Methods and Challenges of Using the Greater Wax Moth (Galleria mellonella) as a Model Organism in Antimicrobial Compound Discovery



Abstract
:1. Introduction
2. Materials and Methods
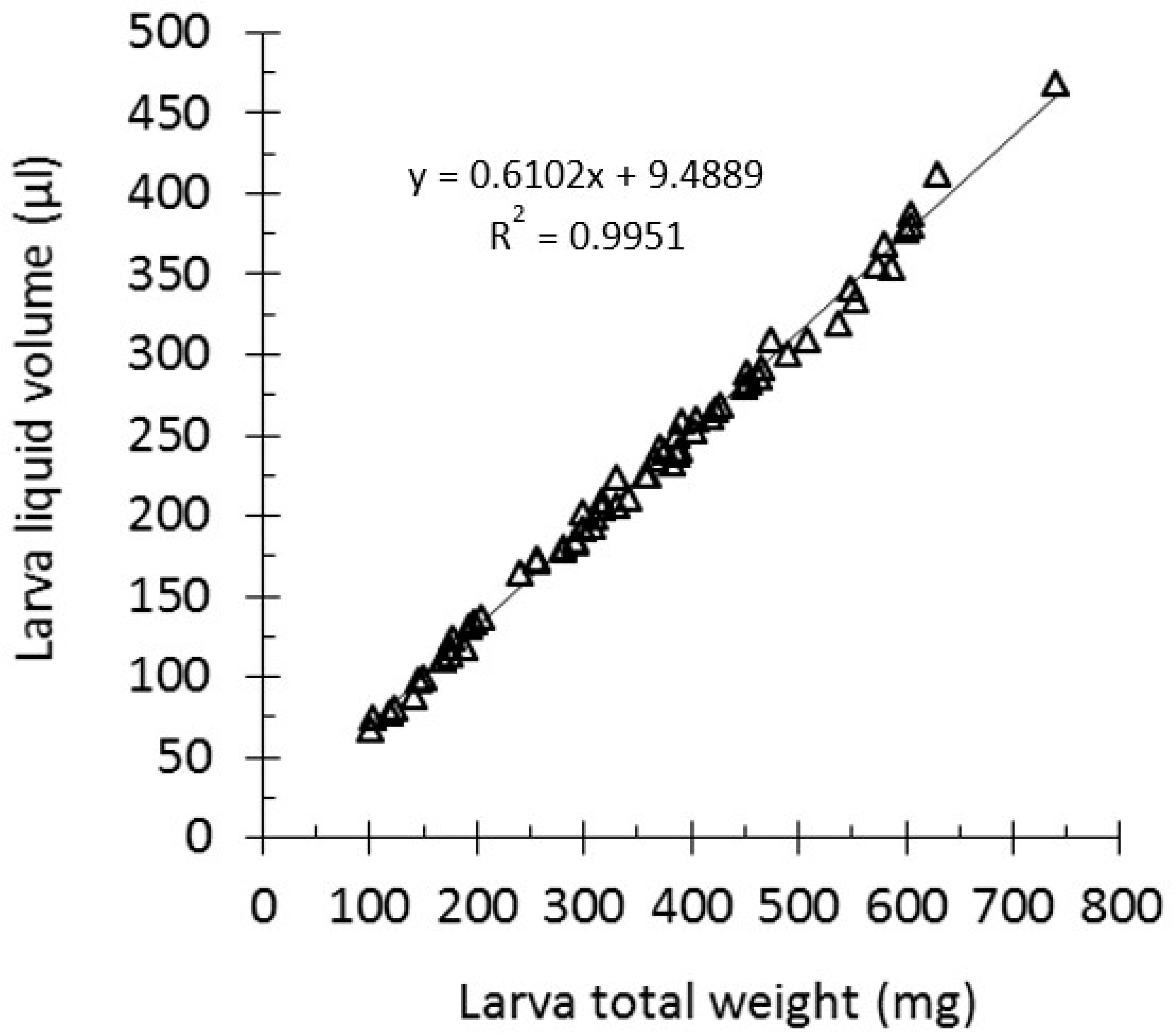
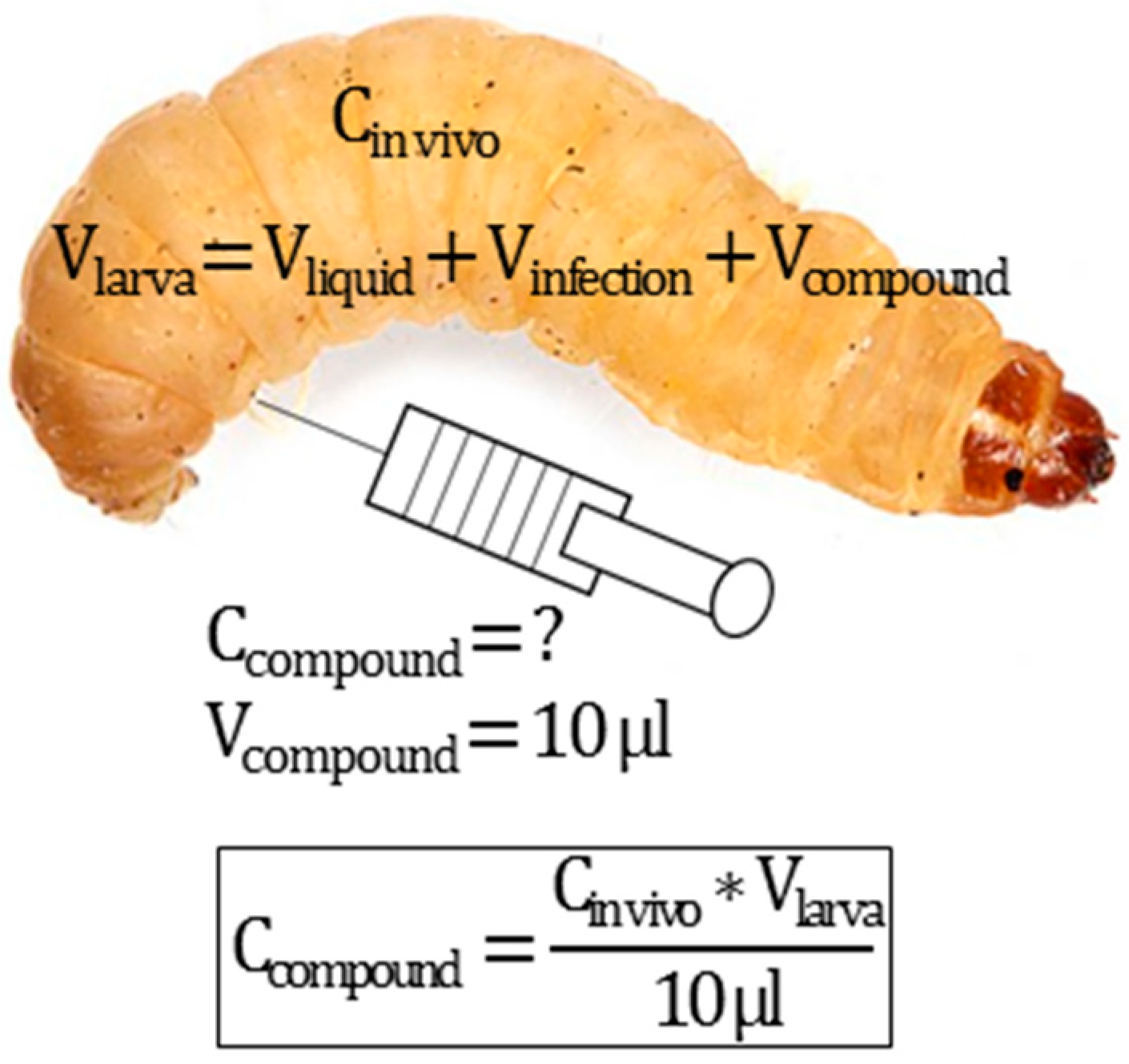
2.1. Galleria mellonella Larvae Liquid Volume Determination
2.2. Bacteria and Growth Conditions
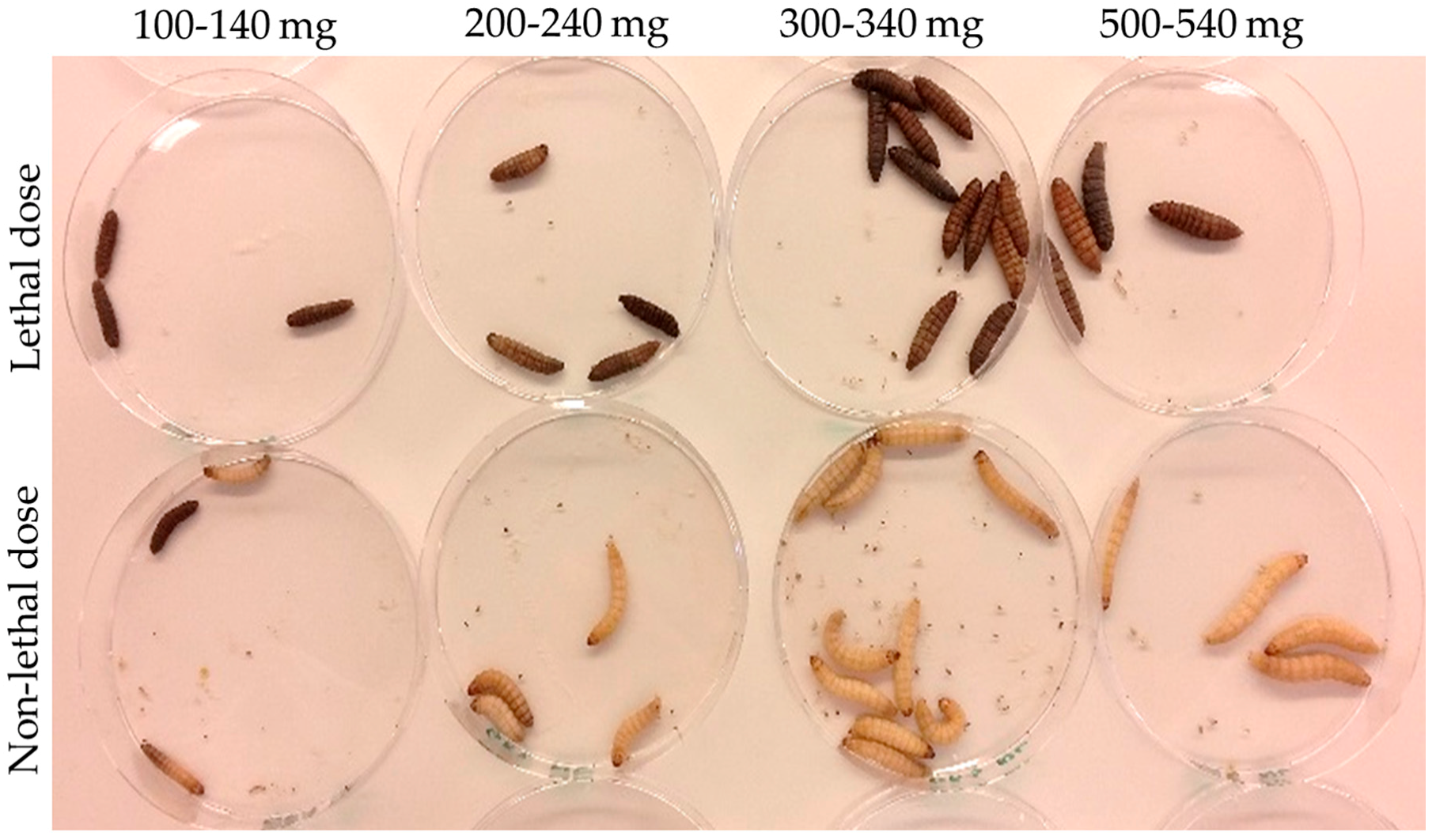
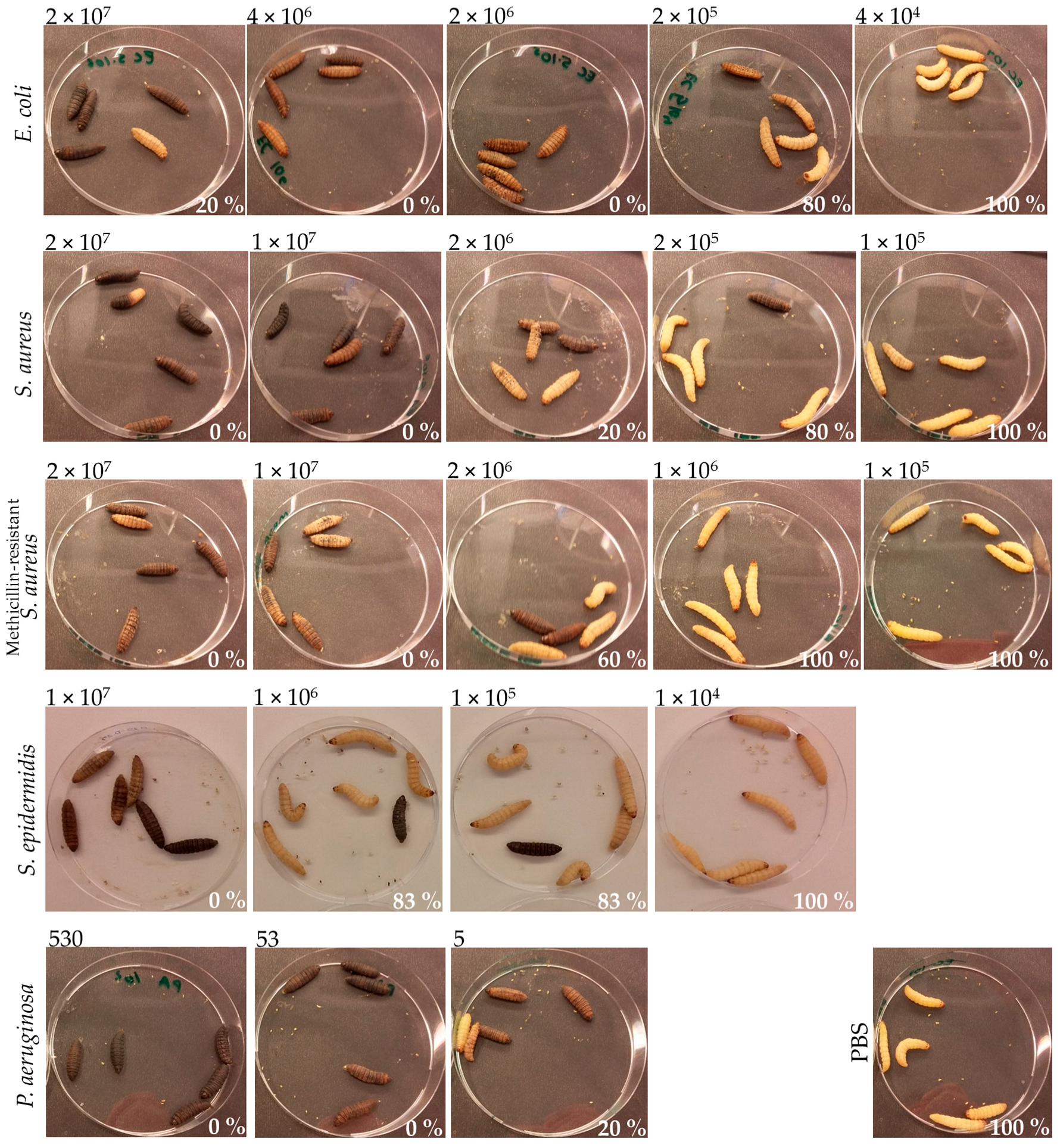
2.3. Galleria mellonella Infection
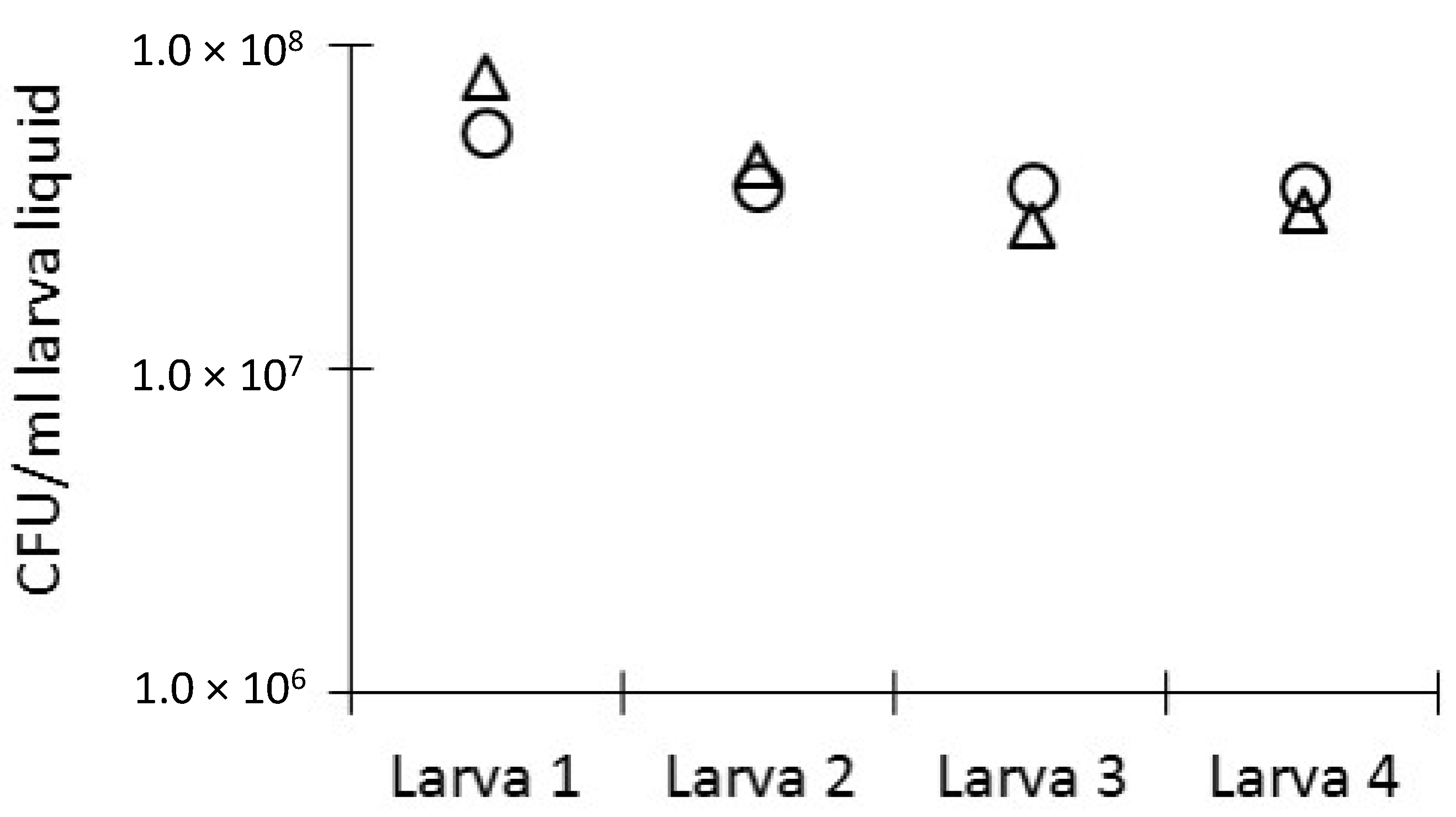
2.4. Bacterial Load in the Haemolymph
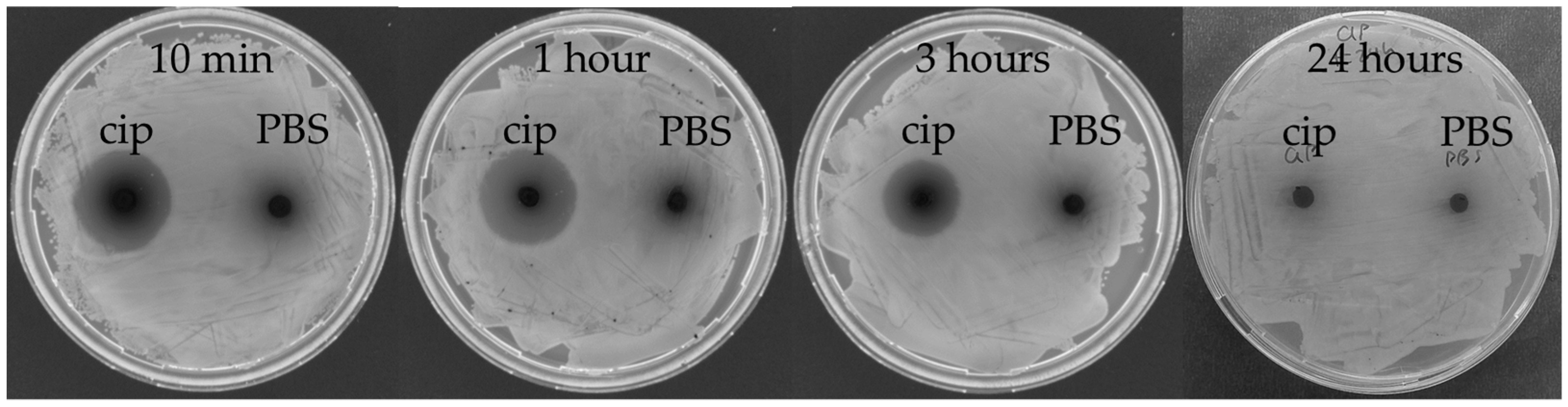
2.5. In Vivo Stability of the Antibiotic
3. Results
3.1. A Linear Regression Curve for G. mellonella Larvae Liquid Volume Determination
3.2. Infection and Treatment Dose Must Be Tuned
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Freires, I.A.; Sardi, J.C.; de Castro, R.D.; Rosalen, P.L. Alternative animal and non-animal models for drug discovery and development: Bonus or burden? Pharm. Res. 2017, 34, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, O.; Theopold, U.; Beckage, N.E. Insect and vertebrate immunity: Key similarities versus differences. In Insect Immunology; Beckage, N.E., Ed.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 1–23. [Google Scholar] [CrossRef]
- Müller, U.; Vogel, P.; Alber, G.; Schaub, G. The innate immune system of mammals and insects. Contrib. Microbiol. 2008, 15, 21–44. [Google Scholar]
- Kavanagh, K.; Reeves, E.P. Insect and mammalian innate immune responses are much alike. Microbe 2007, 2, 596–599. [Google Scholar] [CrossRef]
- Jander, G.; Rahme, L.G.; Ausubel, F.M. Positive correlation between virulence of Pseudomonas aeruginosa mutants in mice and insects. J. Bacteriol. 2000, 182, 3843–3845. [Google Scholar] [CrossRef]
- Jønsson, R.; Struve, C.; Jenssen, H.; Krogfelt, K.A. The wax moth Galleria mellonella as a novel model system to study Enteroaggregative Escherichia coli pathogenesis. Virulence 2017, 8, 1894–1899. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.J.Y.; Loh, J.M.S.; Proft, T. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 2016, 7, 214–229. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Feng, Y.; Zong, Z. Two new lytic bacteriophages of the Myoviridae family against carbapenem-resistant Acinetobacter baumannii. Front. Microbiol. 2018, 9, 850. [Google Scholar] [CrossRef]
- Hornsey, M.; Wareham, D.W. Effects of in vivo emergent tigecycline resistance on the pathogenic potential of Acinetobacter baumannii. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Peleg, A.Y.; Jara, S.; Monga, D.; Eliopoulos, G.M.; Moellering, R.C., Jr.; Mylonakis, E. Galleria mellonella as a model system to study Acinetobacter baumannii pathogenesis and therapeutics. Antimicrob. Agents Chemother. 2009, 53, 2605–2609. [Google Scholar] [CrossRef]
- Wand, M.E.; Bock, L.J.; Turton, J.F.; Nugent, P.G.; Mark Sutton, J. Acinetobacter baumannii virulence is enhanced in Galleria mellonella following biofilm adaptation. J. Med. Microbiol. 2012, 61, 470–477. [Google Scholar] [CrossRef]
- Hornsey, M.; Phee, L.; Longshaw, C.; Wareham, D.W. In vivo efficacy of telavancin/colistin combination therapy in a Galleria mellonella model of Acinetobacter baumannii infection. Int. J. Antimicrob. Agents 2013, 41, 285–287. [Google Scholar] [CrossRef]
- Mannala, G.K.; Koettnitz, J.; Mohamed, W.; Sommer, U.; Lips, K.S.; Spröer, C.; Bunk, B.; Overmann, J.; Hain, T.; Heiss, C.; et al. Whole-genome comparison of high and low virulent Staphylococcus aureus isolates inducing implant-associated bone infections. Int. J. Med. Microbiol. 2018, 308, 505–513. [Google Scholar] [CrossRef]
- Ba, X.; Harrison, E.M.; Lovering, A.L.; Gleadall, N.; Zadoks, R.; Parkhill, J.; Peacock, S.J.; Holden, M.T.G.; Paterson, G.K.; Holmes, M.A. Old drugs to treat resistant bugs: Methicillin-resistant Staphylococcus aureus isolates with mecC are susceptible to a combination of penicillin and clavulanic acid. Antimicrob. Agents Chemother. 2015, 59, 7396–7404. [Google Scholar] [CrossRef] [PubMed]
- Bazaid, A.S.; Forbes, S.; Humphreys, G.J.; Ledder, R.G.; O’Cualain, R.; McBain, A.J. Fatty acid supplementation reverses the small colony variant phenotype in triclosan-adapted Staphylococcus aureus: Genetic, proteomic and phenotypic analyses. Sci. Rep. 2018, 8, 3876. [Google Scholar] [CrossRef]
- Dong, C.L.; Li, L.X.; Cui, Z.H.; Chen, S.W.; Xiong, Y.Q.; Lu, J.Q.; Liao, X.P.; Gao, Y.; Sun, J.; Liu, Y.H. Synergistic effect of pleuromutilins with other antimicrobial agents against Staphylococcus aureus in vitro and in an experimental Galleria mellonella model. Front. Pharmacol. 2017, 8, 553. [Google Scholar] [CrossRef] [PubMed]
- Ebner, P.; Rinker, J.; Nguyen, M.T.; Popella, P.; Nega, M.; Luqman, A.; Schittek, B.; Di Marco, M.; Stevanovic, S.; Götza, F. Excreted cytoplasmic proteins contribute to pathogenicity in Staphylococcus aureus. Infect. Immun. 2016, 84, 1672–1681. [Google Scholar] [CrossRef] [PubMed]
- Ferro, T.A.F.; Araújo, J.M.M.; Pinto, B.L.S.; dos Santos, J.S.; Souza, E.B.; da Silva, B.L.R.; Colares, V.L.; Novais, T.M.G.; Filho, C.M.B.; Struve, C.; et al. Cinnamaldehyde inhibits Staphylococcus aureus virulence factors and protects against infection in a Galleria mellonella model. Front. Microbiol. 2016, 7, 2052. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.C.; DiDone, L.; Jobson, J.; Sofia, M.K.; Krysan, D.; Dunman, P.M. Adenylate kinase release as a high-throughput-screening-compatible reporter of bacterial lysis for identification of antibacterial agents. Antimicrob. Agents Chemother. 2013, 57, 26–36. [Google Scholar] [CrossRef]
- Jorjão, A.L.; Oliveira, L.D.; Scorzoni, L.; Figueiredo-Godoi, L.M.A.; Prata, M.C.A.; Jorge, A.O.C.; Junqueira, J.C. From moths to caterpillars: Ideal conditions for Galleria mellonella rearing for in vivo microbiological studies. Virulence 2018, 9, 383–389. [Google Scholar] [CrossRef]
- Saporito, P.; Mouritzen, M.V.; Lobner-Olesen, A.; Jenssen, H. LL-37 fragments have antimicrobial activity against Staphylococcus epidermidis biofilms and wound healing potential in HaCaT cell line. J. Pept. Sci. 2018, 24, e3080. [Google Scholar] [CrossRef]
- Gaglione, R.; Dell’Olmo, E.; Bosso, A.; Chino, M.; Pane, K.; Ascione, F.; Itri, F.; Caserta, S.; Amoresano, A.; Lombardi, A.; et al. Novel human bioactive peptides identified in Apolipoprotein B: Evaluation of their therapeutic potential. Biochem. Pharmacol. 2017, 130, 34–50. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Aspect | Larva Size | Larva Origin | Larva Storage | Injection Site | Infection Dose CFU/larva | Antimicrobial Dose, Expressed as | Ref. |
|---|---|---|---|---|---|---|---|
| A. baumannii | 250–350 mg | P-China | 4 °C, 7 days | LP | 1 × 105 | mg/larva kg | [8] |
| Not given | P-UK | 15 °C | Left proleg | 1 × 104 | Not relevant | [9] | |
| 250–350 mg | P-USA | 7 days | LP | 5 × 105 | mg/larva kg | [10] | |
| Not given | P-UK | RT, 14 days | FP | 1 × 105, 1 × 104 | Not relevant | [11] | |
| 250 ± 25 mg | P-UK | 15 °C | Not given | 1 × 105 | mg/larva kg | [12] | |
| S. aureus | 150–200 mg | Reared | 30 °C | Not given | 1 × 106 | Not relevant | [13] |
| Not given | P-UK | 4 °C | Between segments | ~1.3 × 106 | mg/larva kg | [14] | |
| 15–25 mm long | P-UK | 7 days | LP | 0.8–2.6 × 106 | Not relevant | [15] | |
| ~250 mg | P-China | Not given | LP | ~1 × 106 | mg/larva kg | [16] | |
| 300–700 mg | P-Netherlands | Not given | LP | 1 × 106 | Not relevant | [17] | |
| ~200 mg | Not given | Not given | LP | 1 × 103 | µg/100 mg larva | [18] | |
| 200–300 mg | P-USA | 4 °C, 14 days | LP | 5 × 106 | mg/larva kg | [19] | |
| Not given | Reared | 28 °C | Not given | Not given | Not relevant | [20] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrea, A.; Krogfelt, K.A.; Jenssen, H. Methods and Challenges of Using the Greater Wax Moth (Galleria mellonella) as a Model Organism in Antimicrobial Compound Discovery. Microorganisms 2019, 7, 85. https://doi.org/10.3390/microorganisms7030085
Andrea A, Krogfelt KA, Jenssen H. Methods and Challenges of Using the Greater Wax Moth (Galleria mellonella) as a Model Organism in Antimicrobial Compound Discovery. Microorganisms. 2019; 7(3):85. https://doi.org/10.3390/microorganisms7030085
Chicago/Turabian StyleAndrea, Athina, Karen Angeliki Krogfelt, and Håvard Jenssen. 2019. "Methods and Challenges of Using the Greater Wax Moth (Galleria mellonella) as a Model Organism in Antimicrobial Compound Discovery" Microorganisms 7, no. 3: 85. https://doi.org/10.3390/microorganisms7030085
APA StyleAndrea, A., Krogfelt, K. A., & Jenssen, H. (2019). Methods and Challenges of Using the Greater Wax Moth (Galleria mellonella) as a Model Organism in Antimicrobial Compound Discovery. Microorganisms, 7(3), 85. https://doi.org/10.3390/microorganisms7030085