Google Scholar] [CrossRef]
- Deschaseaux, E.S.; Jones, G.B.; Deseo, M.A.; Shepherd, K.M.; Kiene, R.; Swan, H.; Harrison, P.L.; Eyre, B.D. Effects of environmental factors on dimethylated sulfur compounds and their potential role in the antioxidant system of the coral holobiont. Limnol. Oceanogr. 2014, 59, 758–768. [Google Scholar] [CrossRef]
- Robbins, S.J.; Singleton, C.M.; Chan, C.X.; Messer, L.F.; Geers, A.U.; Ying, H.; Baker, A.; Bell, S.C.; Morrow, K.M.; Ragan, M.A. A genomic view of the reef-building coral Porites lutea and its microbial symbionts. Nat. Microbiol. 2019, 4, 2090–2100. [Google Scholar] [CrossRef]
- Roughgarden, J.; Gilbert, S.F.; Rosenberg, E.; Zilber-Rosenberg, I.; Lloyd, E.A. Holobionts as units of selection and a model of their population dynamics and evolution. Biol. Theory 2018, 13, 44–65. [Google Scholar] [CrossRef]
- Bernasconi, R.; Stat, M.; Koenders, A.; Paparini, A.; Bunce, M.; Huggett, M.J. Establishment of coral-bacteria symbioses reveal changes in the core bacterial community with host ontogeny. Front. Microbiol. 2019, 10, 1529. [Google Scholar] [CrossRef]
- Ritchie, K.B. Bacterial symbionts of corals and Symbiodinium. In Beneficial Microorganisms in Multicellular Life Forms; Springer: Berlin, Germany, 2012; pp. 139–150. [Google Scholar]
- Apprill, A.; Marlow, H.Q.; Martindale, M.Q.; Rappé, M.S. The onset of microbial associations in the coral Pocillopora meandrina. ISME J. 2009, 3, 685–699. [Google Scholar] [CrossRef]
- Apprill, A.; Marlow, H.Q.; Martindale, M.Q.; Rappé, M.S. Specificity of associations between bacteria and the coral Pocillopora meandrina during early development. Appl. Environ. Microbiol. 2012, 78, 7467–7475. [Google Scholar] [CrossRef]
- Sharp, K.H.; Ritchie, K.B.; Schupp, P.J.; Ritson-Williams, R.; Paul, V.J. Bacterial acquisition in juveniles of several broadcast spawning coral species. PLoS ONE 2010, 5, e10898. [Google Scholar] [CrossRef]
- Ceh, J.; van Keulen, M.; Bourne, D.G. Intergenerational transfer of specific bacteria in corals and possible implications for offspring fitness. Microb. Ecol. 2013, 65, 227–231. [Google Scholar] [CrossRef]
- Ricardo, G.F.; Jones, R.J.; Negri, A.P.; Stocker, R. That sinking feeling: Suspended sediments can prevent the ascent of coral egg bundles. Sci. Rep. 2016, 6, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.D.; Brown, B.E.; Putchim, L.; Sweet, M.J. Age-related shifts in bacterial diversity in a reef coral. PLoS ONE 2015, 10, e0144902. [Google Scholar] [CrossRef]
- van Oppen, M.J.; Blackall, L.L. Coral microbiome dynamics, functions and design in a changing world. Nat. Rev. Microbiol. 2019, 17, 557–567. [Google Scholar] [CrossRef]
- Work, T.M.; Aeby, G.S. Microbial aggregates within tissues infect a diversity of corals throughout the Indo-Pacific. Mar. Ecol. Prog. Ser. 2014, 500, 1–9. [Google Scholar] [CrossRef]
- Ainsworth, T.; Hoegh-Guldberg, O. Bacterial communities closely associated with coral tissues vary under experimental and natural reef conditions and thermal stress. Aquat. Biol. 2009, 4, 289–296. [Google Scholar] [CrossRef]
- Ainsworth, T.; Fine, M.; Blackall, L.; Hoegh-Guldberg, O. Fluorescence in situ hybridization and spectral imaging of coral-associated bacterial communities. Appl. Environ. Microbiol. 2006, 72, 3016–3020. [Google Scholar] [CrossRef]
- Tribollet, A. The boring microflora in modern coral reef ecosystems: A review of its roles. In Current Developments in Bioerosion; Springer: Berlin, Germany, 2008; pp. 67–94. [Google Scholar]
- Verbruggen, H.; Tribollet, A. Boring algae. Curr. Biol. 2011, 21, R876–R877. [Google Scholar] [CrossRef]
- Ricci, F.; Marcelino, V.R.; Blackall, L.L.; Kühl, M.; Medina, M.; Verbruggen, H. Beneath the surface: Community assembly and functions of the coral skeleton microbiome. Microbiome 2019, 7, 159. [Google Scholar] [CrossRef]
- Garren, M.; Azam, F. New method for counting bacteria associated with coral mucus. Appl. Environ. Microbiol. 2010, 76, 6128–6133. [Google Scholar] [CrossRef]
- Wild, C.; Niggl, W.; Naumann, M.S.; Haas, A.F. Organic matter release by Red Sea coral reef organisms—potential effects on microbial activity and in situ O2 availability. Mar. Ecol. Prog. Ser. 2010, 411, 61–71. [Google Scholar] [CrossRef]
- Nakajima, R.; Tanaka, Y.; Yoshida, T.; Fujisawa, T.; Nakayama, A.; Fuchinoue, Y.; Othman, B.H.R.; Toda, T. High inorganic phosphate concentration in coral mucus and its utilization by heterotrophic bacteria in a malaysian coral reef. Mar. Ecol. 2015, 36, 835–841. [Google Scholar] [CrossRef]
- Jatkar, A.A.; Brown, B.E.; Bythell, J.C.; Guppy, R.; Morris, N.J.; Pearson, J.P. Measuring mucus thickness in reef corals using a technique devised for vertebrate applications. Mar. Biol. 2010, 157, 261–267. [Google Scholar] [CrossRef]
- Teplitski, M.; Ritchie, K. How feasible is the biological control of coral diseases? Trends Ecol. Evol. 2009, 24, 378–385. [Google Scholar] [CrossRef]
- Ritchie, K.B. Regulation of microbial populations by coral surface mucus and mucus-associated bacteria. Mar. Ecol. Prog. Ser. 2006, 322, 1–14. [Google Scholar] [CrossRef]
- Kemp, D.W.; Rivers, A.R.; Kemp, K.M.; Lipp, E.K.; Porter, J.W.; Wares, J.P. Spatial homogeneity of bacterial communities associated with the surface mucus layer of the reef-building coral Acropora palmata. PLoS ONE 2015, 10, e0143790. [Google Scholar] [CrossRef]
- Morrow, K.M.; Moss, A.G.; Chadwick, N.E.; Liles, M.R. Bacterial associates of two caribbean coral species reveal species-specific distribution and geographic variability. Appl. Environ. Microbiol. 2012, 78, 6438–6449. [Google Scholar] [CrossRef]
- Ziegler, M.; Grupstra, C.G.; Barreto, M.M.; Eaton, M.; BaOmar, J.; Zubier, K.; Al-Sofyani, A.; Turki, A.J.; Ormond, R.; Voolstra, C.R. Coral bacterial community structure responds to environmental change in a host-specific manner. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- O’Brien, P.A.; Tan, S.; Yang, C.; Frade, P.R.; Andreakis, N.; Smith, H.A.; Miller, D.J.; Webster, N.S.; Zhang, G.; Bourne, D.G. Diverse coral reef invertebrates exhibit patterns of phylosymbiosis. ISME J. 2020, 14, 2211–2222. [Google Scholar] [CrossRef]
- Sunagawa, S.; Woodley, C.M.; Medina, M. Threatened corals provide underexplored microbial habitats. PLoS ONE 2010, 5, e9554. [Google Scholar] [CrossRef]
- Pollock, F.J.; McMinds, R.; Smith, S.; Bourne, D.G.; Willis, B.L.; Medina, M.; Thurber, R.V.; Zaneveld, J.R. Coral-associated bacteria demonstrate phylosymbiosis and cophylogeny. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Kelly, L.W.; Williams, G.J.; Barott, K.L.; Carlson, C.A.; Dinsdale, E.A.; Edwards, R.A.; Haas, A.F.; Haynes, M.; Lim, Y.W.; McDole, T. Local genomic adaptation of coral reef-associated microbiomes to gradients of natural variability and anthropogenic stressors. Proc. Natl. Acad. Sci. USA 2014, 111, 10227–10232. [Google Scholar] [CrossRef] [PubMed]
- Pantos, O.; Bongaerts, P.; Dennis, P.G.; Tyson, G.W.; Hoegh-Guldberg, O. Habitat-specific environmental conditions primarily control the microbiomes of the coral Seriatopora hystrix. ISME J. 2015, 9, 1916–1927. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Agreda, A.; Leggat, W.; Bongaerts, P.; Herrera, C.; Ainsworth, T.D. Rethinking the coral microbiome: Simplicity exists within a diverse microbial biosphere. MBio 2018, 9, e00812–e00818. [Google Scholar] [CrossRef] [PubMed]
- Huggett, M.J.; Apprill, A. Coral microbiome database: Integration of sequences reveals high diversity and relatedness of coral-associated microbes. Environ. Microbiol. Rep. 2019, 11, 372–385. [Google Scholar] [CrossRef]
- Raina, J.-B.; Dinsdale, E.A.; Willis, B.L.; Bourne, D.G. Do the organic sulfur compounds DMSP and DMS drive coral microbial associations? Trends Microbiol. 2010, 18, 101–108. [Google Scholar] [CrossRef]
- Olson, N.; Ainsworth, T.; Gates, R.; Takabayashi, M. Diazotrophic bacteria associated with hawaiian Montipora corals: Diversity and abundance in correlation with symbiotic dinoflagellates. J. Exp. Mar. Biol. Ecol. 2009, 371, 140–146. [Google Scholar] [CrossRef]
- Bernasconi, R.; Stat, M.; Koenders, A.; Huggett, M.J. Global networks of Symbiodinium-bacteria within the coral holobiont. Microb. Ecol. 2019, 77, 794–807. [Google Scholar] [CrossRef]
- Lesser, M.P.; Falcón, L.I.; Rodríguez-Román, A.; Enríquez, S.; Hoegh-Guldberg, O.; Iglesias-Prieto, R. Nitrogen fixation by symbiotic cyanobacteria provides a source of nitrogen for the scleractinian coral Montastraea cavernosa. Mar. Ecol. Prog. Ser. 2007, 346, 143–152. [Google Scholar] [CrossRef]
- Shnit-Orland, M.; Kushmaro, A. Coral mucus-associated bacteria: A possible first line of defense. Fems Microbiol. Ecol. 2009, 67, 371–380. [Google Scholar] [CrossRef]
- Moree, W.J.; Yang, J.Y.; Zhao, X.; Liu, W.-T.; Aparicio, M.; Atencio, L.; Ballesteros, J.; Sánchez, J.; Gavilán, R.G.; Gutiérrez, M. Imaging mass spectrometry of a coral microbe interaction with fungi. J. Chem. Ecol. 2013, 39, 1045–1054. [Google Scholar] [CrossRef]
- Grover, R.; Maguer, J.-F.; Allemand, D.; Ferrier-Pages, C. Nitrate uptake in the scleractinian coral Stylophora pistillata. Limnol. Oceanogr. 2003, 48, 2266–2274. [Google Scholar] [CrossRef]
- Ezzat, L.; Maguer, J.-F.; Grover, R.; Ferrier-Pagès, C. New insights into carbon acquisition and exchanges within the coral–dinoflagellate symbiosis under NH4+ and NO3− supply. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150610. [Google Scholar] [CrossRef]
- Matthews, J.L.; Raina, J.B.; Kahlke, T.; Seymour, J.R.; van Oppen, M.J.; Suggett, D.J. Symbiodiniaceae-bacteria interactions: Rethinking metabolite exchange in reef-building corals as multi-partner metabolic networks. Environ. Microbiol. 2020, 22, 1675–1687. [Google Scholar] [CrossRef]
- Camp, E.F.; Kahlke, T.; Nitschke, M.R.; Varkey, D.; Fisher, N.L.; Fujise, L.; Goyen, S.; Hughes, D.J.; Lawson, C.A.; Ros, M. Revealing changes in the microbiome of Symbiodiniaceae under thermal stress. Environ. Microbiol. 2020, 22, 1294–1309. [Google Scholar] [CrossRef]
- Benavides, M.; Bednarz, V.N.; Ferrier-Pagès, C. Diazotrophs: Overlooked key players within the coral symbiosis and tropical reef ecosystems? Front. Mar. Sci. 2017, 4, 10. [Google Scholar] [CrossRef]
- Skindersoe, M.E.; Ettinger-Epstein, P.; Rasmussen, T.B.; Bjarnsholt, T.; de Nys, R.; Givskov, M. Quorum sensing antagonism from marine organisms. Mar. Biotechnol. 2008, 10, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Golberg, K.; Eltzov, E.; Shnit-Orland, M.; Marks, R.S.; Kushmaro, A. Characterization of quorum sensing signals in coral-associated bacteria. Microb. Ecol. 2011, 61, 783–792. [Google Scholar] [CrossRef]
- Bythell, J.C.; Wild, C. Biology and ecology of coral mucus release. J. Exp. Mar. Biol. Ecol. 2011, 408, 88–93. [Google Scholar] [CrossRef]
- Krediet, C.J.; Ritchie, K.B.; Alagely, A.; Teplitski, M. Members of native coral microbiota inhibit glycosidases and thwart colonization of coral mucus by an opportunistic pathogen. ISME J. 2013, 7, 980. [Google Scholar] [CrossRef] [PubMed]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal–response systems in gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576. [Google Scholar] [CrossRef]
- Miller, M.B.; Bassler, B.L. Quorum sensing in bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef]
- Kalia, V.C.; Patel, S.K.; Kang, Y.C.; Lee, J.-K. Quorum sensing inhibitors as antipathogens: Biotechnological applications. Biotechnol. Adv. 2018, 37, 68–90. [Google Scholar] [CrossRef]
- Certner, R.H.; Vollmer, S.V. Inhibiting bacterial quorum sensing arrests coral disease development and disease-associated microbes. Environ. Microbiol. 2018, 20, 645–657. [Google Scholar] [CrossRef]
- Littman, R.A.; Bourne, D.G.; Willis, B.L. Responses of coral-associated bacterial communities to heat stress differ with Symbiodinium type on the same coral host. Mol. Ecol. 2010, 19, 1978–1990. [Google Scholar] [CrossRef]
- McDevitt-Irwin, J.M.; Baum, J.K.; Garren, M.; Vega Thurber, R.L. Responses of coral-associated bacterial communities to local and global stressors. Front. Mar. Sci. 2017, 4, 262. [Google Scholar] [CrossRef]
- Bourne, D.; Iida, Y.; Uthicke, S.; Smith-Keune, C. Changes in coral-associated microbial communities during a bleaching event. ISME J. 2008, 2, 350. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhao, S.; Tang, J.; Liu, Z.; Wu, Y.; Wang, Y.; Lin, S. Altered immune landscape and disrupted coral-Symbiodinium symbiosis in the scleractinian coral Pocillopora damicornis by Vibrio coralliilyticus challenge. Front. Physiol. 2019, 10, 366. [Google Scholar] [CrossRef]
- Ainsworth, T.; Fine, M.; Roff, G.; Hoegh-Guldberg, O. Bacteria are not the primary cause of bleaching in the mediterranean coral Oculina patagonica. ISME J. 2008, 2, 67. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lin, Z.J.; Cai, Z.H.; Zeng, Y.H.; Zhu, J.M.; Du, X.P. Opportunistic bacteria use quorum sensing to disturb coral symbiotic communities and mediate the occurrence of coral bleaching. Environ. Microbiol. 2020, 22, 1944–1962. [Google Scholar] [CrossRef] [PubMed]
- Thurber, R.V.; Payet, J.P.; Thurber, A.R.; Correa, A.M. Virus–host interactions and their roles in coral reef health and disease. Nat. Rev. Microbiol. 2017, 15, 205. [Google Scholar] [CrossRef]
- Weynberg, K.D. Viruses in marine ecosystems: From open waters to coral reefs. In Advances in Virus Research; Elsevier: Amsterdam, The Netherlands, 2018; Volume 101, pp. 1–38. [Google Scholar]
- Leruste, A.; Bouvier, T.; Bettarel, Y. Enumerating viruses in coral mucus. Appl. Environ. Microbiol. 2012, 78, 6377–6379. [Google Scholar] [CrossRef] [PubMed]
- Nguyen-Kim, H.; Bouvier, T.; Bouvier, C.; Doan-Nhu, H.; Nguyen-Ngoc, L.; Rochelle-Newall, E.; Baudoux, A.C.; Desnues, C.; Reynaud, S.; Ferrier-Pages, C. High occurrence of viruses in the mucus layer of scleractinian corals. Environ. Microbiol. Rep. 2014, 6, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Sweet, M.; Bythell, J. The role of viruses in coral health and disease. J. Invertebr. Pathol. 2017, 147, 136–144. [Google Scholar] [CrossRef]
- Wood-Charlson, E.M.; Weynberg, K.D.; Suttle, C.A.; Roux, S.; Van Oppen, M.J. Metagenomic characterization of viral communities in corals: Mining biological signal from methodological noise. Environ. Microbiol. 2015, 17, 3440–3449. [Google Scholar] [CrossRef]
- Correa, A.; Ainsworth, T.D.; Rosales, S.M.; Thurber, A.R.; Butler, C.R.; Vega Thurber, R.L. Viral outbreak in corals associated with an in situ bleaching event: Atypical herpes-like viruses and a new megavirus infecting Symbiodinium. Front. Microbiol. 2016, 7, 127. [Google Scholar] [CrossRef]
- Barr, J.J.; Auro, R.; Furlan, M.; Whiteson, K.L.; Erb, M.L.; Pogliano, J.; Stotland, A.; Wolkowicz, R.; Cutting, A.S.; Doran, K.S. Bacteriophage adhering to mucus provide a non–host-derived immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 10771–10776. [Google Scholar] [CrossRef]
- Middelboe, M.; Lyck, P.G. Regeneration of dissolved organic matter by viral lysis in marine microbial communities. Aquat. Microb. Ecol. 2002, 27, 187–194. [Google Scholar] [CrossRef]
- Weitz, J.S.; Wilhelm, S.W. Ocean viruses and their effects on microbial communities and biogeochemical cycles. F1000 Biol. Rep. 2012, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Payet, J.P.; McMinds, R.; Burkepile, D.E.; Vega Thurber, R.L. Unprecedented evidence for high viral abundance and lytic activity in coral reef waters of the South Pacific Ocean. Front. Microbiol. 2014, 5, 493. [Google Scholar] [CrossRef] [PubMed]
- Weynberg, K.D.; Voolstra, C.R.; Neave, M.J.; Buerger, P.; Van Oppen, M.J. From cholera to corals: Viruses as drivers of virulence in a major coral bacterial pathogen. Sci. Rep. 2015, 5, 17889. [Google Scholar] [CrossRef]
- Kwong, W.K.; del Campo, J.; Mathur, V.; Vermeij, M.J.; Keeling, P.J. A widespread coral-infecting apicomplexan with chlorophyll biosynthesis genes. Nature 2019, 568, 103. [Google Scholar] [CrossRef] [PubMed]
- Clerissi, C.; Brunet, S.; Vidal-Dupiol, J.; Adjeroud, M.; Lepage, P.; Guillou, L.; Escoubas, J.-M.; Toulza, E. Protists within corals: The hidden diversity. Front. Microbiol. 2018, 9, 2043. [Google Scholar] [CrossRef] [PubMed]
- Kornmann, P.; Sahling, P.-H. Ostreobium quekettii (Codiales, Chlorophyta). Helgoländer Meeresunters 1980, 34, 115. [Google Scholar] [CrossRef]
- del Campo, J.; Pombert, J.-F.; Šlapeta, J.; Larkum, A.; Keeling, P.J. The ‘other’coral symbiont: Ostreobium diversity and distribution. ISME J. 2017, 11, 296. [Google Scholar] [CrossRef]
- Aline, T. Dissolution of dead corals by euendolithic microorganisms across the Northern Great Barrier Reef (Australia). Microb. Ecol. 2008, 55, 569–580. [Google Scholar] [CrossRef]
- Reyes-Nivia, C.; Diaz-Pulido, G.; Kline, D.; Guldberg, O.H.; Dove, S. Ocean acidification and warming scenarios increase microbioerosion of coral skeletons. Glob. Chang. Biol. 2013, 19, 1919–1929. [Google Scholar] [CrossRef]
- Grange, J.; Rybarczyk, H.; Tribollet, A. The three steps of the carbonate biogenic dissolution process by microborers in coral reefs (New Caledonia). Environ. Sci. Pollut. Res. 2015, 22, 13625–13637. [Google Scholar] [CrossRef]
- Leggat, W.P.; Camp, E.F.; Suggett, D.J.; Heron, S.F.; Fordyce, A.J.; Gardner, S.; Deakin, L.; Turner, M.; Beeching, L.J.; Kuzhiumparambil, U. Rapid coral decay is associated with marine heatwave mortality events on reefs. Curr. Biol. 2019, 29, 2723–2730. [Google Scholar] [CrossRef] [PubMed]
- Fine, M.; Loya, Y. Endolithic algae: An alternative source of photoassimilates during coral bleaching. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2002, 269, 1205–1210. [Google Scholar] [CrossRef]
- Golubic, S.; Radtke, G.; Le Campion-Alsumard, T. Endolithic fungi in marine ecosystems. Trends Microbiol. 2005, 13, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Wegley, L.; Edwards, R.; Rodriguez-Brito, B.; Liu, H.; Rohwer, F. Metagenomic analysis of the microbial community associated with the coral Porites astreoides. Environ. Microbiol. 2007, 9, 2707–2719. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, T.; Fordyce, A.; Camp, E. The other microeukaryotes of the coral reef microbiome. Trends Microbiol. 2017, 25, 980–991. [Google Scholar] [CrossRef]
- Nyström, M.; Folke, C.; Moberg, F. Coral reef disturbance and resilience in a human-dominated environment. Trends Ecol. Evol. 2000, 15, 413–417. [Google Scholar] [CrossRef]
- Mcleod, E.; Anthony, K.R.; Mumby, P.J.; Maynard, J.; Beeden, R.; Graham, N.A.; Heron, S.F.; Hoegh-Guldberg, O.; Jupiter, S.; MacGowan, P. The future of resilience-based management in coral reef ecosystems. J. Environ. Manag. 2019, 233, 291–301. [Google Scholar] [CrossRef]
- Palumbi, S.R.; McLeod, K.L.; Grünbaum, D. Ecosystems in action: Lessons from marine ecology about recovery, resistance, and reversibility. BioScience 2008, 58, 33–42. [Google Scholar] [CrossRef]
- Durante, M.K.; Baums, I.B.; Williams, D.E.; Vohsen, S.; Kemp, D.W. What drives phenotypic divergence among coral clonemates of Acropora palmata? Mol. Ecol. 2019, 28, 3208–3224. [Google Scholar] [CrossRef]
- Ainsworth, T.D.; Gates, R.D. Corals’ microbial sentinels. Science 2016, 352, 1518–1519. [Google Scholar] [CrossRef]
- Glasl, B.; Webster, N.S.; Bourne, D.G. Microbial indicators as a diagnostic tool for assessing water quality and climate stress in coral reef ecosystems. Mar. Biol. 2017, 164, 91. [Google Scholar] [CrossRef]
- Sommer, F.; Anderson, J.M.; Bharti, R.; Raes, J.; Rosenstiel, P. The resilience of the intestinal microbiota influences health and disease. Nat. Rev. Microbiol. 2017, 15, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Pratte, Z.A.; Richardson, L.L. Microbiome dynamics of two differentially resilient corals. Dis. Aquat. Org. 2018, 131, 213–226. [Google Scholar] [CrossRef]
- La Riviere, M.; Roumagnac, M.; Garrabou, J.; Bally, M. Transient shifts in bacterial communities associated with the temperate gorgonian Paramuricea clavata in the Northwestern Mediterranean Sea. PLoS ONE 2013, 8, e57385. [Google Scholar] [CrossRef] [PubMed]
- Voolstra, C.R.; Ziegler, M. Adapting with microbial help: Microbiome flexibility facilitates rapid responses to environmental change. BioEssays 2020, 42, 2000004. [Google Scholar] [CrossRef]
- Baker, A.C. Flexibility and specificity in coral-algal symbiosis: Diversity, ecology, and biogeography of Symbiodinium. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 661–689. [Google Scholar] [CrossRef]
- Hume, B.C.; Voolstra, C.R.; Arif, C.; D’Angelo, C.; Burt, J.A.; Eyal, G.; Loya, Y.; Wiedenmann, J. Ancestral genetic diversity associated with the rapid spread of stress-tolerant coral symbionts in response to holocene climate change. Proc. Natl. Acad. Sci. USA 2016, 113, 4416–4421. [Google Scholar] [CrossRef]
- Levin, R.A.; Beltran, V.H.; Hill, R.; Kjelleberg, S.; McDougald, D.; Steinberg, P.D.; Van Oppen, M.J. Sex, scavengers, and chaperones: Transcriptome secrets of divergent Symbiodinium thermal tolerances. Mol. Biol. Evol. 2016, 33, 2201–2215. [Google Scholar] [CrossRef]
- Ladner, J.T.; Barshis, D.J.; Palumbi, S.R. Protein evolution in two co-occurring types of Symbiodinium: An exploration into the genetic basis of thermal tolerance in Symbiodinium clade D. BMC Evol. Biol. 2012, 12, 217. [Google Scholar] [CrossRef]
- Alberdi, A.; Aizpurua, O.; Bohmann, K.; Zepeda-Mendoza, M.L.; Gilbert, M.T.P. Do vertebrate gut metagenomes confer rapid ecological adaptation? Trends Ecol. Evol. 2016, 31, 689–699. [Google Scholar] [CrossRef]
- Silverstein, R.N.; Cunning, R.; Baker, A.C. Change in algal symbiont communities after bleaching, not prior heat exposure, increases heat tolerance of reef corals. Glob. Chang. Biol. 2015, 21, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Berkelmans, R.; van Oppen, M.J.; Mieog, J.C.; Sinclair, W. A community change in the algal endosymbionts of a scleractinian coral following a natural bleaching event: Field evidence of acclimatization. Proc. R. Soc. B Biol. Sci. 2008, 275, 1359–1365. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.J.; Suggett, D.J.; Baker, N.R. Is photoinhibition of zooxanthellae photosynthesis the primary cause of thermal bleaching in corals? Glob. Chang. Biol. 2005, 11, 1–11. [Google Scholar] [CrossRef]
- Gardner, S.G.; Raina, J.-B.; Nitschke, M.R.; Nielsen, D.A.; Stat, M.; Motti, C.A.; Ralph, P.J.; Petrou, K. A multi-trait systems approach reveals a response cascade to bleaching in corals. BMC Biol. 2017, 15, 1–14. [Google Scholar] [CrossRef]
- Rosenberg, E.; Zilber-Rosenberg, I. Symbiosis and development: The hologenome concept. Birth Defects Res. Part C Embryo Today Rev. 2011, 93, 56–66. [Google Scholar] [CrossRef]
- Santos, H.F.; Carmo, F.L.; Duarte, G.; Dini-Andreote, F.; Castro, C.B.; Rosado, A.S.; Van Elsas, J.D.; Peixoto, R.S. Climate change affects key nitrogen-fixing bacterial populations on coral reefs. ISME J. 2014, 8, 2272–2279. [Google Scholar] [CrossRef] [PubMed]
- Cardini, U.; Bednarz, V.N.; Naumann, M.S.; van Hoytema, N.; Rix, L.; Foster, R.A.; Al-Rshaidat, M.M.; Wild, C. Functional significance of dinitrogen fixation in sustaining coral productivity under oligotrophic conditions. Proc. R. Soc. B Biol. Sci. 2015, 282, 20152257. [Google Scholar] [CrossRef] [PubMed]
- Ferrier-Pagès, C.; Rottier, C.; Beraud, E.; Levy, O. Experimental assessment of the feeding effort of three scleractinian coral species during a thermal stress: Effect on the rates of photosynthesis. J. Exp. Mar. Biol. Ecol. 2010, 390, 118–124. [Google Scholar] [CrossRef]
- Godinot, C.; Houlbreque, F.; Grover, R.; Ferrier-Pagès, C. Coral uptake of inorganic phosphorus and nitrogen negatively affected by simultaneous changes in temperature and pH. PLoS ONE 2011, 6, e25024. [Google Scholar] [CrossRef] [PubMed]
- Mao-Jones, J.; Ritchie, K.B.; Jones, L.E.; Ellner, S.P. How microbial community composition regulates coral disease development. PLoS Biol. 2010, 8, e1000345. [Google Scholar] [CrossRef]
- Levas, S.; Grottoli, A.G.; Schoepf, V.; Aschaffenburg, M.; Baumann, J.; Bauer, J.E.; Warner, M.E. Can heterotrophic uptake of dissolved organic carbon and zooplankton mitigate carbon budget deficits in annually bleached corals? Coral Reefs 2016, 35, 495–506. [Google Scholar] [CrossRef]
- Grottoli, A.G.; Rodrigues, L.J.; Palardy, J.E. Heterotrophic plasticity and resilience in bleached corals. Nature 2006, 440, 1186–1189. [Google Scholar] [CrossRef] [PubMed]
- Grottoli, A.G.; Warner, M.E.; Levas, S.J.; Aschaffenburg, M.D.; Schoepf, V.; McGinley, M.; Baumann, J.; Matsui, Y. The cumulative impact of annual coral bleaching can turn some coral species winners into losers. Glob. Chang. Biol. 2014, 20, 3823–3833. [Google Scholar] [CrossRef] [PubMed]
- van Oppen, M.J.; Bongaerts, P.; Frade, P.; Peplow, L.M.; Boyd, S.E.; Nim, H.T.; Bay, L.K. Adaptation to reef habitats through selection on the coral animal and its associated microbiome. Mol. Ecol. 2018, 27, 2956–2971. [Google Scholar] [CrossRef]
- Buddemeier, R.W.; Fautin, D.G. Coral bleaching as an adaptive mechanism. Bioscience 1993, 43, 320–326. [Google Scholar] [CrossRef]
- Kinzie, R.A., III; Takayama, M.; Santos, S.R.; Coffroth, M.A. The adaptive bleaching hypothesis: Experimental tests of critical assumptions. Biol. Bull. 2001, 200, 51–58. [Google Scholar] [CrossRef]
- Manzello, D.P.; Matz, M.V.; Enochs, I.C.; Valentino, L.; Carlton, R.D.; Kolodziej, G.; Serrano, X.; Towle, E.K.; Jankulak, M. Role of host genetics and heat-tolerant algal symbionts in sustaining populations of the endangered coral Orbicella faveolata in the Florida Keys with ocean warming. Glob. Chang. Biol. 2019, 25, 1016–1031. [Google Scholar] [CrossRef] [PubMed]
- Silverstein, R.N.; Cunning, R.; Baker, A.C. Tenacious D: Symbiodinium in clade D remain in reef corals at both high and low temperature extremes despite impairment. J. Exp. Biol. 2017, 220, 1192–1196. [Google Scholar] [CrossRef]
- Goulet, T.L.; Coffroth, M.A. Stability of an octocoral-algal symbiosis over time and space. Mar. Ecol. Prog. Ser. 2003, 250, 117–124. [Google Scholar] [CrossRef]
- McGinley, M.P.; Aschaffenburg, M.D.; Pettay, D.T.; Smith, R.T.; LaJeunesse, T.C.; Warner, M.E. Symbiodinium spp. in colonies of Eastern Pacific Pocillopora spp. are highly stable despite the prevalence of low-abundance background populations. Mar. Ecol. Prog. Ser. 2012, 462, 1–7. [Google Scholar] [CrossRef]
- Quigley, K.; Willis, B.; Kenkel, C. Transgenerational inheritance of shuffled symbiont communities in the coral Montipora digitata. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bay, L.K.; Doyle, J.; Logan, M.; Berkelmans, R. Recovery from bleaching is mediated by threshold densities of background thermo-tolerant symbiont types in a reef-building coral. R. Soc. Open Sci. 2016, 3, 160322. [Google Scholar] [PubMed]
- Berkelmans, R.; Van Oppen, M.J. The role of zooxanthellae in the thermal tolerance of corals: A ‘nugget of hope’for coral reefs in an era of climate change. Proc. R. Soc. B Biol. Sci. 2006, 273, 2305–2312. [Google Scholar] [CrossRef]
- Mieog, J.C.; Olsen, J.L.; Berkelmans, R.; Bleuler-Martinez, S.A.; Willis, B.L.; van Oppen, M.J. The roles and interactions of symbiont, host and environment in defining coral fitness. PLoS ONE 2009, 4, e6364. [Google Scholar] [CrossRef]
- Little, A.F.; Van Oppen, M.J.; Willis, B.L. Flexibility in algal endosymbioses shapes growth in reef corals. Science 2004, 304, 1492–1494. [Google Scholar] [PubMed]
- Jones, A.; Berkelmans, R. Potential costs of acclimatization to a warmer climate: Growth of a reef coral with heat tolerant vs. sensitive symbiont types. PLoS ONE 2010, 5, e10437. [Google Scholar] [CrossRef]
- Thornhill, D.J.; LaJeunesse, T.C.; Kemp, D.W.; Fitt, W.K.; Schmidt, G.W. Multi-year, seasonal genotypic surveys of coral-algal symbioses reveal prevalent stability or post-bleaching reversion. Mar. Biol. 2006, 148, 711–722. [Google Scholar] [CrossRef]
- Matthews, J.L.; Crowder, C.M.; Oakley, C.A.; Lutz, A.; Roessner, U.; Meyer, E.; Grossman, A.R.; Weis, V.M.; Davy, S.K. Optimal nutrient exchange and immune responses operate in partner specificity in the cnidarian-dinoflagellate symbiosis. Proc. Natl. Acad. Sci. USA 2017, 114, 13194–13199. [Google Scholar]
- Gabay, Y.; Weis, V.M.; Davy, S.K. Symbiont identity influences patterns of symbiosis establishment, host growth, and asexual reproduction in a model cnidarian-dinoflagellate symbiosis. Biol. Bull. 2018, 234, 1–10. [Google Scholar]
- Lawrence, S.A.; Floge, S.A.; Davy, J.E.; Davy, S.K.; Wilson, W.H. Exploratory analysis of Symbiodinium transcriptomes reveals potential latent infection by large dsdna viruses. Environ. Microbiol. 2017, 19, 3909–3919. [Google Scholar]
- Levin, R.A.; Voolstra, C.R.; Weynberg, K.D.; Van Oppen, M.J.H. Evidence for a role of viruses in the thermal sensitivity of coral photosymbionts. ISME J. 2017, 11, 808. [Google Scholar] [CrossRef]
- Davey, M.; Holmes, G.; Johnstone, R. High rates of nitrogen fixation (acetylene reduction) on coral skeletons following bleaching mortality. Coral Reefs 2008, 27, 227–236. [Google Scholar] [CrossRef]
- Paerl, H.W.; Prufert, L.E. Oxygen-poor microzones as potential sites of microbial N2 fixation in nitrogen-depleted aerobic marine waters. Appl. Environ. Microbiol. 1987, 53, 1078–1087. [Google Scholar] [CrossRef] [PubMed]
- Tribollet, A.; Chauvin, A.; Cuet, P. Carbonate dissolution by reef microbial borers: A biogeological process producing alkalinity under different PCO2 conditions. Facies 2019, 65, 9. [Google Scholar] [CrossRef]
- Normile, D. El niño’s warmth devastating reefs worldwide. Science 2016, 352, 15–16. [Google Scholar] [PubMed]
- Wilkinson, C. Status of Coral Reefs of the World: 2008; Global Coral Reef Monitoring Network and Reef and Rainforest Research Centre: Townsville, Australia, 2008. [Google Scholar]
- Hughes, T.P.; Kerry, J.T.; Álvarez-Noriega, M.; Álvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar]
- Peixoto, R.S.; Rosado, P.M.; Leite, D.C.D.A.; Rosado, A.S.; Bourne, D.G. Beneficial microorganisms for corals (bmc): Proposed mechanisms for coral health and resilience. Front. Microbiol. 2017, 8, 341. [Google Scholar] [CrossRef]
- ados Santos, H.F.; Duarte, G.A.S.; da Costa Rachid, C.T.; Chaloub, R.M.; Calderon, E.N.; de Barros Marangoni, L.F.; Bianchini, A.; Nudi, A.H.; Do Carmo, F.L.; Van Elsas, J.D. Impact of oil spills on coral reefs can be reduced by bioremediation using probiotic microbiota. Sci. Rep. 2015, 5, 1–11. [Google Scholar]
- Chakravarti, L.J.; Beltran, V.H.; van Oppen, M.J. Rapid thermal adaptation in photosymbionts of reef-building corals. Glob. Chang. Biol. 2017, 23, 4675–4688. [Google Scholar] [CrossRef]
- Chakravarti, L.J.; van Oppen, M.J. Experimental evolution in coral photosymbionts as a tool to increase thermal tolerance. Front. Mar. Sci. 2018, 5, 227. [Google Scholar] [CrossRef]
- Roth, F.; Saalmann, F.; Thomson, T.; Coker, D.; Villalobos, R.; Jones, B.; Wild, C.; Carvalho, S. Coral reef degradation affects the potential for reef recovery after disturbance. Mar. Environ. Res. 2018, 142, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Baird, A.H.; Bellwood, D.R.; Card, M.; Connolly, S.R.; Folke, C.; Grosberg, R.; Hoegh-Guldberg, O.; Jackson, J.B.; Kleypas, J. Climate change, human impacts, and the resilience of coral reefs. Science 2003, 301, 929–933. [Google Scholar] [CrossRef]
- Hughes, T.P.; Graham, N.A.; Jackson, J.B.; Mumby, P.J.; Steneck, R.S. Rising to the challenge of sustaining coral reef resilience. Trends Ecol. Evol. 2010, 25, 633–642. [Google Scholar] [CrossRef]
- Voolstra, C.R.; Miller, D.J.; Ragan, M.A.; Hoffmann, A.; Hoegh-Guldberg, O.; Bourne, D.; Ball, E.; Ying, H.; Foret, S.; Takahashi, S. The refuge 2020 consortium—Using “omics” approaches to explore the adaptability and resilience of coral holobionts to environmental change. Front. Mar. Sci. 2015, 2, 68. [Google Scholar]
- Bhattacharya, D.; Agrawal, S.; Aranda, M.; Baumgarten, S.; Belcaid, M.; Drake, J.L.; Erwin, D.; Foret, S.; Gates, R.D.; Gruber, D.F. Comparative genomics explains the evolutionary success of reef-forming corals. Elife 2016, 5, e13288. [Google Scholar] [CrossRef] [PubMed]
- Cleves, P.A.; Shumaker, A.; Lee, J.; Putnam, H.M.; Bhattacharya, D. Unknown to known: Advancing knowledge of coral gene function. Trends Genet. 2020, 36, 93–104. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boilard, A.; Dubé, C.E.; Gruet, C.; Mercière, A.; Hernandez-Agreda, A.; Derome, N. Defining Coral Bleaching as a Microbial Dysbiosis within the Coral Holobiont. Microorganisms 2020, 8, 1682. https://doi.org/10.3390/microorganisms8111682
Boilard A, Dubé CE, Gruet C, Mercière A, Hernandez-Agreda A, Derome N. Defining Coral Bleaching as a Microbial Dysbiosis within the Coral Holobiont. Microorganisms. 2020; 8(11):1682. https://doi.org/10.3390/microorganisms8111682
Chicago/Turabian StyleBoilard, Aurélie, Caroline E. Dubé, Cécile Gruet, Alexandre Mercière, Alejandra Hernandez-Agreda, and Nicolas Derome. 2020. "Defining Coral Bleaching as a Microbial Dysbiosis within the Coral Holobiont" Microorganisms 8, no. 11: 1682. https://doi.org/10.3390/microorganisms8111682
APA StyleBoilard, A., Dubé, C. E., Gruet, C., Mercière, A., Hernandez-Agreda, A., & Derome, N. (2020). Defining Coral Bleaching as a Microbial Dysbiosis within the Coral Holobiont. Microorganisms, 8(11), 1682. https://doi.org/10.3390/microorganisms8111682