Analysis of the Indole Diterpene Gene Cluster for Biosynthesis of the Epoxy-Janthitrems in Epichloë Endophytes

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Growth Conditions
2.2. Identification of a Candidate Gene for Epoxy-Janthitrem Biosynthesis, jtmD, and the JTM Locus
2.2.1. DNA extraction and short-read sequencing on Illumina HiSeq2000 sequencing platform
2.2.2. DNA Extraction and Long-Read Sequencing on PacBio and MinIon Sequencing Platforms
2.2.3. Sequence Assembly and JTM Locus Identification
2.2.4. Transcriptome Expression of Genes Located Within the JTM Locus
2.3. jtmD Knockdown Constructs and Transformation
2.3.1. Fungal and Bacterial Strains and Culture Conditions
2.3.2. Plasmid Construction
2.3.3. Isolation of Fungal Protoplasts
2.3.4. PEG-mediated Fungal Protoplast Transformation
2.3.5. Fungal Protoplast Regeneration
2.3.6. Molecular Analysis of Transformed Endophytes
2.4. Generation and Analysis of Perennial Ryegrass Infected with jtmD Knockdown Strains
2.4.1. Seedling Inoculation of Transgenic Endophytes
2.4.2. Sample Preparation for Epoxy-Janthitrem Profiling
2.4.3. LC-MS Analysis
3. Results and Discussion
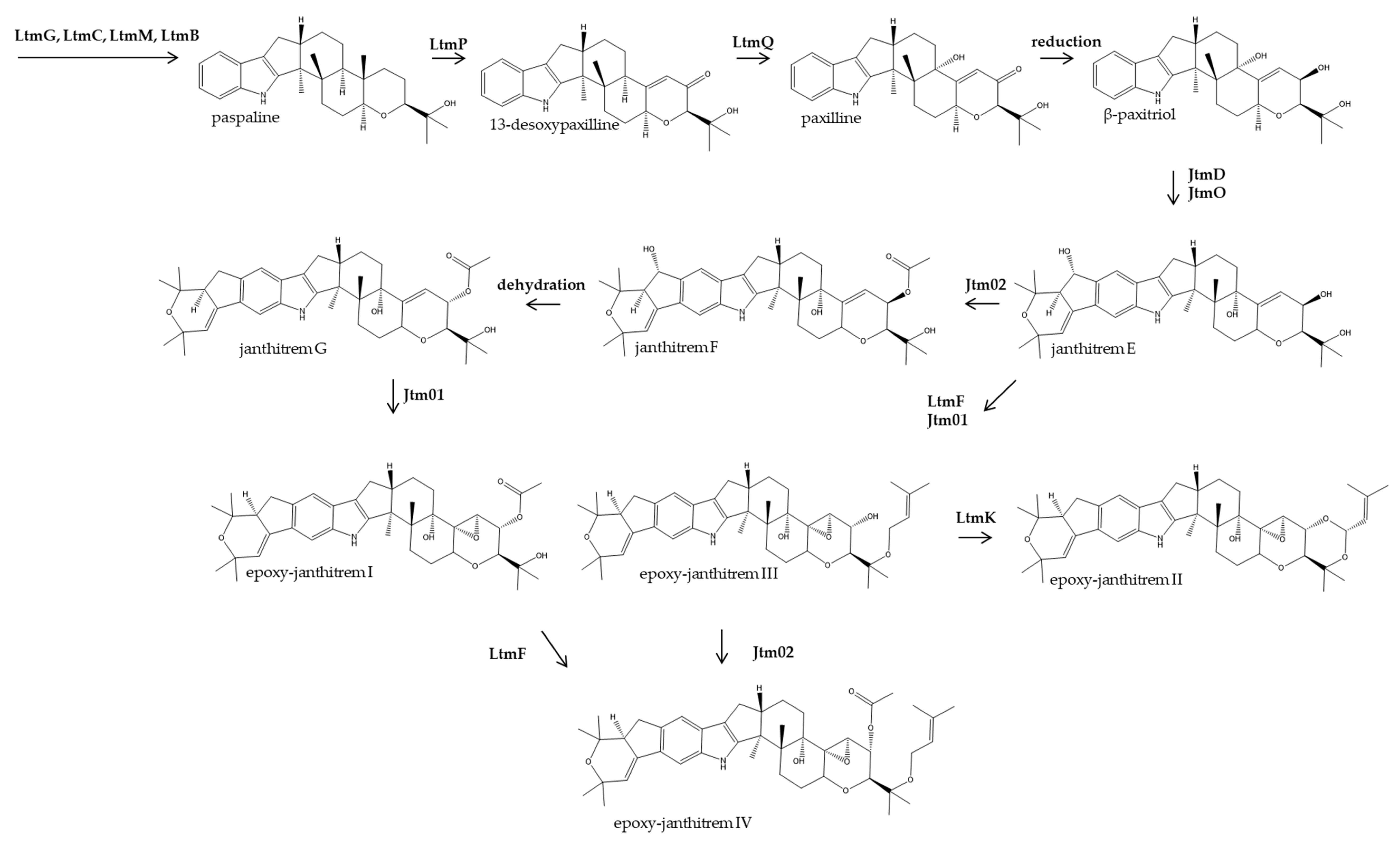
3.1. Identification of a Candidate Gene, jtmD, for Epoxy-Janthitrem Biosynthesis
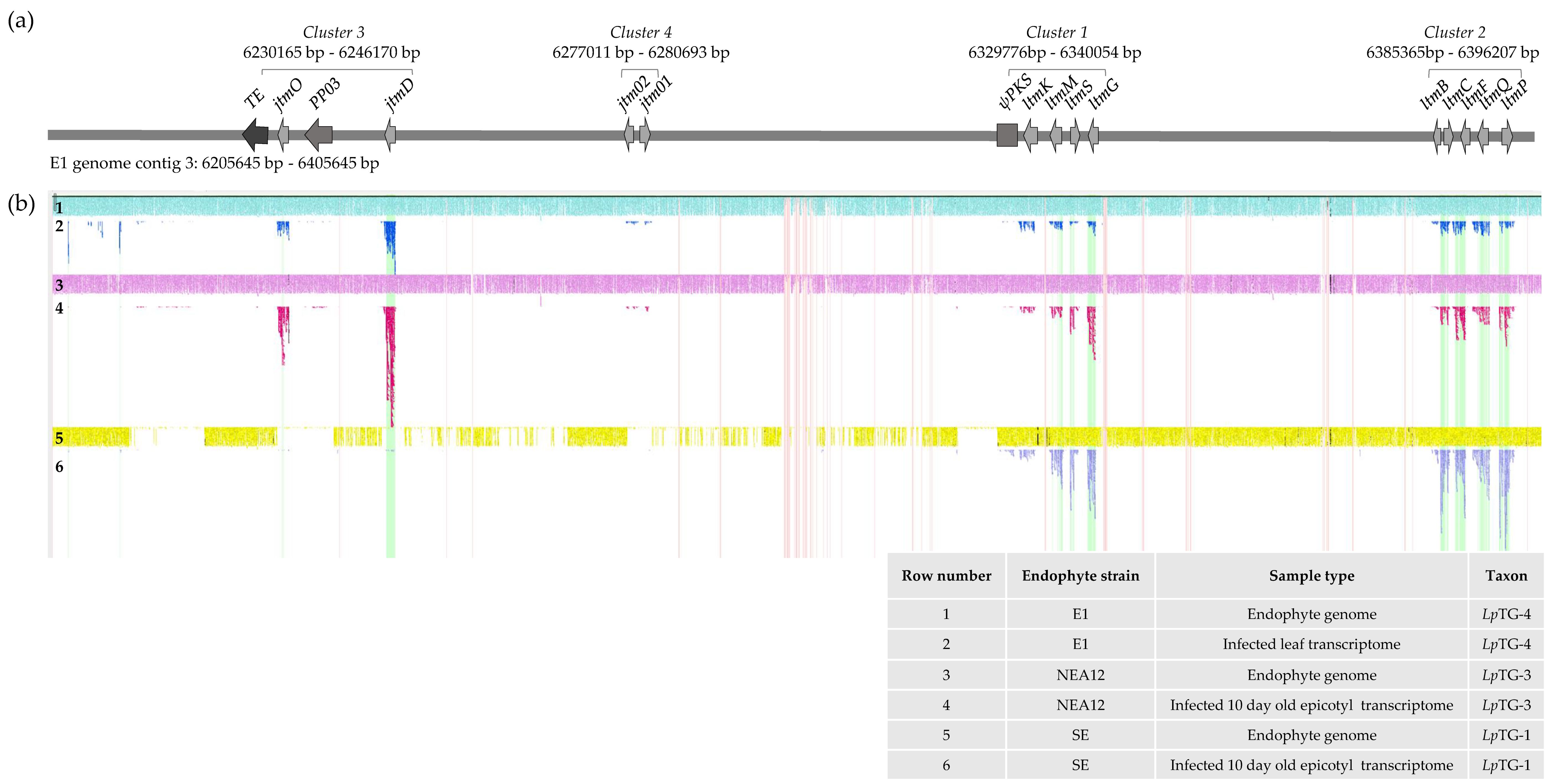
3.2. Identification of the JTM Locus in the LpTG-3 and LpTG-4 Genomes
3.3. Transcriptome Expression of Genes Located in the Janthitrem Biosynthesis Gene Cluster
3.4. Functional Analysis of jtmD to Determine Involvement in Epoxy-Janthitrem Biosynthesis
3.5. Proposed Pathway for Epoxy-Janthitrem Biosynthesis
4. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Scott, B.; Schardl, C.L. Fungal symbionts of grasses: Evolutionary insights and agricultural potential. Trends Microbiol. 1993, 1, 196–200. [Google Scholar] [CrossRef]
- Philippe, G. Lolitrem B and indole diterpene alkaloids produced by endophytic fungi of the genus Epichloë and their toxic effects in livestock. Toxins 2016, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Guerre, P. Ergot alkaloids produced by endophytic fungi of the genus Epichloë. Toxins 2015, 7, 773–790. [Google Scholar] [CrossRef] [PubMed]
- Christensen, M.J.; Leuchtmann, A.; Rowan, D.D.; Tapper, B.A. Taxonomy of Acremonium endophytes of tall fescue (Festuca arundinacea), meadow fescue (F. pratensis) and perennial ryegrass (Lolium perenne). Mycol. Res. 1993, 97, 1083–1092. [Google Scholar] [CrossRef]
- Campbell, M.A.; Tapper, B.A.; Simpson, W.R.; Johnson, R.D.; Mace, W.; Ram, A.; Lukito, Y.; Dupont, P.-Y.; Johnson, L.J.; Scott, D.B.; et al. Epichloë hybrida, sp. nov., an emerging model system for investigating fungal allopolyploidy. Mycologia 2017, 109, 715–729. [Google Scholar] [CrossRef]
- Hettiarachchige, I.K.; Ekanayake, P.N.; Mann, R.C.; Guthridge, K.M.; Sawbridge, T.I.; Spangenberg, G.C.; Forster, J.W. Phylogenomics of asexual Epichloë fungal endophytes forming associations with perennial ryegrass. BMC Evol. Biol. 2015, 15, 72. [Google Scholar] [CrossRef]
- Kaur, J.; Ekanayake, P.; Tian, P.; van Zijll de Jong, E.; Dobrowolski, M.P.; Rochfort, S.J.; Mann, R.; Smith, K.F.; Forster, J.W.; Guthridge, K.M.; et al. Discovery and characterisation of novel asexual Epichloë endophytes from perennial ryegrass (Lolium perenne L.). Crop. Pasture Sci. 2015, 66, 1058–1070. [Google Scholar] [CrossRef]
- Van Zijll de Jong, E.; Dobrowolski, M.P.; Bannan, N.R.; Stewart, A.V.; Smith, K.F.; Spangenberg, G.C.; Forster, J.W. Global genetic diversity of the perennial ryegrass fungal endophyte Neotyphodium lolii. Crop. Sci. 2008, 48, 1487–1501. [Google Scholar] [CrossRef]
- Popay, A.J.; Wyatt, R.T. Resistance to Argentine stem weevil in perennial ryegrass infected with endophytes producing different alkaloids. In Proceedings of the Forty Eighth New Zealand Plant Protection Conference, Rotaura, New Zealand, 8–10 August 1995; pp. 229–236. [Google Scholar]
- Spangenberg, G.C.; Guthridge, K.M.; Forster, J.W.; Sawbridge, T.I.; Ludlow, E.J.I.; Kaur, J.; Rochfort, S.J.; Rabinovich, M.A.; Ekanayake, P. Endophytes and Related Methods. WO2011082455A1, 2011. Available online: http://patents.google.com/patent/WO2011082455A1/en (accessed on 1 October 2019).
- Tapper, B.A.; Cooper, B.M.; Easton, H.S.; Fletcher, L.R.; Hume, D.E.; Lane, G.A.; Latch, G.C.M.; Pennell, C.G.L.; Popay, A.J.; Christensen, M.J. Grass Endophytes. WO2004106487A2, 2004. Available online: https://patents.google.com/patent/WO2004106487A2/un (accessed on 1 October 2019).
- Finch, S.C.; Fletcher, L.R.; Babu, J.V. The evaluation of endophyte toxin residues in sheep fat. N. Z. Vet. J. 2011, 60, 56–60. [Google Scholar] [CrossRef]
- Fletcher, L.R.; Sutherland, B.L. Sheep responses to grazing ryegrass with AR37 endophyte. Proc. N. Z. Grassl. Assoc. 2009, 71, 127–132. [Google Scholar]
- Fletcher, L.R. Managing ryegrass-endophyte toxicoses. In Neotyphodium in Cool-Season Grasses; West, C.P., Roberts, C.A., Spiers, D.E., Eds.; Blackwell Publishing: Hoboken, NJ, USA, 2005; pp. 227–241. [Google Scholar]
- Babu, J.V.; Popay, A.J.; Miles, C.O.; Wilkins, A.L.; di Menna, M.E.; Finch, S.C. Identification and structure elucidation of janthitrems A and D from Penicillium janthinellum and determination of the tremorgenic and anti-insect activity of janthitrems A and B. J. Agric. Food Chem. 2018, 66, 13116–13125. [Google Scholar] [CrossRef] [PubMed]
- Hennessy, L.M.; Popay, A.J.; Finch, S.C.; Clearwater, M.J.; Cave, V.M. Temperature and plant genotype alter alkaloid concentrations in ryegrass infected with an Epichloë endophyte and this affects an insect herbivore. Front. Plant. Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Thom, E.R.; Popay, A.J.; Waugh, C.D.; Minneé, E.M.K. Impact of novel endophytes in perennial ryegrass on herbage production and insect pests from pastures under dairy cow grazing in northern New Zealand. Grass Forage Sci. 2014, 69, 191–204. [Google Scholar] [CrossRef]
- Karpyn Esqueda, M.; Yen, A.L.; Rochfort, S.; Guthridge, K.M.; Powell, K.S.; Edwards, J.; Spangenberg, G.C. A Review of perennial ryegrass endophytes and their potential use in the management of african black beetle in perennial grazing systems in Australia. Front. Plant. Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, R.T.; Latch, G.C.M.; Keogh, R.G. The janthitrems: Fluorescent tremorgenic toxins produced by Penicillium janthinellum isolates from ryegrass pastures. Appl. Environ. Microbiol. 1980, 39, 272–273. [Google Scholar]
- de Jesus, A.E.; Steyn, P.S.; van Heerden, F.R.; Vleggaar, R. Structure elucidation of the janthitrems, novel tremorgenic mycotoxins from Penicillium janthinellum. J. Chem. Soc. Perkin Trans. 1 1984, 697–701. [Google Scholar] [CrossRef]
- Rowan, D.D. Lolitrems, peramine and paxilline: Mycotoxins of the ryegrass/endophyte interactions. Agric. Ecosyst. Environ. 1993, 44, 103–122. [Google Scholar] [CrossRef]
- Mantle, P.G.; Penn, J. A role for paxilline in the biosynthesis of indole–diterpenoid penitrem mycotoxins. J. Chem. Soc. Perkin Trans. 1 1989, 1539–1540. [Google Scholar] [CrossRef]
- Penn, J.; Mantle, P.G. Biosynthetic intermediates of indole-diterpenoid mycotoxins from selected transformations at C-10 of paxilline. Phytochemistry 1994, 35, 921–926. [Google Scholar] [CrossRef]
- Schardl, C.L.; Young, C.A.; Faulkner, J.R.; Florea, S.; Pan, J. Chemotypic diversity of Epichloë, fungal symbionts of grasses. Fungal Ecol. 2012, 5, 331–344. [Google Scholar] [CrossRef]
- Kozák, L.; Szilágyi, Z.; Tóth, L.; Pócsi, I.; Molnár, I. Tremorgenic and neurotoxic paspaline-derived indole diterpenes: Biosynthetic diversity, threats and applications. Appl. Microbiol. Biotechnol. 2019, 103, 1599–1616. [Google Scholar] [CrossRef] [PubMed]
- Laws, I.; Mantle, P.G. Experimental constraints in the study of the biosynthesis of indole alkaloids in fungi. Microbiology 1989, 135, 2679–2692. [Google Scholar] [CrossRef][Green Version]
- Parker, E.J.; Scott, D.B. Indole-diterpene biosynthesis in ascomycetous fungi. In Handbook of Industrial Mycology; An, Z., Ed.; CRC Press: Boca Raton, FL, USA, 2004; pp. 405–426. [Google Scholar]
- Reddy, P.; Deseo, M.A.; Ezernieks, V.; Guthridge, K.; Spangenberg, G.; Rochfort, S. Toxic indole diterpenes from endophyte-infected perennial ryegrass Lolium perenne L.: Isolation and stability. Toxins 2019, 11, 16. [Google Scholar] [CrossRef] [PubMed]
- Young, C.; McMillan, L.; Telfer, E.; Scott, B. Molecular cloning and genetic analysis of an indole-diterpene gene cluster from Penicillium paxilli. Mol. Microbiol. 2001, 39, 754–764. [Google Scholar] [CrossRef]
- Saikia, S.; Parker, E.J.; Koulman, A.; Scott, B. Defining paxilline biosynthesis in Penicillium paxilli: Functional characterisation of two cytochrome P450 monoxygenases. J. Biol. Chem. 2007, 282, 16829–16837. [Google Scholar] [CrossRef]
- Scott, B.; Young, C.A.; Saikia, S.; McMillan, L.K.; Monahan, B.J.; Koulman, A.; Astin, J.; Eaton, C.J.; Bryant, A.; Wrenn, R.E.; et al. Deletion and gene expression analyses define the paxilline biosynthetic gene cluster in Penicillium paxilli. Toxins 2013, 5, 1422–1446. [Google Scholar] [CrossRef]
- Zhang, S.; Monahan, B.J.; Tkacz, J.S.; Scott, B. Indole-diterpene gene cluster from Aspergillus flavus. Appl. Environ. Microbiol. 2004, 70, 6875–6883. [Google Scholar] [CrossRef]
- Nicholson, M.J.; Eaton, C.J.; Stärkel, C.; Tapper, B.A.; Cox, M.P.; Scott, B. Molecular cloning and functional analysis of gene clusters for the biosynthesis of indole diterpenes in Penicillium crustosum and P. janthinellum. Toxins 2015, 7, 2701–2722. [Google Scholar] [CrossRef]
- Saikia, S.; Takemoto, D.; Tapper, B.A.; Lane, G.A.; Fraser, K.; Scott, B. Functional analysis of an indole-diterpene gene cluster for lolitrem B biosynthesis in the grass endosymbiont Epichloë festucae. FEBS Lett. 2012, 586, 2563–2569. [Google Scholar] [CrossRef]
- Young, C.A.; Felitti, S.; Shields, K.; Spangenberg, G.; Johnson, R.D.; Bryan, G.T.; Saikia, S.; Scott, B. A complex gene cluster for indole-diterpene biosynthesis in the grass endophyte Neotyphodium lolii. Fungal Genet. Biol. 2006, 43, 679–693. [Google Scholar] [CrossRef]
- Motoyama, T.; Hayashi, T.; Hirota, H.; Ueki, M.; Osada, H. Terpendole E, a kinesin Eg5 inhibitor, Is a key biosynthetic intermediate of indole diterpenes in the producing fungus Chaunopycnis alba. Chem. Biol. 2012, 19, 1611–1619. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Tagami, K.; Minami, A.; Matsumoto, T.; Frisvad, J.C.; Suzuki, H.; Ishikawa, J.; Gomi, K.; Oikawa, H. Reconstitution of biosynthetic machinery for the synthesis of the highly elaborated indole diterpene penitrem. Angew. Chem. Int. Ed. 2015, 54, 5748–5752. [Google Scholar] [CrossRef] [PubMed]
- Young, C.A.; Bryant, M.K.; Christensen, M.J.; Tapper, B.A.; Bryan, G.T.; Scott, B. Molecular cloning and genetic analysis of a symbiosis-expressed gene cluster for lolitrem biosynthesis from a mutualistic endophyte of perennial ryegrass. Mol. Genet. Genom. 2005, 274, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Fleetwood, D.J.; Scott, B.; Lane, G.A.; Tanaka, A.; Johnson, R.D. A complex ergovaline gene cluster in Epichloë endophytes of grasses. Appl. Environ. Microbiol. 2007, 73, 2571–2579. [Google Scholar] [CrossRef] [PubMed]
- Möller, E.M.; Bahnweg, G.; Sandermann, H.; Geiger, H.H. A simple and efficient protocol for isolation of high molecular weight DNA from filamentous fungi, fruit bodies, and infected plant tissues. Nucleic Acids Res. 1992, 20, 6115–6116. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.-Y.J.; Fan, W.-L.; Wang, W.-F.; Chen, T.; Tang, Y.-C.; Chu, F.-H.; Chang, T.-T.; Wang, S.-Y.; Li, M.-Y.; Chen, Y.-H.; et al. Genomic and transcriptomic analyses of the medicinal fungus Antrodia cinnamomea for its metabolite biosynthesis and sexual development. Proc. Natl. Acad. Sci. USA 2014, 111, E4743–E4752. [Google Scholar] [CrossRef]
- Chin, C.-S.; Alexander, D.H.; Marks, P.; Klammer, A.A.; Drake, J.; Heiner, C.; Clum, A.; Copeland, A.; Huddleston, J.; Eichler, E.E.; et al. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat. Methods 2013, 10, 563–569. [Google Scholar] [CrossRef]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Stanke, M.; Morgenstern, B. AUGUSTUS: A web server for gene prediction in eukaryotes that allows user-defined constraints. Nucleic Acids Res. 2005, 33, W465–W467. [Google Scholar] [CrossRef]
- Edgar, R. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Hettiarachchige, I.K.; Elkins, A.C.; Reddy, P.; Mann, R.C.; Guthridge, K.M.; Sawbridge, T.I.; Forster, J.W.; Spangenberg, G.C. Genetic modification of asexual Epichloë endophytes with the perA gene for peramine biosynthesis. Mol. Genet. Genom. 2019, 294, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Shinozuka, H.; Cogan, N.O.I.; Spangenberg, G.C.; Forster, J.W. Reference transcriptome assembly and annotation for perennial ryegrass. Genome 2017, 60, 1086–1088. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed]
- McMillan, L.K.; Carr, R.L.; Young, C.A.; Astin, J.W.; Lowe, R.G.T.; Parker, E.J.; Jameson, G.B.; Finch, S.C.; Miles, C.O.; McManus, O.B.; et al. Molecular analysis of two cytochrome P450 monooxygenase genes required for paxilline biosynthesis in Penicillium paxilli, and effects of paxilline intermediates on mammalian maxi-K ion channels. Mol. Genet. Genom. 2003, 270, 9–23. [Google Scholar] [CrossRef]
- Liu, C.; Minami, A.; Dairi, T.; Gomi, K.; Scott, B.; Oikawa, H. Biosynthesis of shearinine: Diversification of a tandem prenyl moiety of fungal indole diterpenes. Org. Lett. 2016, 18, 5026–5029. [Google Scholar] [CrossRef]
- Van de Bittner, K.C.; Nicholson, M.J.; Bustamante, L.Y.; Kessans, S.A.; Ram, A.; van Dolleweerd, C.J.; Scott, B.; Parker, E.J. Heterologous biosynthesis of nodulisporic acid F. J. Am. Chem. Soc. 2018, 140, 582–585. [Google Scholar] [CrossRef]
- Winter, D.J.; Ganley, A.R.D.; Young, C.A.; Liachko, I.; Schardl, C.L.; Dupont, P.-Y.; Berry, D.; Ram, A.; Scott, B.; Cox, M.P. Repeat elements organise 3D genome structure and mediate transcription in the filamentous fungus Epichloë festucae. PLoS Genet. 2018, 14, e1007467. [Google Scholar] [CrossRef]
- Ekanayake, P.N.; Kaur, J.; Tian, P.; Rochfort, S.J.; Guthridge, K.M.; Sawbridge, T.I.; Spangenberg, G.C.; Forster, J.W. Genomic and metabolic characterisation of alkaloid biosynthesis by asexual Epichloë fungal endophytes of tall fescue pasture grasses. Genome 2017, 60, 496–509. [Google Scholar] [CrossRef] [PubMed]
- Young, C.; Tapper, B.; May, K.; Moon, C.; Schardl, C.; Scott, B. Indole-diterpene biosynthetic capability of Epichloë endophytes as predicted by ltm gene analysis. Appl. Environ. Microbiol. 2009, 75, 2200–2211. [Google Scholar] [CrossRef] [PubMed]
- Chujo, T.; Lukito, Y.; Eaton, C.J.; Dupont, P.-Y.; Johnson, L.J.; Winter, D.; Cox, M.P.; Scott, B. Complex epigenetic regulation of alkaloid biosynthesis and host interaction by heterochromatin protein I in a fungal endophyte-plant symbiosis. Fungal Genet. Biol. 2019, 125, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Saikia, S.; Nicholson, M.J.; Young, C.; Parker, E.J.; Scott, B. The genetic basis for indole diterpene chemical diversity in filamentous fungi. Mycol. Res. 2008, 112, 184–199. [Google Scholar] [CrossRef] [PubMed]
- de Man, T.J.B.; Stajich, J.E.; Kubicek, C.P.; Teiling, C.; Chenthamara, K.; Atanasova, L.; Druzhinina, I.S.; Levenkova, N.; Birnbaum, S.S.L.; Barribeau, S.M.; et al. Small genome of the fungus Escovopsis weberi, a specialized disease agent of ant agriculture. Proc. Natl. Acad. Sci. USA 2016, 113, 3567–3572. [Google Scholar] [CrossRef]
- Dhodary, B.; Schilg, M.; Wirth, R.; Spiteller, D. Secondary metabolites from Escovopsis weberi and their role in attacking the garden fungus of leaf-cutting ants. Chem. A Eur. J. 2018, 24, 4445–4452. [Google Scholar] [CrossRef]
- Boya P., C.A.; Fernández-Marín, H.; Mejía, L.C.; Spadafora, C.; Dorrestein, P.C.; Gutiérrez, M. Imaging mass spectrometry and MS/MS molecular networking reveals chemical interactions among cuticular bacteria and pathogenic fungi associated with fungus-growing ants. Sci. Rep. 2017, 7, 5604. [Google Scholar] [CrossRef]
- Bills, G.F.; González-Menéndez, V.; Martín, J.; Platas, G.; Fournier, J.; Peršoh, D.; Stadler, M. Hypoxylon pulicicidum sp. nov. (Ascomycota, Xylariales), a pantropical insecticide-producing endophyte. PLoS ONE 2012, 7, e46687. [Google Scholar] [CrossRef]
- Ondeyka, J.G.; Helms, G.L.; Hensens, O.D.; Goetz, M.A.; Zink, D.L.; Tsipouras, A.; Shoop, W.L.; Slayton, L.; Dombrowski, A.W.; Polishook, J.D.; et al. Nodulisporic acid A, a novel and potent insecticide from a Nodulisporium sp. isolation, structure determination, and chemical transformations. J. Am. Chem. Soc. 1997, 119, 8809–8816. [Google Scholar] [CrossRef]
- Andersen, J.F.; Walding, J.K.; Evans, P.H.; Bowers, W.S.; Feyereisen, R. Substrate specificity for the epoxidation of terpenoids and active site topology of house fly cytochrome P450 6A1. Chem. Res. Toxicol. 1997, 10, 156–164. [Google Scholar] [CrossRef]
- Lai, Y.; Liu, K.; Zhang, X.; Zhang, X.; Li, K.; Wang, N.; Shu, C.; Wu, Y.; Wang, C.; Bushley, K.E.; et al. Comparative genomics and transcriptomics analyses reveal divergent lifestyle features of nematode endoparasitic fungus Hirsutella minnesotensis. Genome Biol. Evol. 2014, 6, 3077–3093. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon a | E. festucae var. lolii (LpTG-1) | LpTG-3 | LpTG-4 |
|---|---|---|---|
| Endophyte strain | SE | NEA12 | E1 |
| Alkaloid biosynthesis profile b | lolitrem B, ergovaline, peramine | epoxy-janthitrems | epoxy-janthitrems |
| Position in JTM Cluster | Top BLASTp Hit | ||||||
|---|---|---|---|---|---|---|---|
| Feature ID | Gene Cluster | Predicted Function | Homologous Gene | Percent Identity (aa) | Organism | Genbank Accession No. | Reference |
| TE | - | Transposable element | 85% | Hirsutella minnesotensis | KJZ70955 | [66] | |
| jtmO | 3 | FAD-dependent oxygenase | nodO | 70% | Hypoxylon pulicicidum | AUM60052.1 | [54] |
| PP03 | 3 | Transposase | hypothetical protein | 86% | Hirsutella minnesotensis | KJZ68513 | [66] |
| jtmD | 3 | Aromatic prenyl transferase | nodD1 | 67% | Hypoxylon pulicicidum | AUM60056.1 | [54] |
| jtm02 | 4 | Membrane bound O-acyl transferase | hypothetical protein | 34% | Oidiodendron maius Zn | KIM95229 | unpublished |
| jtm01 | 4 | Cytochrome P450 monooxygenase | hypothetical protein | 68% | Hirsutella minnesotensis | KJZ77225 | [66] |
| ψpks | - | Polyketide synthase (pseudogene) | 73% | Fusarium equiseti | ALQ32965.1 | unpublished | |
| ltmK | 1 | Cytochrome P450 monooxygenase | ltmK | 99% | LpTG-1 | AY742903 | [38] |
| ltmM | 1 | FAD-dependent monooxygenase | ltmM | 99% | LpTG-1 | AY742903 | [38] |
| ltmS | 1 | Integral membrane protein | ltmS | 100% | LpTG-1 | AY742903 | [38] |
| ltmG | 1 | GGPP synthase | ltmG | 99% | LpTG-1 | AY742903 | [38] |
| ltmB | 2 | Integral membrane protein | ltmB | 100% | LpTG-1 | DQ443465 | [35] |
| ltmC | 2 | Prenyl transferase | ltmC | 100% | LpTG-1 | DQ443465 | [35] |
| ltmF | 2 | Prenyl transferase | ltmF | 99% | LpTG-1 | DQ443465 | [35] |
| ltmQ | 2 | Cytochrome P450 monooxygenase | ltmQ | 100% | LpTG-1 | DQ443465 | [35] |
| ltmP | 2 | Cytochrome P450 monooxygenase | ltmP | 100% | LpTG-1 | DQ443465 | [35] |
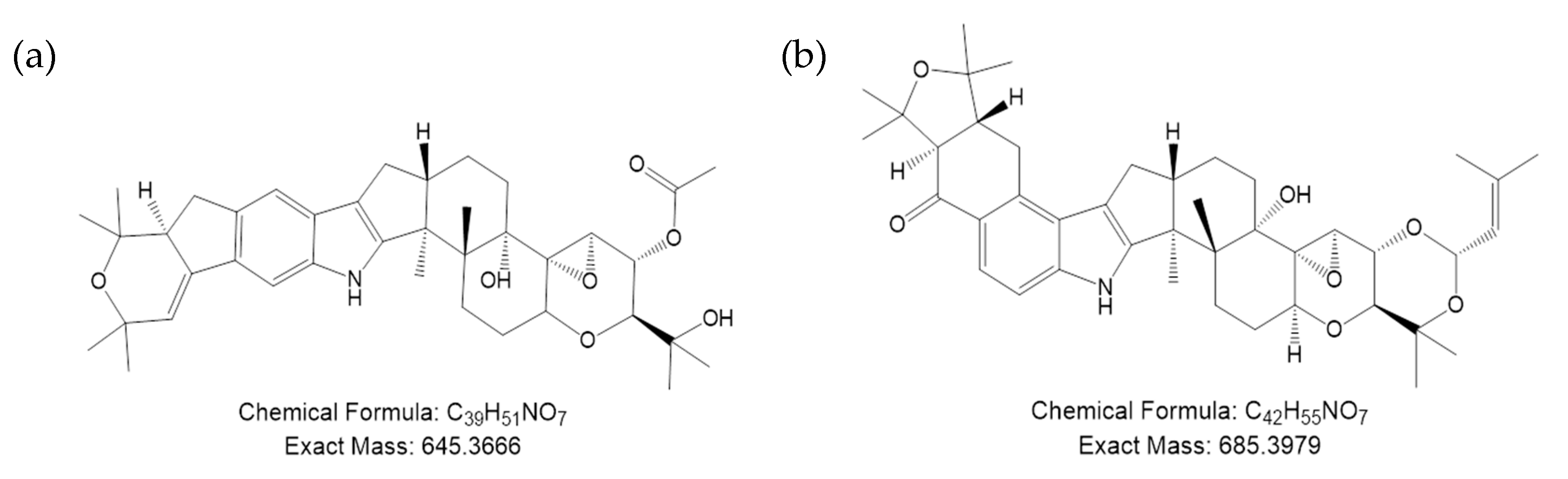
| Metabolite | RT (min) | m/z (M+H) | Product Ions (MS2) | Formula (M+H) | Theoretical Mass (M+H) | Δ (ppm) | |||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ||||||
| Paxilline * | 9.76 | 436.2481 | 130.0653 | 182.0966 | 288.9844 | 418.2376 | C27 H34 O4 N | 436.2482 | −0.36 |
| epoxy-janthitrem I | 11.07 | 646.3726 | 222.1276 | 280.1694 | 588.3315 | 631.3452 | C39 H52 O7 N | 646.3738 | −1.81 |
| epoxy-janthitrem II | 12.22 | 670.4078 | 222.1277 | 280.1693 | 612.3679 | 655.3818 | C42 H56 O6 N | 670.4102 | −3.63 |
| epoxy-janthitrem III | 12.37 | 672.4230 | 222.1277 | 280.1687 | 614.3801 | 657.3967 | C42 H58 O6 N | 672.4259 | −4.31 |
| epoxy-janthitrem IV | 12.35 | 714.4329 | 222.1276 | 280.1695 | 656.3939 | 699.4077 | C44 H60 O7 N | 714.4364 | −3.96 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ludlow, E.J.; Vassiliadis, S.; Ekanayake, P.N.; Hettiarachchige, I.K.; Reddy, P.; Sawbridge, T.I.; Rochfort, S.J.; Spangenberg, G.C.; Guthridge, K.M. Analysis of the Indole Diterpene Gene Cluster for Biosynthesis of the Epoxy-Janthitrems in Epichloë Endophytes. Microorganisms 2019, 7, 560. https://doi.org/10.3390/microorganisms7110560
Ludlow EJ, Vassiliadis S, Ekanayake PN, Hettiarachchige IK, Reddy P, Sawbridge TI, Rochfort SJ, Spangenberg GC, Guthridge KM. Analysis of the Indole Diterpene Gene Cluster for Biosynthesis of the Epoxy-Janthitrems in Epichloë Endophytes. Microorganisms. 2019; 7(11):560. https://doi.org/10.3390/microorganisms7110560
Chicago/Turabian StyleLudlow, Emma J., Simone Vassiliadis, Piyumi N. Ekanayake, Inoka K. Hettiarachchige, Priyanka Reddy, Tim I. Sawbridge, Simone J. Rochfort, German C. Spangenberg, and Kathryn M. Guthridge. 2019. "Analysis of the Indole Diterpene Gene Cluster for Biosynthesis of the Epoxy-Janthitrems in Epichloë Endophytes" Microorganisms 7, no. 11: 560. https://doi.org/10.3390/microorganisms7110560
APA StyleLudlow, E. J., Vassiliadis, S., Ekanayake, P. N., Hettiarachchige, I. K., Reddy, P., Sawbridge, T. I., Rochfort, S. J., Spangenberg, G. C., & Guthridge, K. M. (2019). Analysis of the Indole Diterpene Gene Cluster for Biosynthesis of the Epoxy-Janthitrems in Epichloë Endophytes. Microorganisms, 7(11), 560. https://doi.org/10.3390/microorganisms7110560