The Effects of Colicin Production Rates on Allelopathic Interactions in Escherichia coli Populations

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids and Growth Conditions
2.2. General DNA Techniques
2.3. Bacteriocin Production Assay
2.4. Competition Assays
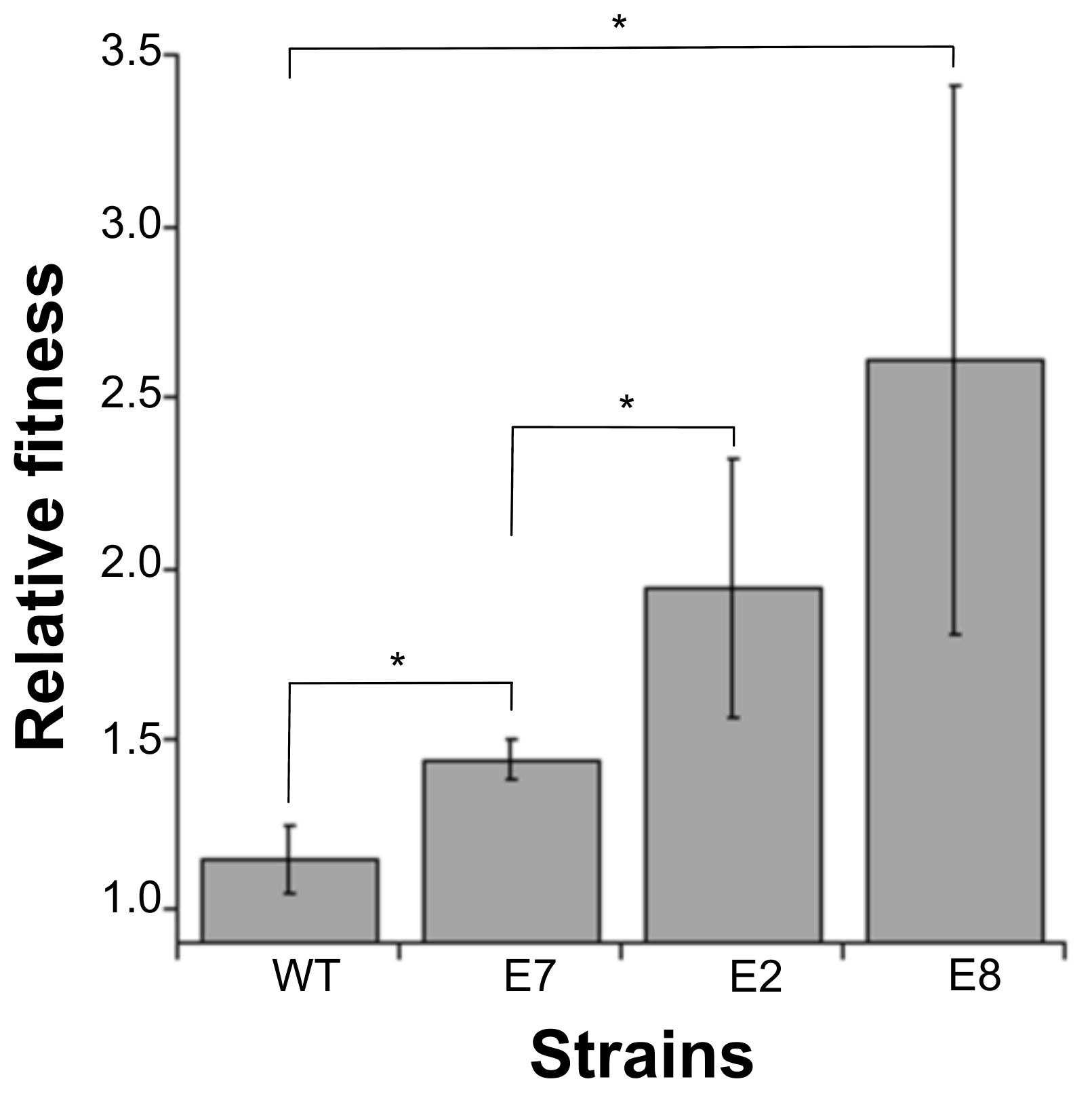
2.5. Relative Fitness of the Colicinogenic Strains
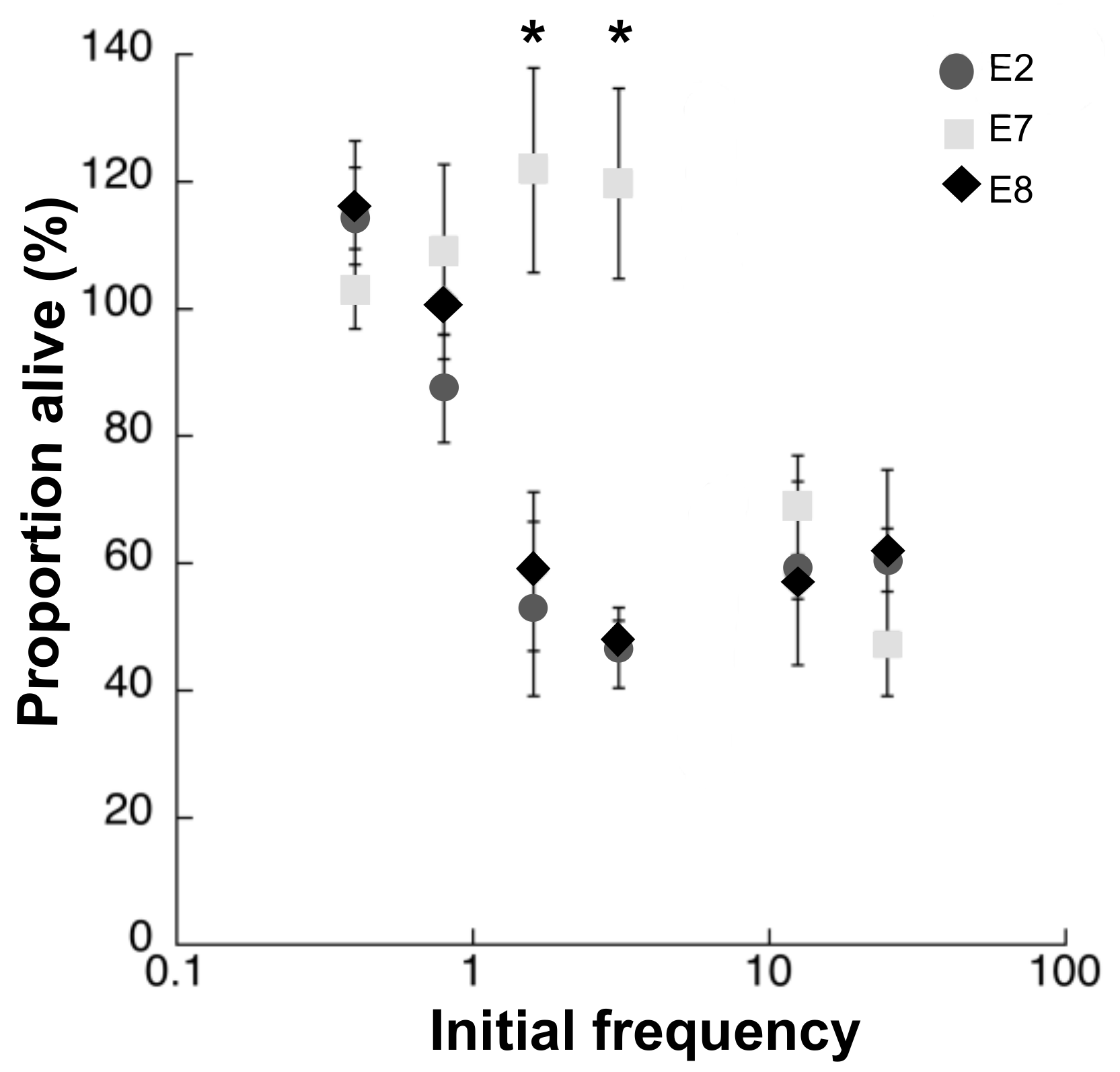
3. Results
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Granato, E.T.; Meiller-Legrand, T.A.; Foster, K.R. The Evolution and Ecology of Bacterial Warfare. Curr. Biol. 2019, 29, R521–R537. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Cascales, E. Antibacterial Weapons: Targeted Destruction in the Microbiota. Trends Microbiol. 2018, 26, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Gardner, A.; West, S.A.; Buckling, A. Bacteriocins, spite and virulence. Proc. R. Soc. Lond. B Biol. 2004, 271, 1529–1535. [Google Scholar] [CrossRef] [PubMed]
- Inglis, R.F.; Gardner, A.; Cornelis, P.; Buckling, A. Spite and virulence in the bacterium Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2009, 106, 5703–5707. [Google Scholar] [CrossRef] [PubMed]
- Nadell, C.D.; Drescher, K.; Foster, K.R. Spatial structure, cooperation and competition in biofilms. Nat. Rev. Microbiol. 2016, 14, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Greig, D.; Travisano, M. Density-dependent effects on allelopathic interactions in yeast. Evolution 2008, 62, 521–527. [Google Scholar] [CrossRef]
- Wloch-Salamon, D.M.; Gerla, D.; Hoekstra, R.F.; de Visser, J. Effect of dispersal and nutrient availability on the competitive ability of toxin-producing yeast. Proc. R. Soc. Lond. B Biol. 2008, 275, 535–541. [Google Scholar] [CrossRef]
- Chao, L.; Levin, B.R. Structured habitats and the evolution of anticompetitor toxins in bacteria. Proc. Natl. Acad. Sci. USA 1981, 78, 6324–6328. [Google Scholar] [CrossRef]
- Iwasa, Y.; Nakamaru, M.; Levin, S.A. Allelopathy of bacteria in a lattice population: Competition between colicin-sensitive and colicin-producing strains. Evol. Ecol. 1988, 12, 785–802. [Google Scholar] [CrossRef]
- Abrudan, M.I.; Smakman, F.; Grimbergen, A.J.; Westhoff, S.; Miller, E.L.; van Wezel, G.P.; Rozen, D.E. Socially mediated induction and suppression of antibiosis during bacterial coexistence. Proc. Natl. Acad. Sci. USA 2015, 112, 11054–11059. [Google Scholar] [CrossRef]
- Libberton, B.; Horsburgh, M.J.; Brockhurst, M.A. The effects of spatial structure, frequency dependence and resistance evolution on the dynamics of toxin-mediated microbial invasions. Evol. Appl. 2015, 8, 738–750. [Google Scholar] [CrossRef] [PubMed]
- Cornforth, D.M.; Foster, K.R. Competition sensing: The social side of bacterial stress responses. Nat. Rev. Microbiol. 2013, 11, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Biernaskie, J.M.; Gardner, A.; West, S.A. Multicoloured greenbeards, bacteriocin diversity and the rock-paper-scissors game. J. Evol. Biol. 2013, 26, 2081–2094. [Google Scholar] [CrossRef] [PubMed]
- Majeed, H.; Lampert, A.; Ghazaryan, L.; Gillor, O. The Weak Shall Inherit: Bacteriocin-Mediated Interactions in Bacterial Populations. PLoS ONE 2013, 8, e63837. [Google Scholar] [CrossRef] [PubMed]
- Durrett, R.; Levin, S. Allelopathy in spatially distributed populations. J. Theor. Biol. 1997, 185, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Nakamaru, M.; Iwasa, Y. Competition by allelopathy proceeds in traveling waves: Colicin-immune strain aids colicin-sensitive strain. Theor. Popul. Biol. 2000, 57, 131–144. [Google Scholar] [CrossRef] [PubMed]
- Frean, M.; Abraham, E.R. Rock–scissors–paper and the survival of the weakest. Proc. R. Soc. Lond. B Biol. Sci. 2001, 268, 1323–1327. [Google Scholar] [CrossRef]
- Berr, M.; Reichenbach, T.; Schottenloher, M.; Frey, E. Zero-One Survival Behavior of Cyclically Competing Species. Phys. Rev. Lett. 2009, 102, 048102. [Google Scholar] [CrossRef]
- Pintar, J.; Starmer, W.T. The costs and benefits of killer toxin production by the yeast Pichia kluyveri. Van Leeuwenhoek J. Microbiol. 2003, 83, 89–97. [Google Scholar] [CrossRef]
- Hibbing, M.; Fuqua, C.; Parsek, M.; Peterson, S. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol. 2010, 8, 15–25. [Google Scholar] [CrossRef]
- Cascales, E.; Buchanan, S.K.; Duche, D.; Kleanthous, C.; Lloubes, R.; Postle, K.; Riley, M.; Slatin, S.; Cavard, D. Colicin Biology. Microbiol. Mol. Biol. Rev. 2007, 71, 158–229. [Google Scholar] [CrossRef] [PubMed]
- Ghazaryan, L.; Soares, M.I.M.; Gillor, O. Auto-regulation of DNA degrading bacteriocins: Molecular and ecological aspects. Antonie Van Leeuwenhoek 2014, 105, 823–834. [Google Scholar] [CrossRef] [PubMed]
- Pugsley, A. Escherichia coli K12 strains for use in the identification and characterization of Colicins. J. Gen. Microbiol. 1985, 131, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Zaslaver, A.; Bren, A.; Ronen, M.; Itzkovitz, S.; Kikoin, I.; Shavit, S.; Liebermeister, W.; Surette, M.G.; Alon, U. A comprehensive library of fluorescent transcriptional reporters for Escherichia coli. Nat. Methods 2006, 3, 623–628. [Google Scholar] [CrossRef]
- Gordon, D.M.; Riley, M.A. A theoretical and empirical investigation of the invasion dynamics of colicinogeny. Microbiology 1999, 145, 655–661. [Google Scholar] [CrossRef]
- Gordon, D.M.; Riley, M.A.; Pinou, T. Temporal changes in the frequency of colicinogeny in Escherichia coli from house mice. Microbiology 1998, 144, 2233–2240. [Google Scholar] [CrossRef]
- Cormack, B.P.; Valdivia, R.H.; Falkow, S. FACS-optimized mutants of the green fluorescent protein (GFP). Gene 1996, 173, 33–38. [Google Scholar] [CrossRef]
- Dunny, G.M.; Brickman, T.J.; Dworkin, M. Multicellular behavior in bacteria: Communication, cooperation, competition and cheating. Bioessays 2008, 30, 296–298. [Google Scholar] [CrossRef]
- Leroi, A.M.; Bennett, A.F.; Lenski, R.E. Temperature acclimation and competitive fitness: An experimental test of the beneficial acclimation assumption. Proc. Natl. Acad. Sci. USA 1994, 91, 1917–1921. [Google Scholar] [CrossRef]
- Hardy, K.G.; Harwood, C.R.; Meynell, G.G. Expression of colicin factor E2-P9. MGG Mol. Gen. Genet. 1974, 131, 313–331. [Google Scholar] [CrossRef]
- Inglis, R.F.; Roberts, P.G.; Gardner, A.; Buckling, A. Spite and the scale of competition in Pseudomonas aeruginosa. Am. Nat. 2011, 178, 276–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, S.A. Spatial polymorphism of bacteriocins and other allelopathic traits. Evol. Ecol. 1994, 8, 369–386. [Google Scholar] [CrossRef]
- Levin, B.R.; Antonovics, J.; Sharma, H. Frequency-Dependent Selection in Bacterial Populations: Discussion. Philos. Trans. R. Soc. B Biol. Sci. 1988, 319, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Resch, A.; Rosenstein, R.; Nerz, C.; Gotz, F. Differential gene expression profiling of staphylococcus aureus cultivated under biofilm and planktonic conditions. Appl. Environ. Microbiol. 2005, 71, 2663–2676. [Google Scholar] [CrossRef] [Green Version]
- Beloin, C.; Roux, A.; Ghigo, J.M. Escherichia coli biofilms. Curr. Top. Microbiol. Immunol. 2008, 322, 249–289. [Google Scholar]
- Bayramoglu, B.; Toubiana, D.; Gillor, O. Genome-wide transcription profiling of aerobic and anaerobic Escherichia coli biofilm and planktonic cultures. FEMS Microbiol. Lett. 2017, 364, fnx006. [Google Scholar] [CrossRef] [Green Version]
- Micenková, L.; Bosák, J.; Kucera, J.; Hrala, M.; Dolejšová, T.; Šedo, O.; Linke, D.; Fišer, R.; Šmajs, D. Colicin Z, a structurally and functionally novel colicin type that selectively kills enteroinvasive Escherichia coli and Shigella strains. Sci. Rep. 2019, 9, 11127. [Google Scholar] [CrossRef]
- Bosák, J.; Micenková, L.; Hrala, M.; Pomorská, K.; Kunova Bosakova, M.; Krejci, P.; Göpfert, E.; Faldyna, M.; Šmajs, D. Colicin FY inhibits pathogenic Yersinia enterocolitica in mice. Sci. Rep. 2018, 8, 12242. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains/ Plasmids | Relevant Properties | Ref. | |
|---|---|---|---|
| Bacterial Strains | ID | ||
| BZB1011 | WT | F-, λ-, gyrA586(NalR), in(rrnD-rrnE)1, rpsL-(strR), rph−1 | [23] |
| Plasmids | |||
| pUArrnB | WT-GFP | pUArrnB::gfp, Knr | [24] |
| ColA-pUArrnB | A-GFP | caa, cai, cal (colicin A) & pUArrnB | [14] |
| PColE2 (pColE2-P9) | E2 | ce2a, ce2i, ce2l (colicin E2) | [23] |
| PColE7 (pColE7-K317) | E7 | ce7a, ce7i, ce7l (colicin E7) | [23] |
| PColE8 (pColE8-J) | E8 | ce8a, ce8i, ce8l (colicin E8) | [23] |
| Colicin Type | Rate of Colicin Expressing Cells (%) | |
|---|---|---|
| Late log | Stationary | |
| A | 2.7 ± 1.2 | 3.7 ± 1.5 |
| E2 | 18.3 ± 6.8 | 21.1 ± 8.5 |
| E7 | 3.4 ± 1.4 | 5.7 ± 0.5 |
| E8 | 14.6 ± 2.0 | 28.1 ± 6.5 |
| E. coli Strains | Growth Rate | ||
|---|---|---|---|
| Strain | Plasmid | ID | |
| BZB1011 | ColE2 | E2 | 0.34 ± 0.02 |
| ColE7 | E7 | 0.34 ± 0.03 | |
| ColE8 | E8 | 0.38 ± 0.07 | |
| pUArrnB | WT-GFP | 0.41 ± 0.07 | |
| ColA-pUArrnB | A-GFP | 0.35 ± 0.03 | |
| WT | 0.41 ± 0.06 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghazaryan, L.; Giladi, I.; Gillor, O. The Effects of Colicin Production Rates on Allelopathic Interactions in Escherichia coli Populations. Microorganisms 2019, 7, 564. https://doi.org/10.3390/microorganisms7110564
Ghazaryan L, Giladi I, Gillor O. The Effects of Colicin Production Rates on Allelopathic Interactions in Escherichia coli Populations. Microorganisms. 2019; 7(11):564. https://doi.org/10.3390/microorganisms7110564
Chicago/Turabian StyleGhazaryan, Lusine, Itamar Giladi, and Osnat Gillor. 2019. "The Effects of Colicin Production Rates on Allelopathic Interactions in Escherichia coli Populations" Microorganisms 7, no. 11: 564. https://doi.org/10.3390/microorganisms7110564
APA StyleGhazaryan, L., Giladi, I., & Gillor, O. (2019). The Effects of Colicin Production Rates on Allelopathic Interactions in Escherichia coli Populations. Microorganisms, 7(11), 564. https://doi.org/10.3390/microorganisms7110564