Autothermal Thermophilic Aerobic Digestion (ATAD) for Heat, Gas, and Production of a Class A Biosolids with Fertilizer Potential



Abstract
1. Introduction
2. Limitations of Culture-Based Techniques in Determining ATAD Microbial Diversity
3. Sampling of ATAD Diversity
4. PCR-DGGE Based Molecular Methodology to Evaluate ATAD Diversity
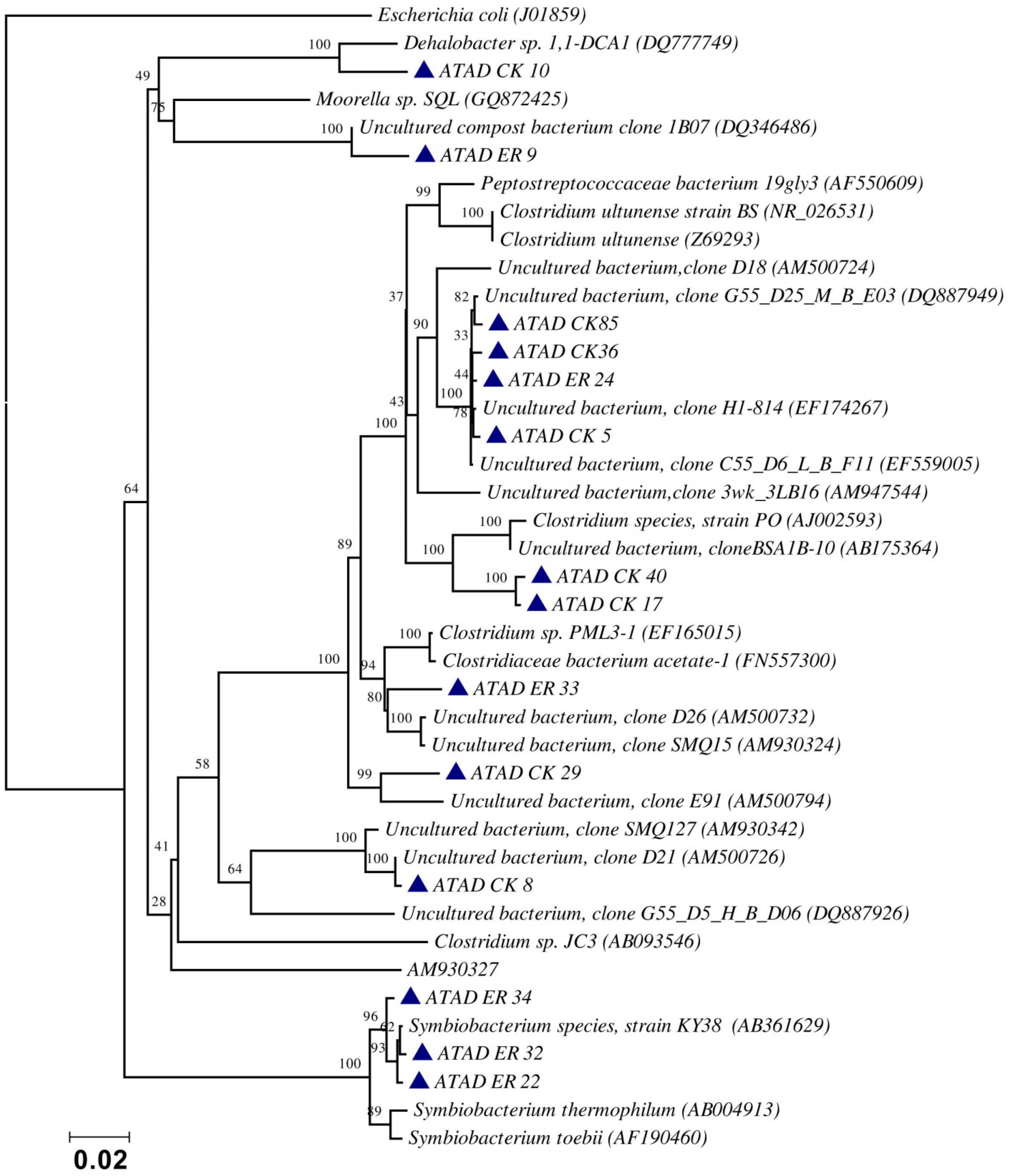
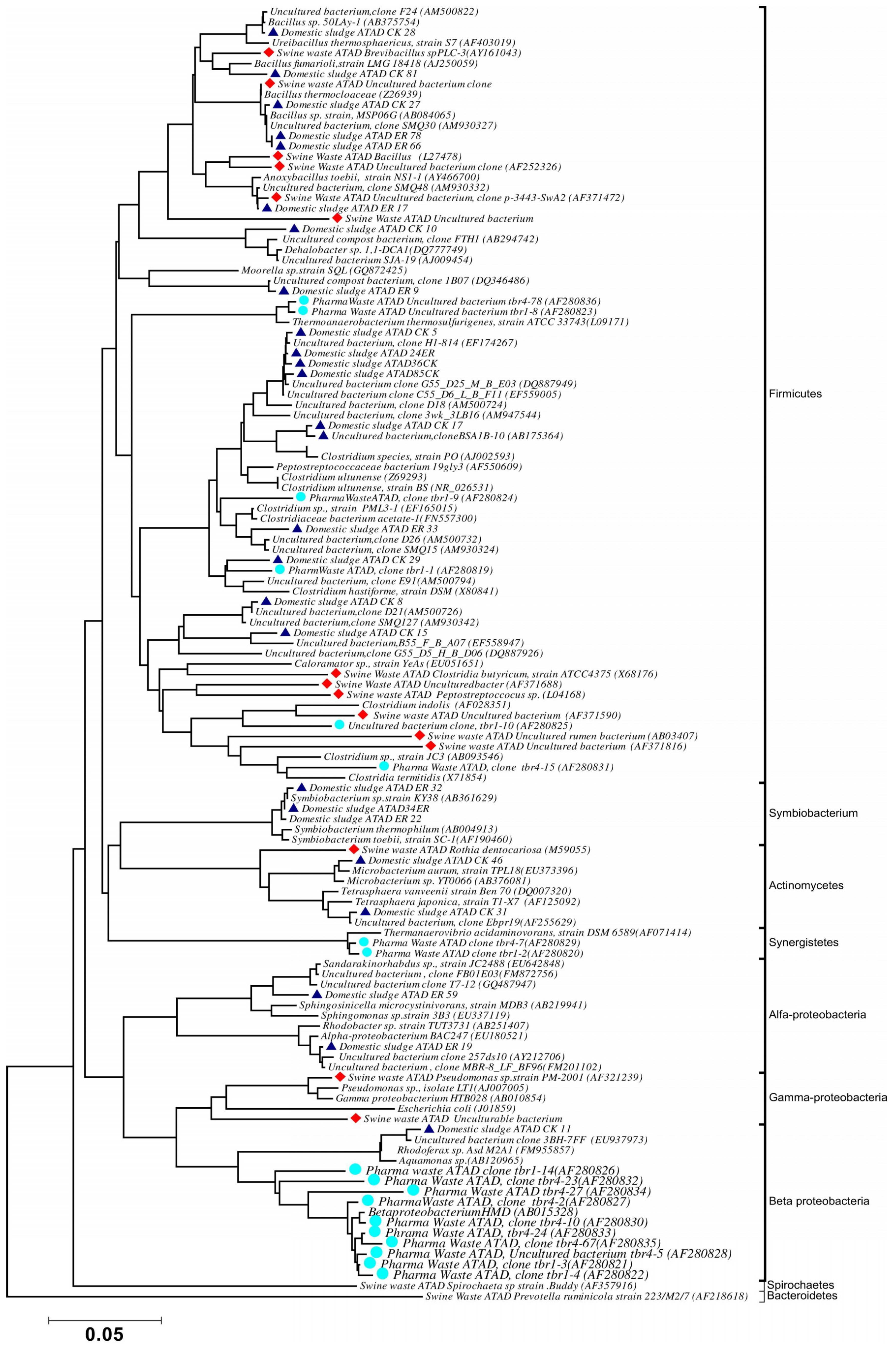
5. Phylogenetic Analysis of the ATAD Community at the Thermophilic Stage
6. Elimination of Pathogens
7. Metabolic Drivers of the Autothermal Niche
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kelly, H.G.; Melcer, H.; Mavinic, D.S. Autothermal thermophilic aerobic digestion of municipal sludges: A one-year, full-scale demonstration project. Water Environ. Res. 1993, 65, 849–861. [Google Scholar] [CrossRef]
- Piterina, A.V.; Bartlett, J.; Tony Pembroke, J. Phylogenetic analysis of the bacterial community in a full scale autothermal thermophilic aerobic digester (ATAD) treating mixed domestic wastewater sludge for land spread. Water Res. 2012, 46, 2488–2504. [Google Scholar] [CrossRef] [PubMed]
- Tashiro, Y.; Kanda, K.; Asakura, Y.; Kii, T.; Cheng, H.; Poudel, P.; Okugawa, Y.; Tashiro, K.; Sakai, K. A Unique Autothermal Thermophilic Aerobic Digestion Process Showing a Dynamic Transition of Physicochemical and Bacterial Characteristics from the Mesophilic to the Thermophilic Phase. Appl. Environ. Microbiol. 2018, 84, e02537. [Google Scholar] [CrossRef] [PubMed]
- Malladi, B.; Ingham, S.C. Thermophilic aerobic treatment of potato-processing wastewater. World J. Microbiol. Biotechnol. 1993, 9, 45–49. [Google Scholar] [CrossRef] [PubMed]
- LaPara, T.M.; Alleman, J.E. Thermophilic aerobic biological wastewater treatment. Water Res. 1999, 33, 895–908. [Google Scholar] [CrossRef]
- Liu, S.; Ning, P.; Zhu, N.; Li, L.Y.; Gong, X. Semicontinuous Operation of One-Stage Autothermal Thermophilic Aerobic Digestion of Sewage Sludge: Effects of Retention Time. J. Environ. Eng. 2012, 139, 422–427. [Google Scholar] [CrossRef]
- Piterina, A.V.; McCusland, C.; Bartlett, J.; Pembroke, J.T. Microbial Ecology of Authothermal Aerobic Digestion (ATAD): Diversity, Dynamics and Activity of Bacterial Communities Involved in Treatment of a Municipal Wastewater. In Modern Multidisciplinary Applied Microbiology; John Wiley and Sons: Hoboken, NJ, USA, 2008; pp. 526–535. ISBN 9783527611904. [Google Scholar]
- Layden, N.M.; Mavinic, D.S.; Kelly, H.G.; Moles, R.; Bartlett, J. Autothermal thermophilic aerobic digestion (ATAD)—Part I: Review of origins, design, and process operation. J. Environ. Eng. Sci. 2007, 6, 665–678. [Google Scholar] [CrossRef]
- Piterina, A.V.; Bartlett, J.; Pembroke, T.J. Evaluation of the removal of indicator bacteria from domestic sludge processed by autothermal thermophilic aerobic digestion (ATAD). Int. J. Environ. Res. Public Health 2010, 7, 3422–3441. [Google Scholar] [CrossRef]
- Liu, S.; Yang, X.; Yao, X. Effects of pH on the biodegradation characteristics of thermophilic micro-aerobic digestion for sludge stabilization. RSC Adv. 2019, 9, 8379–8388. [Google Scholar] [CrossRef]
- Beaudet, R.; Gagnon, C.; Bisaillon, J.G.; Ishaque, M. Microbiological aspects of aerobic thermophilic treatment of swine waste. Appl. Environ. Microbiol. 1990, 56, 971–976. [Google Scholar]
- Shi, Y.; Ren, H. VSS Degradation Kinetics in High Temperature Aerobic Digestion and Microbial Community Characteristics. J. Chem. 2018, 2018, 8131820. [Google Scholar] [CrossRef]
- Dhanarani Sridevi, T.; Shankar, C.; Prakash, P.; Poornima Priyadharshani, T.K.; Thamaraiselvi, K. Conversion of poultry litter into class A biosolids using autothermal thermophilic aerobic digestion: A remedy for solid waste management. Manag. Environ. Qual. An Int. J. 2016, 27, 4–14. [Google Scholar] [CrossRef]
- Piterina, A.V.; Pembroke, J.T. Use of PCR-DGGE Based Molecular Methods to Analyse Microbial Community Diversity and Stability during the Thermophilic Stages of an ATAD Wastewater Sludge Treatment Process as an Aid to Performance Monitoring. ISRN Biotechnol. 2013, 2013, 162645. [Google Scholar] [CrossRef] [PubMed]
- Couillard, D.; Zhu, S. Thermophilic aerobic process for the treatment of slaughterhouse effluents with protein recovery. Environ. Pollut. 1993, 79, 121–126. [Google Scholar] [CrossRef]
- Zvauya, R.; Parawira, W.; Mawadza, C. Aspects of aerobic thermophilic treatment of Zimbabwean traditional opaque-beer brewery wastewater. Bioresour. Technol. 1994, 48, 273–274. [Google Scholar] [CrossRef]
- LaPara, T.M.; Nakatsu, C.H.; Pantea, L.M.; Alleman, J.E. Stability of the bacterial communities supported by a seven-stage biological process treating pharmaceutical wastewater as revealed by PCR-DGGE. Water Res. 2002, 36, 638–646. [Google Scholar] [CrossRef]
- Rintala, J.; Lepisto, R. Thermophilic anaerobic-aerobic and aerobic treatment of kraft bleaching effluents. Water Sci. Technol. 1993, 28, 11–16. [Google Scholar] [CrossRef]
- Hayes, D.; Izzard, L.; Seviour, R. Microbial ecology of autothermal thermophilic aerobic digester (ATAD) systems for treating waste activated sludge. Syst. Appl. Microbiol. 2011, 34, 127–138. [Google Scholar] [CrossRef]
- Piterina, A.V.; Bartlett, J.; Tony Pembroke, J. Molecular analysis of bacterial community DNA in sludge undergoing autothermal thermophilic aerobic digestion (ATAD): Pitfalls and improved methodology to enhance diversity recovery. Diversity 2010, 2, 505–526. [Google Scholar] [CrossRef]
- Juteau, P.; Tremblay, D.; Villemur, R.; Bisaillon, J.G.; Beaudet, R. Analysis of the bacterial community inhabiting an aerobic thermophilic sequencing batch reactor (AT-SBR) treating swine waste. Appl. Microbiol. Biotechnol. 2004, 66, 115–122. [Google Scholar] [CrossRef]
- Liu, S.; Song, F.; Zhu, N.; Yuan, H.; Cheng, J. Chemical and microbial changes during autothermal thermophilic aerobic digestion (ATAD) of sewage sludge. Bioresour. Technol. 2010, 101, 9438–9444. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Asakura, Y.; Kanda, K.; Fukui, R.; Kawano, Y.; Okugawa, Y.; Tashiro, Y.; Sakai, K. Dynamic bacterial community changes in the autothermal thermophilic aerobic digestion process with cell lysis activities, shaking and temperature increase. J. Biosci. Bioeng. 2018, 126, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Sonnleitner, B.; Fiechter, A. Bacterial diversity in thermophilic aerobic sewage sludge–II. Types of organisms and their capacities. Eur. J. Appl. Microbiol. Biotechnol. 1983, 18, 174–180. [Google Scholar] [CrossRef]
- Gilbride, K.A.; Lee, D.Y.; Beaudette, L.A. Molecular techniques in wastewater: Understanding microbial communities, detecting pathogens, and real-time process control. J. Microbiol. Methods 2006, 66, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Storm, P.F. Effect of temperature on bacterial species diversity in thermophilic solid–waste composting. Appl. Environ. Microbiol. 1985, 50, 899–905. [Google Scholar]
- Bomio, M.; Sonnleitner, B.; Fiechter, A. Growth and biocatalytic activities of aerobic thermophilic populations in sewage sludge. Appl. Microbiol. Biotechnol. 1989, 32, 356–362. [Google Scholar] [CrossRef]
- Sonnleitner, B.; Bomio, M. Physiology and performance of thermophilic microorganisms in sewage sludge treatment processes. Biodegradation 1990, 1, 133–146. [Google Scholar] [CrossRef]
- Amann, R.; Lemmer, H.; Wagner, M. Monitoring the community structure of wastewater treatment plants: A comparison of old and new techniques. FEMS Microbiol. Ecol. 1998, 25, 205–215. [Google Scholar] [CrossRef]
- McCaig, A.E.; Grayston, S.J.; Prosser, J.I.; Glover, L.A. Impact of cultivation on characterisation of species composition of soil bacterial communities. FEMS Microbiol. Ecol. 2001, 35, 37–48. [Google Scholar] [CrossRef]
- Marshall, K.C.; Cruickshank, R.H. Cell surface hydrophobicity and the orientation of certain bacteria at interfaces. Arch. Mikrobiol. 1973, 91, 29–40. [Google Scholar] [CrossRef]
- Dudman, W.F. The role of surface polysaccharide in natural environments. In Surface Carbohydrates of the Prokaryotic Cell; Academic Press: London, UK, 1977; pp. 357–414. [Google Scholar]
- Piterina, A.V.; Bartlett, J.; Pembroke, J.T. Morphological characterisation of ATAD thermophilic sludge; sludge ecology and settleability. Water Res. 2011, 45, 3427–3438. [Google Scholar] [CrossRef] [PubMed]
- Pike, E.B.; Carrington, E.G.; Ashburner, P.A. An Evaluation of Procedures for Enumerating Bacteria in Activated Sludge. J. Appl. Bacteriol. 1972, 35, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Kell, D.B.; Kaprelyants, A.S.; Weichart, D.H.; Harwood, C.R.; Barer, M.R. Viability and activity in readily culturable bacteria: A review and discussion of the practical issues. Antonie van Leeuwenhoek. Int. J. Gen. Mol. Microbiol. 1998, 73, 169–187. [Google Scholar]
- Vorgias, C.E.; Anitranikian, G.A. Extremophiles: pH, Temperature and salinity. In Microbial Diversity and Bioprospecting; Bull, A.B., Ed.; ASM Press: Washington, DC, USA, 2003. [Google Scholar]
- Zengler, K.; Toledo, G.; Rappé, M.; Elkins, J.; Mathur, E.; Short, J.M.; Keller, M.; Rappe, M. Cultivating the uncultured. Proc. Natl. Acad. Sci. USA 2002, 99, 15681–15686. [Google Scholar] [CrossRef] [PubMed]
- Piterina, AV.; Pembroke, J.T. Preparation and Analysis of Environmental DNA: Optimisation of techniques for phylogenetic analysis of ATAD Sludge. In Communicating Current Research and Educational Topics and Trends in Applied Microbiology; Mendez-Vilas, A., Ed.; Formatex: Badajoz, Spain, 2010. [Google Scholar]
- Álvarez, A.J.; Buttner, M.P.; Stetzenbach, L.D. PCR for bioaerosol monitoring: Sensitivity and environmental interference. Appl. Environ. Microbiol. 1995, 61, 3639. [Google Scholar] [PubMed]
- Pomp, D.; Medrano, J.F. Organic solvents as facilitators of polymerase chain reaction. Biotechniques 1991, 10, 58–59. [Google Scholar] [PubMed]
- Rådström, P.; Knutsson, R.; Wolffs, P.; Lövenklev, M.; Löfström, C. Pre-PCR processing: Strategies to generate PCR-compatible samples. Appl. Biochem. Biotechnol. 2004, 26, 133–146. [Google Scholar] [CrossRef]
- Muyzer, G.; De Waal, E.C.; Uitterlinden, A.G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar]
- Marzorati, M.; Wittebolle, L.; Boon, N.; Daffonchio, D.; Verstraete, W. How to get more out of molecular fingerprints: Practical tools for microbial ecology. Environ. Microbiol. 2008, 10, 1571–1581. [Google Scholar] [CrossRef]
- Sabat, G.; Rose, P.; Hickey, W.J.; Harkin, J.M. Selective and sensitive method for PCR amplification of Escherichia coli 16S rRNA genes in soil. Appl. Environ. Microbiol. 2000, 66, 844–849. [Google Scholar] [CrossRef]
- Zwirglmaier, K.; Ludwig, W.; Schleifer, K.H. Improved Method for Polynucleotide Probe-Based Cell Sorting, Using DNA-Coated Microplates. Appl. Environ. Microbiol. 2004, 70, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Da Mota, F.F.; Gomes, E.A.; Paiva, E.; Seldin, L. Assessment of the diversity of Paenibacillus species in environmental samples by a novel rpoB-based PCR-DGGE method. FEMS Microbiol. Ecol. 2005, 53, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Morrison, M. Comparisons of different hypervariable regions of rrs genes for use in fingerprinting of microbial communities by PCR-denaturing gradient gel electrophoresis. Appl. Environ. Microbiol. 2004, 70, 4800–4806. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.F.; Lu, C.J.; Sung, L.K.; Wu, Y.S. Full-scale evaluation of heat balance for autothermal thermophilic aerobic treatment of food processing wastewater. Water Sci. Technol. 2001, 43, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Korbie, D.J.; Mattick, J.S. Touchdown PCR for increased specificity and sensitivity in PCR amplification. Nat. Protoc. 2008, 3, 1452–1456. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.M.; Cho, H.U.; Park, K.Y.; Park, H.; Cho, K.H. Identification of the bacterial community of a pilot-scale thermophilic aerobic bioreactor treating sewage sludge. Int. Biodeterior. Biodegrad. 2014, 92, 66–70. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Shokoohi, R.; Rahmani, A.; Asgari, G.; Dargahi, A.; Vaziri, Y.; Abbasi, M.A. Evaluation of autothermal thermophilic aerobic digester performance for the stabilization of municipal wastewater sludge. Pak. J. Biol. Sci. 2017, 20, 260–266. [Google Scholar] [PubMed]
- Wang, P.; Ota, M.; Tanaka, M. Thermal balance analysis of autothermal thermophilic aerobic treatment of human excreta. Eng. Agric. Environ. Food 2014, 7, 1–6. [Google Scholar] [CrossRef]
- Bornscheuer, U.T. Microbial carboxyl esterases: Classification, properties and application in biocatalysis. FEMS Microbiol. Rev. 2002, 26, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Dodds, W.K.; Banks, M.K.; Skalsky, J.; Strauss, E.A. Optimal staining and sample storage time for direct microscopic enumeration of total and active bacteria in soil with two fluorescent dyes. Appl. Environ. Microbiol. 1995, 61, 3367–3372. [Google Scholar] [PubMed]
- Piterina, A.V.; Barlett, J.; Pembroke, J.T. 13C-NMR assessment of the pattern of organic matter transformation during domestic wastewater treatment by autothermal aerobic digestion (ATAD). Int. J. Environ. Res. Public Health 2009, 6, 2288–2306. [Google Scholar] [CrossRef] [PubMed]
- Smernik, R.J.; Oliver, I.W.; Merrington, G. Characterization of Sewage Sludge Organic Matter Using Solid-State Carbon-13 Nuclear Magnetic Resonance Spectroscopy. J. Environ. Qual. 2003, 32, 1516–1522. [Google Scholar] [CrossRef] [PubMed]
- Smernik, R.J.; Oliver, I.W.; Merrington, G. Advanced Solid-State Carbon-13 Nuclear Magnetic Resonance Spectroscopic Studies of Sewage Sludge Organic Matter. J. Environ. Qual. 2003, 32, 1523–1533. [Google Scholar] [CrossRef] [PubMed]
- Maillard, L.C. Reaction of amino acids on sugars: Melanoidin formation through a methodologic way. Comptes Rendus De L’Académie Des Sci. 1912, 156, 148–149. [Google Scholar]
- Zhou, J.; Mavinic, D.S.; Kelly, H.G.; Ramey, W.D. Effects of temperatures and extracellular proteins on dewaterability of thermophilically digested biosolids. J. Environ. Eng. Sci. 2003, 1, 409–415. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, H.H.P. Influences of extracellular polymeric substances (EPS) on flocculation, settling, and dewatering of activated sludge. Crit. Rev. Environ. Sci. Technol. 2003, 33, 237–273. [Google Scholar] [CrossRef]
- Vladova, R.K.; Vaklieva-Bancheva, N.G.; Kirilova, E.G. Improving the energy efficiency of the ATAD system through redesign using integration superstructure. Chem. Eng. Trans. 2018, 70, 1021–1026. [Google Scholar]



{kind=link}
{kind=link}
| Waste Type | Reference |
|---|---|
| Livestock manure | [11] |
| Piggery waste | [12] |
| Poultry litter | [13] |
| Domestic municipal waste | [14] |
| Slaughter house wastes | [15] |
| Brewery waste | [16] |
| Pharmaceutical wastes | [17] |
| Paper waste | [18] |
| Food waste | [4] |
| Fermentation waste | [5] |
| DNA Extraction | Eukarya | Archaea | Bacteria | Fungi |
|---|---|---|---|---|
| Sludge inlet | + | + | + | + |
| Reactor 1 40 °C | − | − | + | + |
| Reactor 2 70 °C | − | − | + | − |
| Sludge storage day 1 | − | − | + | − |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pembroke, J.T.; Ryan, M.P. Autothermal Thermophilic Aerobic Digestion (ATAD) for Heat, Gas, and Production of a Class A Biosolids with Fertilizer Potential. Microorganisms 2019, 7, 215. https://doi.org/10.3390/microorganisms7080215
Pembroke JT, Ryan MP. Autothermal Thermophilic Aerobic Digestion (ATAD) for Heat, Gas, and Production of a Class A Biosolids with Fertilizer Potential. Microorganisms. 2019; 7(8):215. https://doi.org/10.3390/microorganisms7080215
Chicago/Turabian StylePembroke, J. Tony, and Michael P. Ryan. 2019. "Autothermal Thermophilic Aerobic Digestion (ATAD) for Heat, Gas, and Production of a Class A Biosolids with Fertilizer Potential" Microorganisms 7, no. 8: 215. https://doi.org/10.3390/microorganisms7080215
APA StylePembroke, J. T., & Ryan, M. P. (2019). Autothermal Thermophilic Aerobic Digestion (ATAD) for Heat, Gas, and Production of a Class A Biosolids with Fertilizer Potential. Microorganisms, 7(8), 215. https://doi.org/10.3390/microorganisms7080215