Functional Genomics of Aspergillus oryzae: Strategies and Progress

Abstract
1. Introduction
2. Genome Sequencing of A. oryzae
3. Strategies for Functional Genomics of A. oryzae
3.1. Selection Markers
3.1.1. Drug Resistance Markers
3.1.2. Auxotrophic Markers
3.2. Strategies for the Transformation of A. oryzae
3.2.1. Protoplast-Mediated Transformation
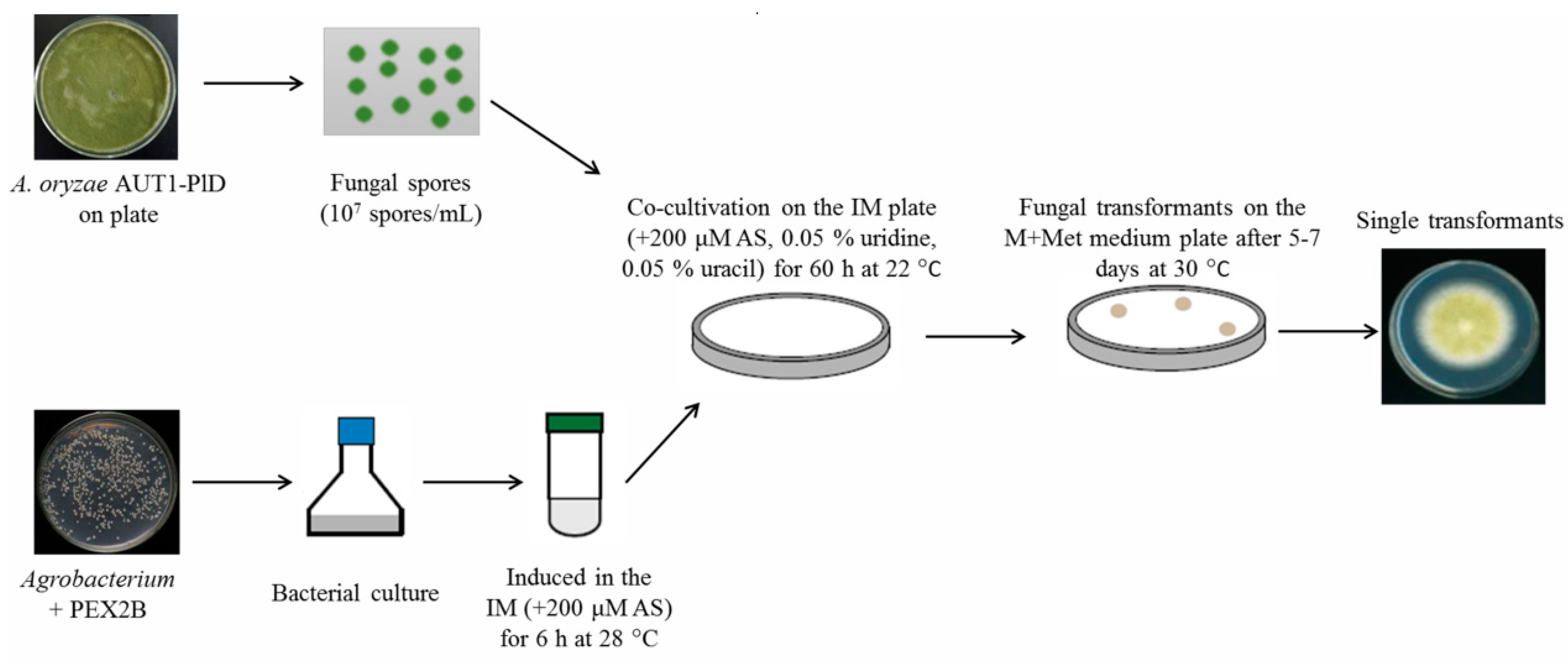
3.2.2. Agrobacterium-Mediated Transformation
3.2.3. Electroporation
3.3. Genetic Manipulations
4. Advances in the Functional Genomics of A. oryzae
4.1. Conidiation
4.2. Protein Secretion and Expression
4.3. Secondary Metabolites
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- He, B.; Hu, Z.; Ma, L.; Li, H.; Ai, M.; Han, J.; Zeng, B. Transcriptome analysis of different growth stages of Aspergillus oryzae reveals dynamic changes of distinct classes of genes during growth. BMC Microbiol. 2018, 18, 12. [Google Scholar] [CrossRef] [PubMed]
- Machida, M.; Yamada, O.; Gomi, K. Genomics of Aspergillus oryzae: Learning from the history of Koji mold and exploration of its future. DNA Res. 2008, 15, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Lu, X.; Xing, L.; Ho, S.W.A.; Kwan, H.S. Genomic and transcriptomic comparison of Aspergillus oryzae strains: A case study in soy sauce koji fermentation. J. Ind. Microbiol. Biotechnol. 2018, 45, 839–853. [Google Scholar] [CrossRef]
- Brandl, J.; Andersen, M.R. Aspergilli: Models for Systems Biology in Filamentous Fungi. Curr. Opin. Syst. Biol. 2017, 6, 67–73. [Google Scholar] [CrossRef]
- Machida, M.; Asai, K.; Sano, M.; Tanaka, T.; Kumagai, T.; Terai, G.; Kusumoto, K.I.; Arima, T.; Akita, O.; Kashiwagi, Y. Genome sequencing and analysis of Aspergillus oryzae. Nature 2005, 438, 1157. [Google Scholar] [CrossRef]
- He, B.; Tu, Y.; Hu, Z.; Ma, L.; Dai, J.; Cheng, X.; Li, H.; Liu, L.; Zeng, B. Genome-wide identification and expression profile analysis of the HOG gene family in Aspergillus oryzae. World J. Microbiol. Biotechnol. 2018, 34, 35. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Yao, Y.; Qi, W.; Wang, C.; Hou, L.; Zeng, B.; Cao, X. Draft genome sequence of Aspergillus oryzae strain 3.042. Eukaryot. Cell 2012, 11, 1178. [Google Scholar] [CrossRef]
- Lin, W.; Song, J.; Hu, W.; Miao, J.; Gao, X. Relationship Between Extracellular Cellulase, Pectinase and Xylanase Activity of Isolated Aspergillus oryzae Strains Grown on Koji and the Umami-Tasting Amino Acid Content of Soy Sauce. Food Biotechnol. 2016, 30, 278–291. [Google Scholar] [CrossRef]
- Umemura, M.; Koike, H.; Yamane, N.; Koyama, Y.; Satou, Y.; Kikuzato, I.; Teruya, M.; Tsukahara, M.; Imada, Y.; Wachi, Y.; et al. Comparative Genome Analysis Between Aspergillus oryzae Strains Reveals Close Relationship Between Sites of Mutation Localization and Regions of Highly Divergent Genes among Aspergillus Species. DNA Res. 2012, 19, 375–382. [Google Scholar] [CrossRef]
- Zhao, G.; Yao, Y.; Hou, L.; Wang, C.; Cao, X. Draft Genome Sequence of Aspergillus oryzae 100-8, an Increased Acid Protease Production Strain. Genome Announc. 2014, 2, e00548-14. [Google Scholar] [CrossRef] [PubMed]
- Thammarongtham, C.; Nookaew, I.; Vorapreeda, T.; Srisuk, T.; Land, M.L.; Jeennor, S.; Laoteng, K. Genome Characterization of Oleaginous Aspergillus oryzae BCC7051: A Potential Fungal-Based Platform for Lipid Production. Curr. Microbiol. 2018, 75, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Pomraning, K.R.; Bohutskyi, P.; Magnuson, J.K. Draft Genome Sequence of Aspergillus oryzae ATCC 12892. Genome Announc. 2018, 6, e00251-18. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.Q.; Moyer, D.L.; Maiyuran, S.; Labaro, A.; Brody, H. Vector-initiated transitive RNA interference in the filamentous fungus Aspergillus oryzae. Fungal Genet. Biol. 2012, 49, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Tada, S.; Fukuoka, M.; Taketani, H.; Tsukakoshi, Y.; Matsushita, M.; Oda, K.; Kusumoto, K.-I.; Kashiwagi, Y.; Sugiyama, M. A novel transformation system using a bleomycin resistance marker with chemosensitizers for Aspergillus oryzae. Biochem. Biophys. Res. Commun. 2009, 383, 42–47. [Google Scholar] [CrossRef]
- Chou, T.H.; Tzean, S.S. Protoplasting, regeneration and transformation of medicinal mushroom Ganoderma multipileum using succinate dehydrogenase mutation gene as a selection marker. Ann. Microbiol. 2016, 66, 111–120. [Google Scholar] [CrossRef]
- Zhang, S.; Ban, A.; Ebara, N.; Mizutani, O.; Tanaka, M.; Shintani, T.; Gomi, K. Self-excising Cre/mutant lox marker recycling system for multiple gene integrations and consecutive gene deletions in Aspergillus oryzae. J. Biosci. Bioeng. 2017, 123, 403–411. [Google Scholar] [CrossRef]
- Newsome, R.; Tran, N.; Paoli, G.M.; Jaykus, L.A.; Tompkin, B.; Miliotis, M.; Ruthman, T.; Hartnett, E.; Busta, F.F.; Petersen, B. Development of a risk-ranking framework to evaluate potential high-threat microorganisms, toxins, and chemicals in food. J. Food Sci. 2010, 74, R39–R45. [Google Scholar] [CrossRef]
- Zhu, L.; Maruyama, J.; Kitamoto, K. Further enhanced production of heterologous proteins by double-gene disruption (ΔAosedD ΔAovps10) in a hyper-producing mutant of Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2013, 97, 6347–6357. [Google Scholar] [CrossRef]
- Yoshino-Yasuda, S.; Mori, A.; Ishihara, N.; Hasegawa, O.; Kato, M.; Kitamoto, N. Development of a Highly Efficient Gene Replacement System for an Industrial Strain of Aspergillus oryzae Used in the Production of Miso, a Japanese Fermented Soybean Paste. Food Sci. Technol. Int. Tokyo 2011, 17, 161–166. [Google Scholar] [CrossRef][Green Version]
- Nguyen, K.T.; Ho, Q.N.; Do, L.T.B.X.; Mai, L.T.D.; Pham, D.N.; Tran, H.T.T.; Le, D.H.; Nguyen, H.Q.; Tran, V.T. A new and efficient approach for construction of uridine/uracil auxotrophic mutants in the filamentous fungus Aspergillus oryzae using Agrobacterium tumefaciens-mediated transformation. World J. Microb. Biotechnol. 2017, 33, 107. [Google Scholar] [CrossRef]
- Maruyama, J.I.; Kitamoto, K. Multiple gene disruptions by marker recycling with highly efficient gene-targeting background (ΔligD) in Aspergillus oryzae. Biotechnol. Lett. 2008, 30, 1811–1817. [Google Scholar] [CrossRef]
- Yoon, J.; Maruyama, J.; Kitamoto, K. Disruption of ten protease genes in the filamentous fungus Aspergillus oryzae highly improves production of heterologous proteins. Appl. Microbiol. Biotechnol. 2011, 89, 747–759. [Google Scholar] [CrossRef]
- Sun, Y.; Niu, Y.; He, B.; Ma, L.; Li, G.; Tran, V.T.; Zeng, B.; Hu, Z. A dual selection marker transformation system using Agrobacterium tumefaciens for the industrial Aspergillus oryzae 3.042. J. Microbiol. Biotechnol. 2018, 29, 230–234. [Google Scholar] [CrossRef]
- Nguyen, K.T.; Ho, Q.N.; Pham, T.H.; Phan, T.N.; Tran, V.T. The construction and use of versatile binary vectors carrying pyrG auxotrophic marker and fluorescent reporter genes for Agrobacterium -mediated transformation of Aspergillus oryzae. World J. Microbiol. Biotechnol. 2016, 32, 204. [Google Scholar] [CrossRef]
- Tamano, K.; Bruno, K.S.; Koike, H.; Ishii, T.; Ai, M.; Umemura, M.; Culley, D.E.; Baker, S.E.; Machida, M. Increased production of free fatty acids in Aspergillus oryzae by disruption of a predicted acyl-CoA synthetase gene. Appl. Microbiol. Biotechnol. 2015, 99, 3103. [Google Scholar] [CrossRef]
- Jin, F.J.; Maruyama, J.; Juvvadi, P.R.; Arioka, M.; Kitamoto, K. Development of a novel quadruple auxotrophic host transformation system by argB gene disruption using adeA gene and exploiting adenine auxotrophy in Aspergillus oryzae. FEMS Microbiol. Lett. 2010, 239, 79–85. [Google Scholar] [CrossRef]
- Nakamura, H.; Katayama, T.; Okabe, T.; Iwashita, K.; Fujii, W.; Kitamoto, K.; Maruyama, J.I. Highly efficient gene targeting in Aspergillus oryzae industrial strains under ligD mutation introduced by genome editing: Strain-specific differences in the effects of deleting EcdR, the negative regulator of sclerotia formation. J. Gen. Appl. Microbiol. 2017, 63, 172–178. [Google Scholar] [CrossRef]
- Jin, F.J.; Maruyama, J.; Juvvadi, P.R.; Arioka, M.; Kitamoto, K. Adenine auxotrophic mutants of Aspergillus oryzae: Development of a novel transformation system with triple auxotrophic hosts. Biosci. Biotechnol. Biochem. 2004, 68, 656–662. [Google Scholar] [CrossRef]
- Nemoto, T.; Watanabe, T.; Mizogami, Y.; Maruyama, J.-i.; Kitamoto, K. Isolation of Aspergillus oryzae mutants for heterologous protein production from a double proteinase gene disruptant. Appl. Microbiol. Biotechnol. 2009, 82, 1105. [Google Scholar] [CrossRef]
- Jin, F.J.; Katayama, T.; Maruyama, J.I.; Kitamoto, K. Comparative genomic analysis identified a mutation related to enhanced heterologous protein production in the filamentous fungus Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2016, 100, 9163–9174. [Google Scholar] [CrossRef]
- Escaño, C.S.; Juvvadi, P.R.; Jin, F.J.; Takahashi, T.; Koyama, Y.; Yamashita, S.; Maruyama, J.; Kitamoto, K. Disruption of the Aopex11-1 gene involved in peroxisome proliferation leads to impaired Woronin body formation in Aspergillus oryzae. Eukaryot. Cell 2009, 8, 296–305. [Google Scholar] [CrossRef]
- Jiang, D.; Zhu, W.; Wang, Y.; Sun, C.; Zhang, K.-Q.; Yang, J. Molecular tools for functional genomics in filamentous fungi: Recent advances and new strategies. Biotechnol. Adv. 2013, 31, 1562–1574. [Google Scholar] [CrossRef]
- Weyda, I.; Yang, L.; Vang, J.; Ahring, B.K.; Lübeck, M.; Lübeck, P.S. A comparison of Agrobacterium—Mediated transformation and protoplast-mediated transformation with CRISPR-Cas9 and bipartite gene targeting substrates, as effective gene targeting tools for Aspergillus carbonarius. J. Microbiol. Methods 2017, 135, 26–34. [Google Scholar] [CrossRef]
- Murakami, K.; Aikawa, J.I.; Horinouchi, S.; Beppu, T. Characterization of an Aspartic Proteinase of Mucor pusillus expressed in Aspergillus oryzae. Mol. Gen. Genet. 1993, 241, 312–318. [Google Scholar] [CrossRef]
- Tada, S.; Matsushita-Morita, M.; Suzuki, S.; Kusumoto, K.I.; Kashiwagi, Y. Characterization of a neutral ceramidase orthologue from Aspergillus oryzae. FEMS Microbiol. Lett. 2010, 298, 157–165. [Google Scholar] [CrossRef]
- Williams, J.A. Strain Engineering, 1st ed.; Humana Press: New York, NY, USA, 2011; pp. 447–455. [Google Scholar]
- Wang, S.; Chen, H.; Tang, X.; Zhang, H.; Chen, W.; Chen, Y.Q. Molecular tools for gene manipulation in filamentous fungi. Appl. Microbiol. Biotechnol. 2017, 101, 8063–8075. [Google Scholar] [CrossRef]
- Ulrich, K.; Birgit, H. New tools for the genetic manipulation of filamentous fungi. Appl. Microbiol. Biotechnol. 2010, 86, 51–62. [Google Scholar]
- Van den Berg, M.A.; Maruthachalam, K. (Eds.) Genetic Transformation Systems in Fungi, Volume 1; Springer Nature: Basel, Switzerland, 2015; pp. 8–15. [Google Scholar]
- Li, S.; Cong, Y.; Liu, Y.; Wang, T.; Shuai, Q.; Chen, N.; Gai, J.; Li, Y. Optimization of Agrobacterium-mediated Transformation in Soybean. Front. Plant Sci. 2017, 8, 246. [Google Scholar] [CrossRef]
- Piers, K.L.; Heath, J.D.; Liang, X.; Stephens, K.M.; Nester, E.W. Agrobacterium tumefaciens-mediated Transformation of Yeast. Proc. Natl. Acad. Sci. USA 1996, 93, 1613–1618. [Google Scholar] [CrossRef]
- Wang, D.; Dan, H.; Li, G.; Song, G.; Lv, H.; Shan, Q.; Li, W. An efficient tool for random insertional mutagenesis: Agrobacterium tumefaciens-mediated transformation of the filamentous fungus Aspergillus terreus. J. Microbiol. Methods 2014, 98, 114–118. [Google Scholar] [CrossRef]
- Singh, R.K.; Prasad, M. Advances in Agrobacterium tumefaciens-mediated genetic transformation of graminaceous crops. Protoplasma 2016, 253, 691–707. [Google Scholar] [CrossRef]
- Govender, N.T.; Wong, M.Y. Detection of oil palm root penetration by Agrobacterium-mediated transformed Ganoderma boninense expressing green fluorescent protein. Phytopathology 2017, 107, 483–490. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, R.; Mao, W.J.; Gong, M.; Gao, Y.N.; Tang, L.H.; Yang, R.F.; Li, Y.; Zhou, C.L.; Bao, D.P. A simple and efficient transformation system for the edible mushroom Pleurotus eryngii. Mycoscience 2016, 57, 356–360. [Google Scholar] [CrossRef]
- Madhavan, A.; Pandey, A.; Sukumaran, R.K. Expression system for heterologous protein expression in the filamentous fungus Aspergillus unguis. Bioresour. Technol. 2017, 245, 1334–1342. [Google Scholar] [CrossRef]
- Liu, H.; Jiao, X.; Wang, Y.; Yang, X.; Sun, W.; Wang, J.; Zhang, S.; Zhao, Z.K. Fast and efficient genetic transformation of oleaginous yeast Rhodosporidium toruloides by using electroporation. FEMS Yeast Res. 2017, 17, fox017. [Google Scholar] [CrossRef]
- Kotnik, T.; Frey, W.; Sack, M.; Meglič, S.H.; Peterka, M.; Miklavčič, D. Electroporation-based applications in biotechnology. Trends Biotechnol. 2015, 33, 480–488. [Google Scholar] [CrossRef]
- Xu, D.; Pan, L.; Zhao, H.; Zhao, M.; Sun, J.; Liu, D. Breeding and identification of novel koji molds with high activity of acid protease by genome recombination between Aspergillus oryzae and Aspergillus niger. J. Ind. Microbiol. Biotechnol. 2011, 38, 1255–1265. [Google Scholar] [CrossRef]
- Bocking, S.P.; Wiebe, M.G.; Robson, G.D.; Hansen, K.; Christiansen, L.H.; Trinci, A.P. Effect of branch frequency in Aspergillus oryzae on protein secretion and culture viscosity. Biotechnol. Bioeng. 2015, 65, 638–648. [Google Scholar] [CrossRef]
- Yamada, O.; Ohkita, R.I. Gene Silencing by RNA Interference in the Koji Mold Aspergillus oryzae. Biosci. Biotechnol. Biochem. 2007, 71, 138–144. [Google Scholar] [CrossRef][Green Version]
- Nødvig, C.S.; Nielsen, J.B.; Kogle, M.E.; Mortensen, U.H. A CRISPR-Cas9 System for Genetic Engineering of Filamentous Fungi. PLoS ONE 2015, 10, e0133085. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Abdel-Ghany, S.E.; Ali, G.S. Genome editing approaches: manipulating of lovastatin and taxol synthesis of filamentous fungi by CRISPR/Cas9 system. Appl. Microbiol. Biotechnol. 2017, 101, 3953–3976. [Google Scholar] [CrossRef]
- Katayama, T.; Tanaka, Y.; Okabe, T.; Nakamura, H.; Fujii, W.; Kitamoto, K.; Maruyama, J. Development of a genome editing technique using the CRISPR/Cas9 system in the industrial filamentous fungus Aspergillus oryzae. Biotechnol. Lett. 2015, 38, 637–642. [Google Scholar] [CrossRef]
- Kitamoto, K. Cell biology of the Koji mold Aspergillus oryzae. J. Agric Chem. Soc. Japan 2015, 79, 863–869. [Google Scholar]
- Masahiro, O.; Masafumi, T.; Jie, J.F.; Tadashi, T.; Yasuji, K. Genetic analysis of conidiation regulatory pathways in koji-mold Aspergillus oryzae. Fungal Genet. Biol. 2010, 47, 10–18. [Google Scholar]
- Krijgsheld, P.; Bleichrodt, R.; Veluw, G.J.V.; Wang, F.; Müller, W.H.; Dijksterhuis, J.; Wösten, H.A.B. Development in Aspergillus. Stud. Mycol. 2013, 74, 1–29. [Google Scholar] [CrossRef]
- Hatakeyama, R.; Nakahama, T.Y.; Kitamoto, K. Light represses conidiation in koji mold Aspergillus oryzae. J. Agric. Chem. Soc. Japan 2007, 71, 1844–1849. [Google Scholar]
- Nakamura, H.; Kikuma, T.; Jin, F.J.; Maruyama, J.I.; Kitamoto, K. AoRim15 is involved in conidial stress tolerance, conidiation and sclerotia formation in the filamentous fungus Aspergillus oryzae. J. Biosci. Bioeng. 2016, 121, 365–371. [Google Scholar] [CrossRef]
- Fleißner, A.; Dersch, P. Expression and export: recombinant protein production systems for Aspergillus. Appl. Microbiol. Biotechnol. 2010, 87, 1255–1270. [Google Scholar] [CrossRef]
- Yokota, J.I.; Shiro, D.; Tanaka, M.; Onozaki, Y.; Mizutani, O.; Kakizono, D.; Ichinose, S.; Shintani, T.; Gomi, K.; Shintani, T. Cellular responses to the expression of unstable secretory proteins in the filamentous fungus Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2017, 101, 2437–2446. [Google Scholar] [CrossRef]
- Hoang, H.-D.; Maruyama, J.-i.; Kitamoto, K. Modulating endoplasmic reticulum-Golgi cargo receptors for improving secretion of carrier-fused heterologous proteins in the filamentous fungus Aspergillus oryzae. Appl. Environ. Microbiol. 2015, 81, 533–543. [Google Scholar] [CrossRef]
- Yoon, J.; Aishan, T.; Maruyama, J.I.; Kitamoto, K. Enhanced Production and Secretion of Heterologous Proteins by the Filamentous Fungus Aspergillus oryzae via Disruption of Vacuolar Protein Sorting Receptor Gene Aovps10. Appl. Environ. Microbiol. 2010, 76, 5718–5727. [Google Scholar] [CrossRef]
- Yoon, J.; Kikuma, T.; Maruyama, J.; Kitamoto, K. Enhanced Production of Bovine Chymosin by Autophagy Deficiency in the Filamentous Fungus Aspergillus oryzae. PLoS ONE 2013, 8, e62512. [Google Scholar] [CrossRef]
- Nakajima, K.; Asakura, T.; Maruyama, J.; Morita, Y.; Oike, H.; Shimizu-Ibuka, A.; Misaka, T.; Sorimachi, H.; Arai, S.; Kitamoto, K. Extracellular production of neoculin, a sweet-tasting heterodimeric protein with taste-modifying activity, by Aspergillus oryzae. Appl. Environ. Microbiol. 2006, 72, 3716–3723. [Google Scholar] [CrossRef]
- Zhou, M.; Zhou, K.; He, P.; Wang, K.M.; Zhu, R.Z.; Wang, Y.D.; Dong, W.; Li, G.P.; Yang, H.Y.; Ye, Y.Q. Antiviral and Cytotoxic Isocoumarin Derivatives from an Endophytic Fungus Aspergillus oryzae. Planta Med. 2016, 82, 414–417. [Google Scholar] [CrossRef]
- Chankhamjon, P.; Boettgerschmidt, D.; Scherlach, K.; Urbansky, B.; Lackner, G.; Kalb, D.; Dahse, H.M.; Hoffmeister, D.; Hertweck, C. Biosynthesis of the halogenated mycotoxin aspirochlorine in koji mold involves a cryptic amino acid conversion. Angew. Chem. 2015, 53, 13409–13413. [Google Scholar] [CrossRef]
- Nishimura, A.; Okamoto, S.; Yoshizako, F.; Morishima, I.; Ueno, T. Stimulatory effect of acetate and propionate on aspergillic acid formation by Aspergillus oryzae A 21. J. Ferment. Bioeng. 1991, 72, 461–464. [Google Scholar] [CrossRef]
- Marui, J.; Ohashi-Kunihiro, S.; Ando, T.; Nishimura, M.; Koike, H.; Machida, M. Penicillin biosynthesis in Aspergillus oryzae and its overproduction by genetic engineering. J. Biosci. Bioeng. 2010, 110, 8–11. [Google Scholar] [CrossRef]
- Blumenthal, C.Z. Production of toxic metabolites in Aspergillus niger, Aspergillus oryzae, and Trichoderma reesei: Justification of mycotoxin testing in food grade enzyme preparations derived from the three fungi. Regul. Toxicol. Pharmacol. 2004, 39, 214–228. [Google Scholar] [CrossRef]
- Marui, J.; Yamane, N.; Ohashikunihiro, S.; Ando, T.; Terabayashi, Y.; Sano, M.; Ohashi, S.; Ohshima, E.; Tachibana, K.; Higa, Y.; et al. Kojic acid biosynthesis in Aspergillus oryzae is regulated by a Zn(II)2Cys6 transcriptional activator and induced by kojic acid at the transcriptional level. J. Biosci. Bioeng. 2011, 112, 40–43. [Google Scholar] [CrossRef]
- Yamada, R.; Yoshie, T.; Wakai, S.; Asainakashima, N.; Okazaki, F.; Ogino, C.; Hisada, H.; Tsutsumi, H.; Hata, Y.; Kondo, A. Aspergillus oryzae-based cell factory for direct kojic acid production from cellulose. Microb. Cell Fact. 2014, 13, 71. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Møller, L.L.H.; Larsen, T.O.; Kumar, R.; Arnau, J. Safety of the fungal workhorses of industrial biotechnology: Update on the mycotoxin and secondary metabolite potential of Aspergillus niger, Aspergillus oryzae, and Trichoderma reesei. Appl. Microbiol. Biotechnol. 2018, 102, 9481–9515. [Google Scholar] [CrossRef]
- Shaaban, M.; Elmetwally, M.M.; Nasr, H. A new diketopiperazine alkaloid from Aspergillus oryzae. Nat. Prod. Res. 2014, 28, 86–94. [Google Scholar] [CrossRef]
- Yao, G.; Yue, Y.; Fu, Y.; Fang, Z.; Xu, Z.; Ma, G.; Wang, S. Exploration of the Regulatory Mechanism of Secondary Metabolism by Comparative Transcriptomics in Aspergillus flavus. Front. Microbiol. 2018, 9, 1568. [Google Scholar] [CrossRef]
- Yao, G.; Zhang, F.; Nie, X.; Wang, X.; Yuan, J.; Zhuang, Z.; Wang, S. Essential APSES Transcription Factors for Mycotoxin Synthesis, Fungal Development, and Pathogenicity in Aspergillus flavus. Front. Microbiol. 2017, 8, 2277. [Google Scholar] [CrossRef]
- Lee, J.H.; Jo, E.H.; Hong, E.J.; Kim, K.M.; Lee, I. Safety Evaluation of Filamentous Fungi Isolated from Industrial Doenjang Koji. J. Microbiol. Biotechnol. 2014, 24, 1397–1404. [Google Scholar] [CrossRef]
- Tao, L.; Chung, S.H. Non-aflatoxigenicity of commercial Asepergillus oryzae strains due to genetic defects compared to aflatoxigenic Aspergillus flavus. J. Microbiol. Biotechnol. 2014, 24, 1081–1087. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Strains | Assembly | Level | Sequencing Strategies | Size (Mb) | GC% | Gene | Ref. |
|---|---|---|---|---|---|---|---|
| RIB40 | GCA_000184455 | Chromosome | Whole-genome Shotgun | 37.91 | 48.26 | 12,074 | [5] |
| 3042 | GCA_000269785 | Contig | Solexa | 36.57 | 48.28 | 11,639 | [7] |
| AS 3.951 | GCA_000278405 | Contig | Illumina | 36.33 | 48.30 | / | [8] |
| AS 3.863 | GCA_000278425 | Contig | Illumina | 36.40 | 48.30 | / | [8] |
| RIB326 | GCA_000320905 | Contig | SOLiD | 35.42 | 48.40 | / | [9] |
| 100-8 | GCA_000691885 | Contig | Solexa | 36.76 | 48.30 | 11,188 | [10] |
| BCC7051 | GCA_002007945 | Scaffold | PacBio | 38.50 | 47.20 | 11,456 | [11] |
| SRCM101975 | GCA_002214955 | Contig | Ion Torrent | 37.12 | 48.30 | / | [3] |
| SRCM101989 | GCA_002214965 | Contig | Ion Torrent | 36.97 | 48.30 | / | [3] |
| ATCC 12892 | GCA_002894705 | Scaffold | Illumina | 41.16 | 47.60 | / | [12] |
| Wild Strains | Mutant Strains | Selection Marker | Ref. |
|---|---|---|---|
| 3042 | pyrG auxotroph | pyrG− | [23] |
| RIB40 | pyrG auxotroph | pyrG− | [20,24] |
| RIB40 | niaD300 | niaD− | [25] |
| RIB40 | NSR13 | argB−, adeA− | [26] |
| RIB40 | NS4 | ligD, pyrG− | [27] |
| RIB40 | NSR1 | niaD−, SC−, adeA− | [28] |
| RIB40 | NSlD1 | niaD−, SC−, ligD− | [22] |
| RIB40 | NSPlD1 | niaD−, SC−, pyrG−, ligD− | [21] |
| RIB40 | NS-tApE | niaD−, sC−, adeA−, argB− | [29,30] |
| RIB40 | NSRKu70-1-1A | niaD−, SC−, argB−, adeA−, ku70− | [31] |
| Methods | PMT a | AMT b | EP c |
|---|---|---|---|
| Principles | Use cell-wall-degrading enzymes to prepare protoplasts. Uptake the DNA by the addition of PEG and CaCl2. | Agrobacterium tumefaciens is able to transfer the T-DNA region of the Ti plasmid to the genome of A. oryzae. | Uptake DNA is mediated by reversible membrane permeabilization induced by the local application of electric pulses. |
| Advantages | 1. It is easier to get homozygotes because of the large number of receptor cells. 2. Spores, germlings, and mycelium can be used. | 1. Low copy number of inserted DNA. 2. Spores, germlings, and mycelium can be used. 3. Improves targeted integration. | 1. Simple and cheap. 2. Spores and germlings can be used. |
| Disadvantages | 1. The transformation rate is affected by the efficiency of different batches of cell-wall-degrading enzymes. 2. Requires a regeneration procedure. | 1. It is difficult to obtain adequate binary vectors containing the vir genes and the heterologous DNA. 2. Various parameters during co-cultivation affect the transformation rate. | 1. The protocols need to be optimized and are relatively complicated. 2. Protoplast formation is needed. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, B.; Tu, Y.; Jiang, C.; Zhang, Z.; Li, Y.; Zeng, B. Functional Genomics of Aspergillus oryzae: Strategies and Progress. Microorganisms 2019, 7, 103. https://doi.org/10.3390/microorganisms7040103
He B, Tu Y, Jiang C, Zhang Z, Li Y, Zeng B. Functional Genomics of Aspergillus oryzae: Strategies and Progress. Microorganisms. 2019; 7(4):103. https://doi.org/10.3390/microorganisms7040103
Chicago/Turabian StyleHe, Bin, Yayi Tu, Chunmiao Jiang, Zhe Zhang, Yongkai Li, and Bin Zeng. 2019. "Functional Genomics of Aspergillus oryzae: Strategies and Progress" Microorganisms 7, no. 4: 103. https://doi.org/10.3390/microorganisms7040103
APA StyleHe, B., Tu, Y., Jiang, C., Zhang, Z., Li, Y., & Zeng, B. (2019). Functional Genomics of Aspergillus oryzae: Strategies and Progress. Microorganisms, 7(4), 103. https://doi.org/10.3390/microorganisms7040103