The Non-Simultaneous Enhancement of Phosphorus Acquisition and Mobilization Respond to Enhanced Arbuscular Mycorrhization on Maize (Zea mays L.)


Abstract
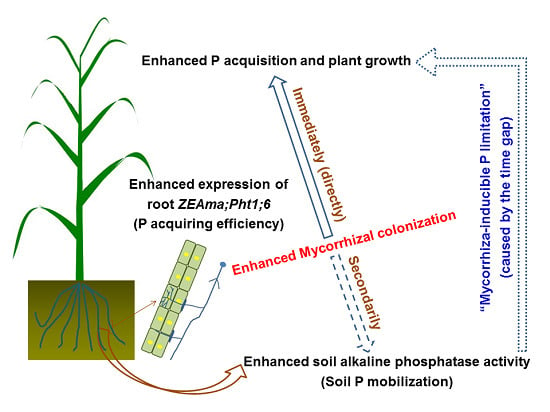
1. Introduction
2. Materials and Methods
2.1. Soil and Mycorrhizal Inoculum Preparation
2.2. Pot Experiment
2.3. Mycorrhizal Colonization, Plant Biomass, and P Concentration Analysis
2.4. Root ZEAma;Pht1;6 Expression Analysis
2.5. Soil pH, ALP Activity, and Available P Concentration Analysis
2.6. Statistical Analysis
3. Results
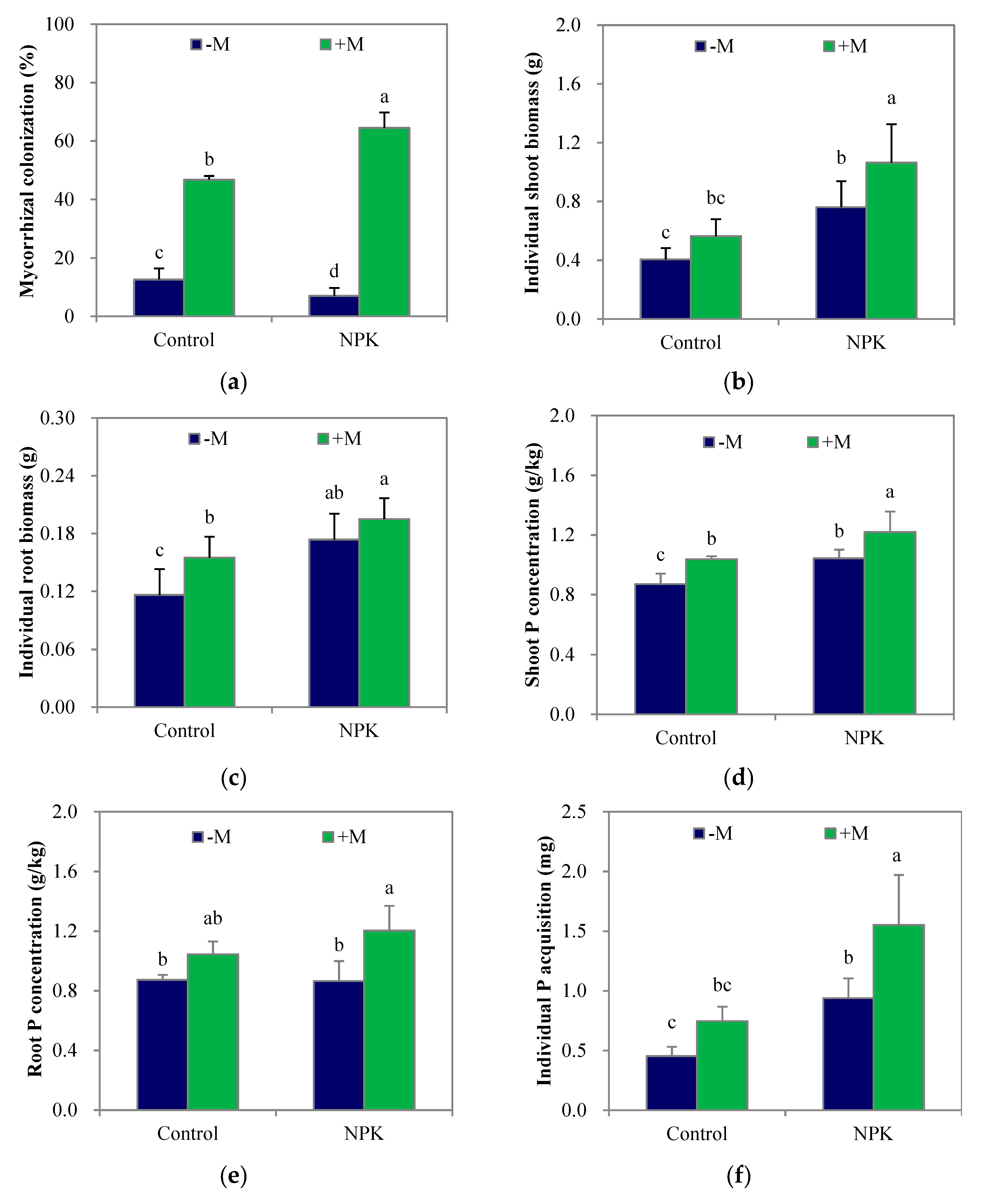
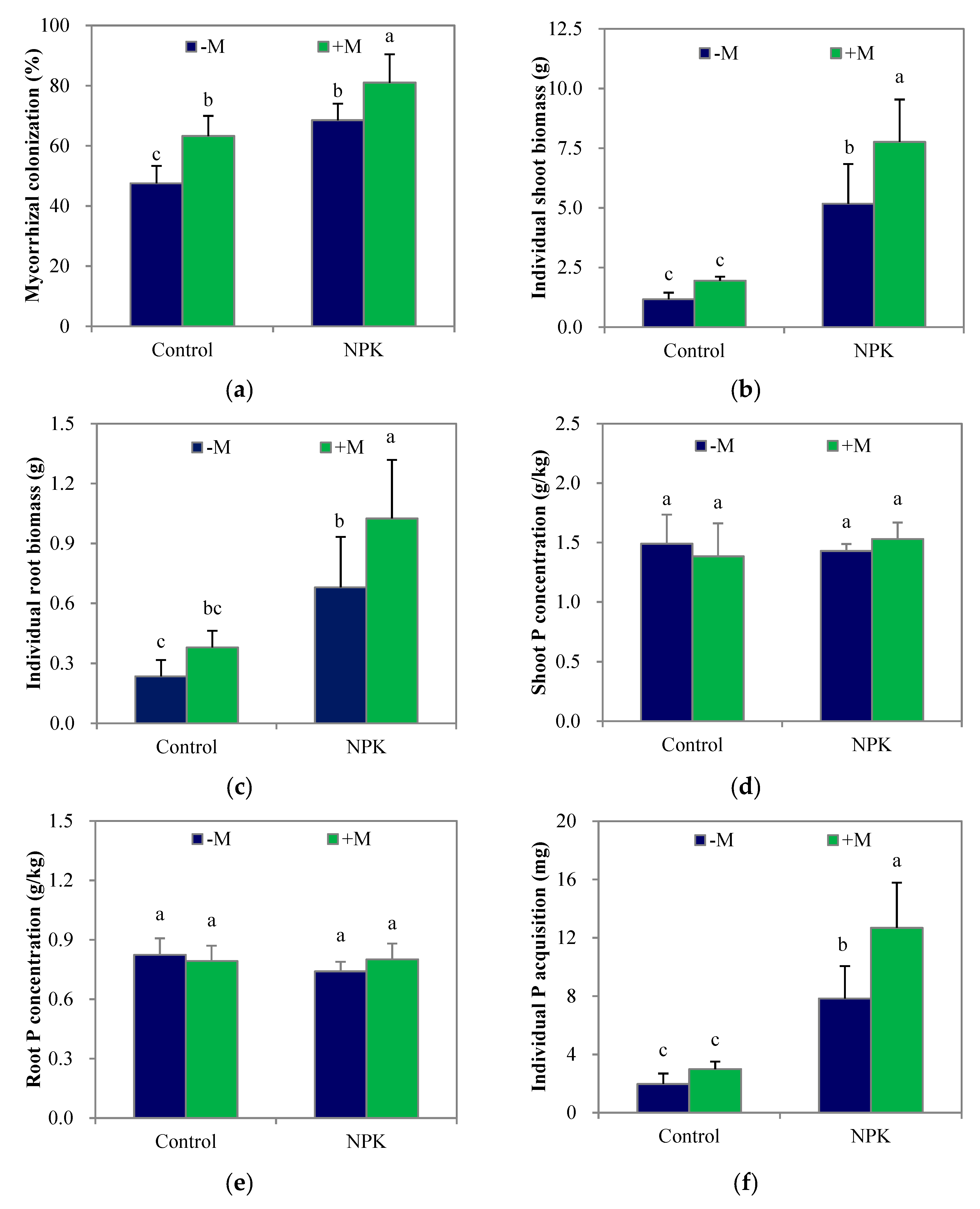
3.1. Mycorrhizal Colonization, Plant Biomass and P Acquisition
3.2. Mycorrhiza-Inducible Pi Transporter Expression and Soil P Mobilization
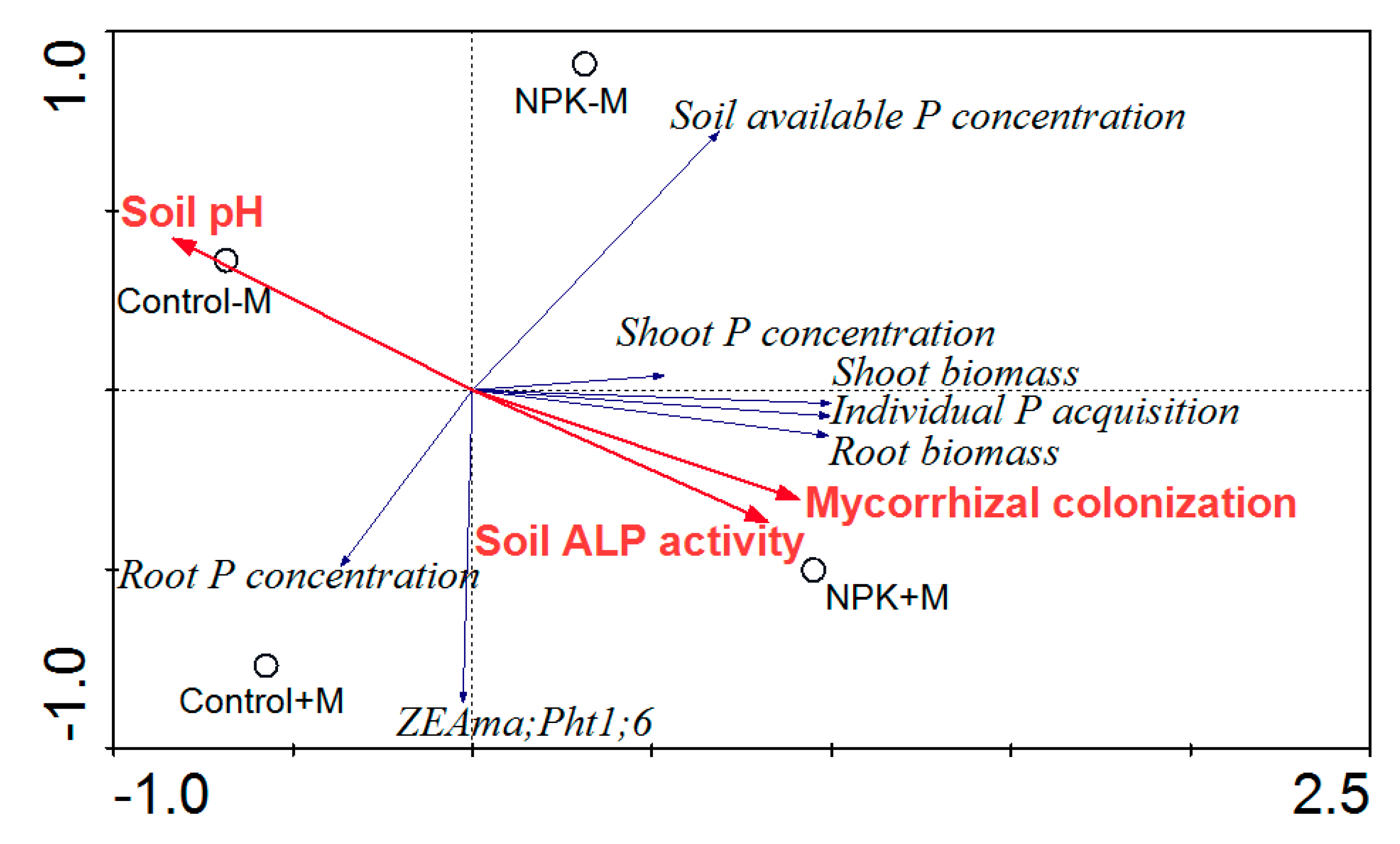
3.3. RDA Results
4. Discussion
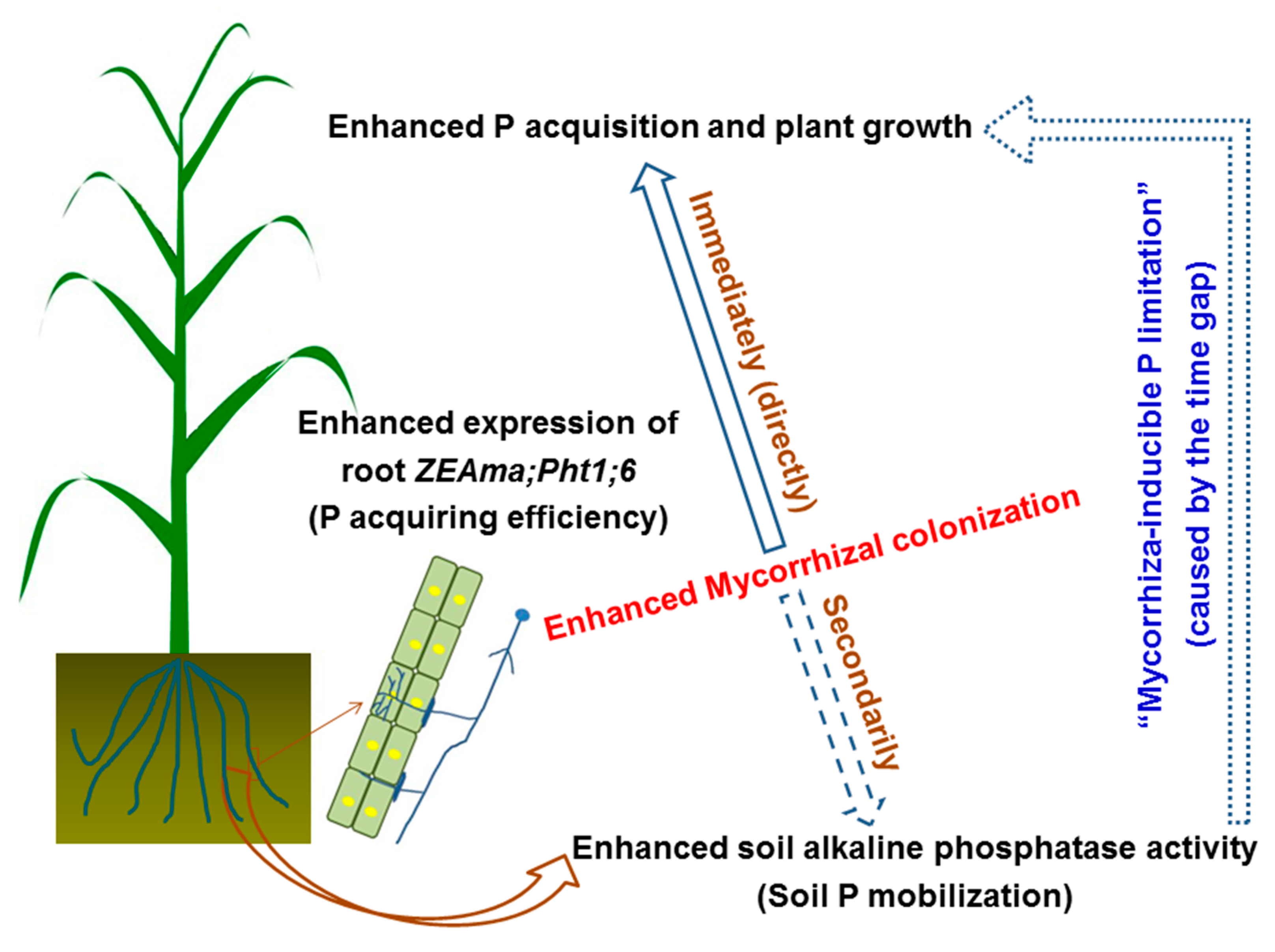
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khan, M.S.; Zaidi, A.; Wani, P.A. Role of phosphate-solubilizing microorganisms in sustainable agriculture—A review. Agron. Sustain. Dev. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Wang, F.; Sun, Y.; Shi, Z. Arbuscular mycorrhiza enhances biomass production and salt tolerance of sweet sorghum. Microorganisms 2019, 7, 289. [Google Scholar] [CrossRef] [PubMed]
- Baslam, M.; Garmendia, I.; Goicoechea, N. Arbuscular mycorrhizal fungi (AMF) improved growth and nutritional quality of greenhouse-grown lettuce. J. Agric. Food Chem. 2011, 59, 5504–5515. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.Y.; Lin, X.G.; Yin, R.; Wu, L.H. Effects of arbuscular mycorrhizal inoculation on the growth of Elsholtzia splendens and Zea mays and the activities of phosphatase and urease in a multi-metal- contaminated soil under unsterilized conditions. Appl. Soil Ecol. 2006, 31, 110–119. [Google Scholar] [CrossRef]
- Brundrett, M.C. Mycorrhizal associations and other means of nutrition of vascular plants: Understanding the global diversity of hostplants by resolving conflicting information and developing reliable means of diagnosis. Plant Soil 2009, 320, 37–77. [Google Scholar] [CrossRef]
- Feng, K.; Lu, H.M.; Sheng, H.J.; Wang, X.L.; Mao, J. Effect of organic ligands on biological availability of inorganic phosphorus in soils. Pedosphere 2004, 14, 85–92. [Google Scholar]
- Doolette, A.; Armstrong, R.; Tang, C.; Guppy, C.; Mason, S.; McNeill, A. Phosphorus uptake benefit for wheat following legume break crops in semi-arid Australian farming systems. Nutr. Cycl. Agroecosyst. 2019, 113, 247–266. [Google Scholar] [CrossRef]
- Zheng, C.; Zhang, J.; Li, X. Phosphorus supply level affects the regulation of phosphorus uptake by different arbuscular mycorrhizal fungal species in a highly P-efficient backcross maize line. Crop Past Sci. 2013, 64, 881–891. [Google Scholar] [CrossRef]
- Willmann, M.; Gerlach, N.; Buer, B.; Polatajko, A.; Nagy, R.; Koebke, E.; Jansa, J.; Flisch, R.; Bucher, M. Mycorrhizal phosphate uptake pathway in maize: Vital for growth and cob development on nutrient poor agricultural and greenhouse soils. Front. Plant Sci. 2013, 4, 533. [Google Scholar] [CrossRef]
- Hu, J.; Lin, X.; Wang, J.; Chu, H.; Yin, R.; Zhang, J. Population size and specific potential of P-mineralizing and -solubilizing bacteria under long-term P-deficiency fertilization in a sandy loam soil. Pedobiologia 2009, 53, 49–58. [Google Scholar] [CrossRef]
- Jone, E.J. The effect of long-term fertilization with organic or inorganic fertilizers on mycorrhiza-mediated phosphorus uptake in subterranean clover. Biol. Fertil. Soils 2000, 32, 435–440. [Google Scholar] [CrossRef]
- Chu, H.; Lin, X.; Fujii, T.; Morimoto, S.; Yagi, K.; Hu, J.; Zhang, J. Soil microbial biomass, dehydrogenase activity, bacterial community structure in response to long-term fertilizer management. Soil Biol. Biochem. 2007, 39, 2971–2976. [Google Scholar] [CrossRef]
- Chowdhury, R.B.; Milne, N.; Chakraborty, P. Overuse of phosphorus resources. In Encyclopedia of Food Security and Sustainability, 1st ed.; Ferranti, P., Berry, E.M., Anderson, I.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 249–254. [Google Scholar]
- Hu, J.; Lin, X.; Wang, J.; Dai, J.; Cui, X.; Chen, R.; Zhang, J. Arbuscular mycorrhizal fungus enhances crop yield and P-uptake of maize (Zea mays L.): A field case study on a sandy loam soil as affected by long-term P-deficiency fertilization. Soil Biol. Biochem. 2009, 41, 2460–2465. [Google Scholar] [CrossRef]
- Bucher, M. Functional biology of plant phosphate uptake at root and mycorrhiza interfaces. New Phytol. 2007, 173, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Jakobsen, I.; Grønlund, M.; Smith, F.A. Roles of arbuscular mycorrhizas in plant phosphorus nutrition: Interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol. 2011, 156, 1050–1057. [Google Scholar] [CrossRef]
- Smith, S.E.; Smith, F.A. Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef]
- Smith, S.E.; Smith, F.A. Fresh perspectives on the roles of arbuscular mycorrhizal fungi in plant nutrition and growth. Mycologia 2012, 104, 1–13. [Google Scholar] [CrossRef]
- Benedetto, A.; Magurno, F.; Bonfante, P.; Lanfranco, L. Expression profiles of a phosphate transporter gene (GmosPT) from the endomycorrhizal fungus Glomus mosseae. Mycorrhiza 2005, 15, 620–627. [Google Scholar] [CrossRef]
- Javot, H.; Pumplin, N.; Harrison, M.J. Phosphate in the arbuscular mycorrhizal symbiosis: Transport properties and regulatory roles. Plant Cell Environ. 2007, 30, 310–322. [Google Scholar] [CrossRef]
- Grace, E.J.; Cotsaftis, O.; Tester, M.; Smith, F.A.; Smith, S.E. Arbuscular mycorrhizal inhibition of growth in barley cannot be attributed to extent of colonization, fungal phosphorus uptake or effects on expression of plant phosphate transporter genes. New Phytol. 2009, 181, 938–949. [Google Scholar] [CrossRef]
- Liu, F.; Xu, Y.; Han, G.; Wang, W.; Li, X.; Cheng, B. Identification and functional characterization of a maize phosphate transporter induced by mycorrhiza formation. Plant Cell Physiol. 2018, 59, 1683–1694. [Google Scholar] [CrossRef] [PubMed]
- Plassard, C.; Becquer, A.; Garcia, K. Phosphorus transport in mycorrhiza: How far are we? Trends Plant Sci. 2019, 24, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Glassop, D.; Smith, S.E.; Smith, F.W. Cereal phosphate transporters associated with the mycorrhizal pathway of phosphate uptake into roots. Planta 2005, 222, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Nagy, R.; Vasconcelos, M.J.V.; Zhao, S.; McElver, J.; Bruce, W.; Amrhein, N.; Raghothama, K.G.; Bucher, M. Differential regulation of five Pht1 phosphate transporters from maize (Zea mays L.). Plant Biol. 2006, 8, 186–197. [Google Scholar] [CrossRef]
- Chen, B.; Li, X.; Christie, P. Two arbuscular mycorrhizal fungi colonizing maize under different phosphorus regimes in a compartment cultivation system. Pedosphere 2002, 12, 121–130. [Google Scholar]
- Cao, J.; Huang, Y.; Wang, C. Rhizosphere interactions between earthworms (Eisenia fetida) and arbuscular mycorrhizal fungus (Funneliformis mosseae) promote utilization efficiency of phytate phosphorus in maize. Appl. Soil Ecol. 2015, 94, 30–39. [Google Scholar] [CrossRef]
- Watts-Williams, S.J.; Smith, F.A.; Jakobsen, I. Soil phosphorus availability is a driver of the responses of maize (Zea mays) to elevated CO2 concentration and arbuscular mycorrhizal colonisation. Symbiosis 2019, 77, 73–82. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular- arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An evaluation of techniques for measuring vesicular–arbuscular mycorrhizal infection in roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Zhu, Y.; Smith, F.A.; Smith, S.E. Phosphorus efficiencies and responses of barley (Hordeum vulgare L.) to arbuscular mycorrhizal fungi grown in highly calcareous soil. Mycorrhiza 2003, 13, 93–100. [Google Scholar] [CrossRef]
- Hanson, W.C. The photometric determination of phosphorus in fertilisers using the phosphovanado–molybdate complex. J. Sci. Food Agric. 1950, 1, 172–173. [Google Scholar] [CrossRef]
- Ding, D.; Zhang, L.F.; Wang, H.; Liu, Z.J.; Zhang, Z.X.; Zheng, Y.L. Differential expression of miRNAs in response to salt stress in maize roots. Ann. Bot. 2009, 103, 29–38. [Google Scholar] [CrossRef]
- Tian, H.; Drijber, R.A.; Li, X.; Miller, D.N.; Wienhold, B.J. Arbuscular mycorrhizal fungi differ in their ability to regulate the expression of phosphate transporters in maize (Zea mays L.). Mycorrhiza 2013, 23, 507–514. [Google Scholar] [CrossRef]
- Tabatabai, M.A. Soil enzymes. In Methods of Soil Analyses, Part 2, Chemical and Microbiological Properties, 2nd ed.; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 903–947. [Google Scholar]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; United States Department of Agriculture: Washington, DC, USA, 1954; p. 19.
- Igiehon, N.O.; Babalola, O.O. Biofertilizers and sustainable agriculture: Exploring arbuscular mycorrhizal fungi. Appl. Microbiol. Biotechnol. 2017, 101, 4871–4881. [Google Scholar] [CrossRef]
- Deng, Y.; Feng, G.; Chen, X.P.; Zou, C.Q. Arbuscular mycorrhizal fungal colonization is considerable at optimal Olsen-P levels for maximized yields in an intensive wheat-maize cropping system. Field Crop Res. 2017, 209, 1–9. [Google Scholar] [CrossRef]
- Duffková, R.; Fučík, P.; Jurkovská, L.; Janoušková, M. Experimental evaluation of the potential of arbuscular mycorrhiza to modify nutrient leaching in three arable soils located on one slope. Appl. Soil Ecol. 2019, 143, 116–125. [Google Scholar] [CrossRef]
- Li, X.L.; George, E.; Marschner, H. Extension of the phosphorus depletion zone in VA-mycorrhizal white clover in a calcareous soil. Plant Soil 1991, 136, 41–48. [Google Scholar] [CrossRef]
- Hu, J.; Lin, X.; Wang, J.; Cui, X.; Dai, J.; Chu, H.; Zhang, J. Arbuscular mycorrhizal fungus enhances P-acquisition of wheat (Triticum aestivum L.) in a sandy loam soil with long-term inorganic fertilization regime. Appl. Microbiol. Biotechnol. 2010, 88, 781–787. [Google Scholar] [CrossRef]
- Shi, S.; Chen, K.; Gao, Y.; Liu, B.; Yang, X.; Huang, X.; Liu, G.; Zhu, L.; He, X. Arbuscular mycorrhizal fungus species dependency governs better plant physiological characteristics and leaf quality of mulberry (Morus alba L.) seedlings. Front. Microbiol. 2016, 7, 1030. [Google Scholar] [CrossRef]
- Chen, S.; Zhao, H.; Zou, C.; Li, Y.; Chen, Y.; Wang, Z.; Jiang, Y.; Liu, A.; Zhao, P.; Wang, M.; et al. Combined inoculation with multiple arbuscular mycorrhizal fungi improves growth, nutrient uptake and photosynthesis in cucumber seedlings. Front. Microbiol. 2017, 8, 2516. [Google Scholar] [CrossRef]
- Miransari, M. Interactions between arbuscular mycorrhizal fungi and soil bacteria. Appl. Microbiol. Biotechnol. 2011, 89, 917–930. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mycorrhizal Colonization | Soil pH | Soil ALP Activity | Soil Available P Concentration | Root ZEAma;Pht1;6 | Shoot Biomass | Root Biomass | |
|---|---|---|---|---|---|---|---|
| Soil pH | −0.988 ** | ||||||
| Soil ALP activity | 0.983 ** | −0.996 ** | |||||
| Soil available P concentration | 0.431 | −0.302 | 0.340 | ||||
| Root ZEAma;Pht1;6 | 0.381 | −0.521 | 0.513 | −0.600 | |||
| Shoot biomass | 0.928 * | −0.858 | 0.847 | 0.661 | 0.021 | ||
| Root biomass | 0.952 ** | −0.895 | 0.880 | 0.593 | 0.106 | 0.996 ** | |
| Individual P acquisition | 0.922 * | −0.853 | 0.836 | 0.630 | 0.026 | 0.998 ** | 0.996 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Cui, X.; Wang, J.; Lin, X. The Non-Simultaneous Enhancement of Phosphorus Acquisition and Mobilization Respond to Enhanced Arbuscular Mycorrhization on Maize (Zea mays L.). Microorganisms 2019, 7, 651. https://doi.org/10.3390/microorganisms7120651
Hu J, Cui X, Wang J, Lin X. The Non-Simultaneous Enhancement of Phosphorus Acquisition and Mobilization Respond to Enhanced Arbuscular Mycorrhization on Maize (Zea mays L.). Microorganisms. 2019; 7(12):651. https://doi.org/10.3390/microorganisms7120651
Chicago/Turabian StyleHu, Junli, Xiangchao Cui, Junhua Wang, and Xiangui Lin. 2019. "The Non-Simultaneous Enhancement of Phosphorus Acquisition and Mobilization Respond to Enhanced Arbuscular Mycorrhization on Maize (Zea mays L.)" Microorganisms 7, no. 12: 651. https://doi.org/10.3390/microorganisms7120651
APA StyleHu, J., Cui, X., Wang, J., & Lin, X. (2019). The Non-Simultaneous Enhancement of Phosphorus Acquisition and Mobilization Respond to Enhanced Arbuscular Mycorrhization on Maize (Zea mays L.). Microorganisms, 7(12), 651. https://doi.org/10.3390/microorganisms7120651