The Microbial Community of a Terrestrial Anoxic Inter-Tidal Zone: A Model for Laboratory-Based Studies of Potentially Habitable Ancient Lacustrine Systems on Mars

 , and
, and
Abstract
1. Introduction
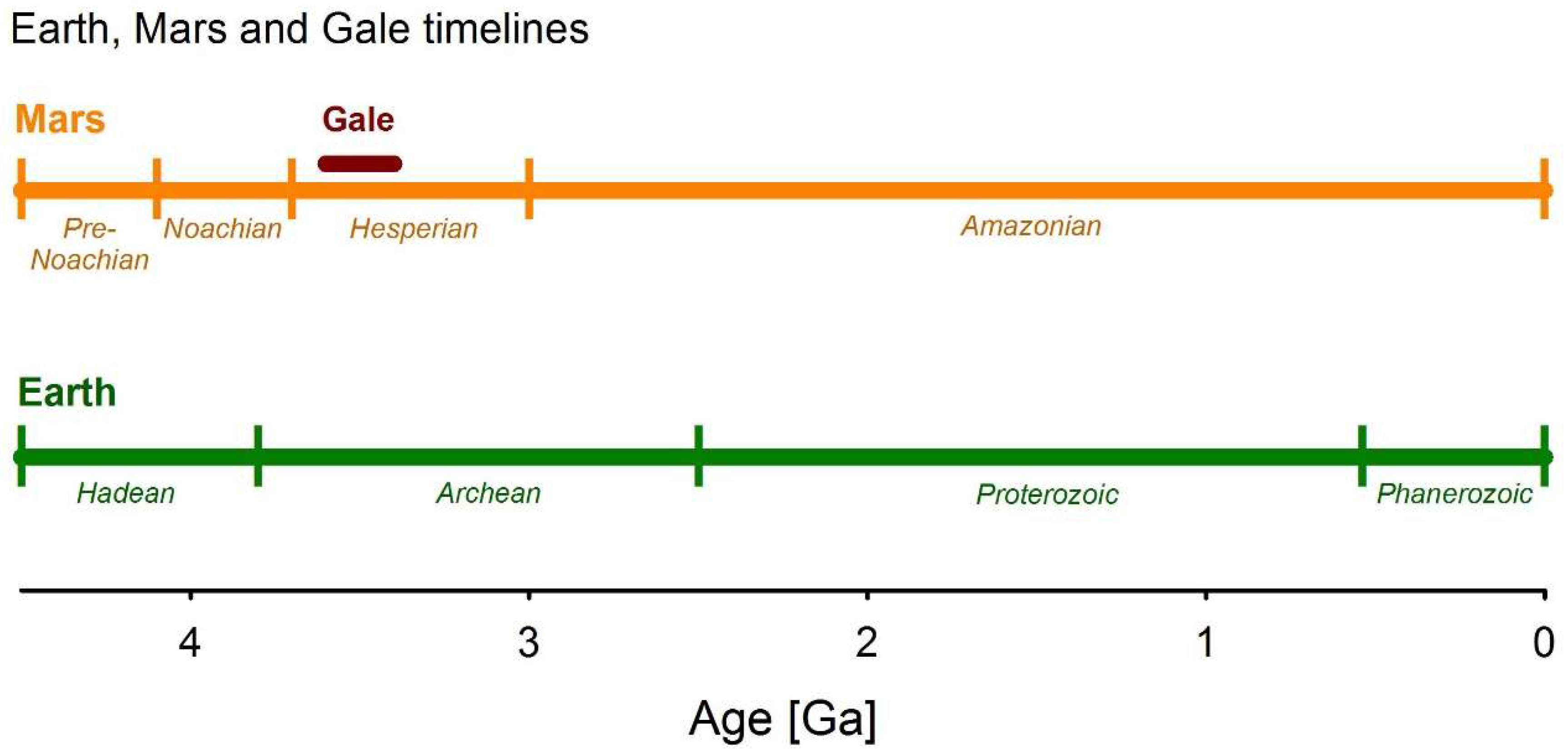
1.1. Geology and Lake Evolution at Gale Crater
1.2. Microbes in Analogue Systems
1.3. Comparing and Contrasting the Analogue Site Prerequisites to Gale Crater
2. Materials and Methods
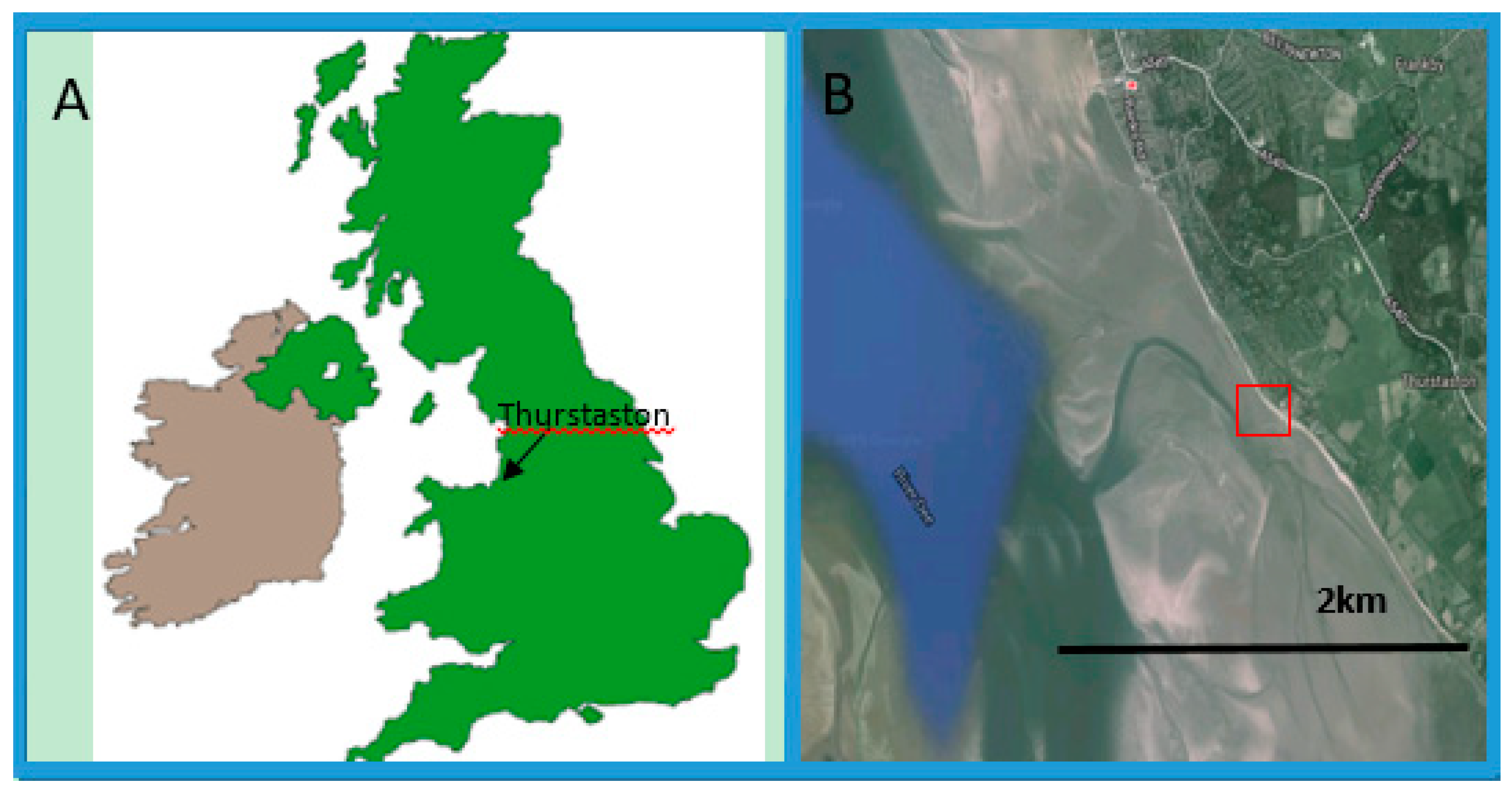
2.1. Sample Site and Sample Collection
2.2. Temperature, pH and TOC
2.3. Mineralogy
2.4. Cell Enumeration
2.5. Microbial Community Analysis
2.6. Isolation of Microorganisms
2.7. Microbial Growth Experiments
2.8. Statistical Analyse
3. Results
3.1. Temperature, pH and TOC
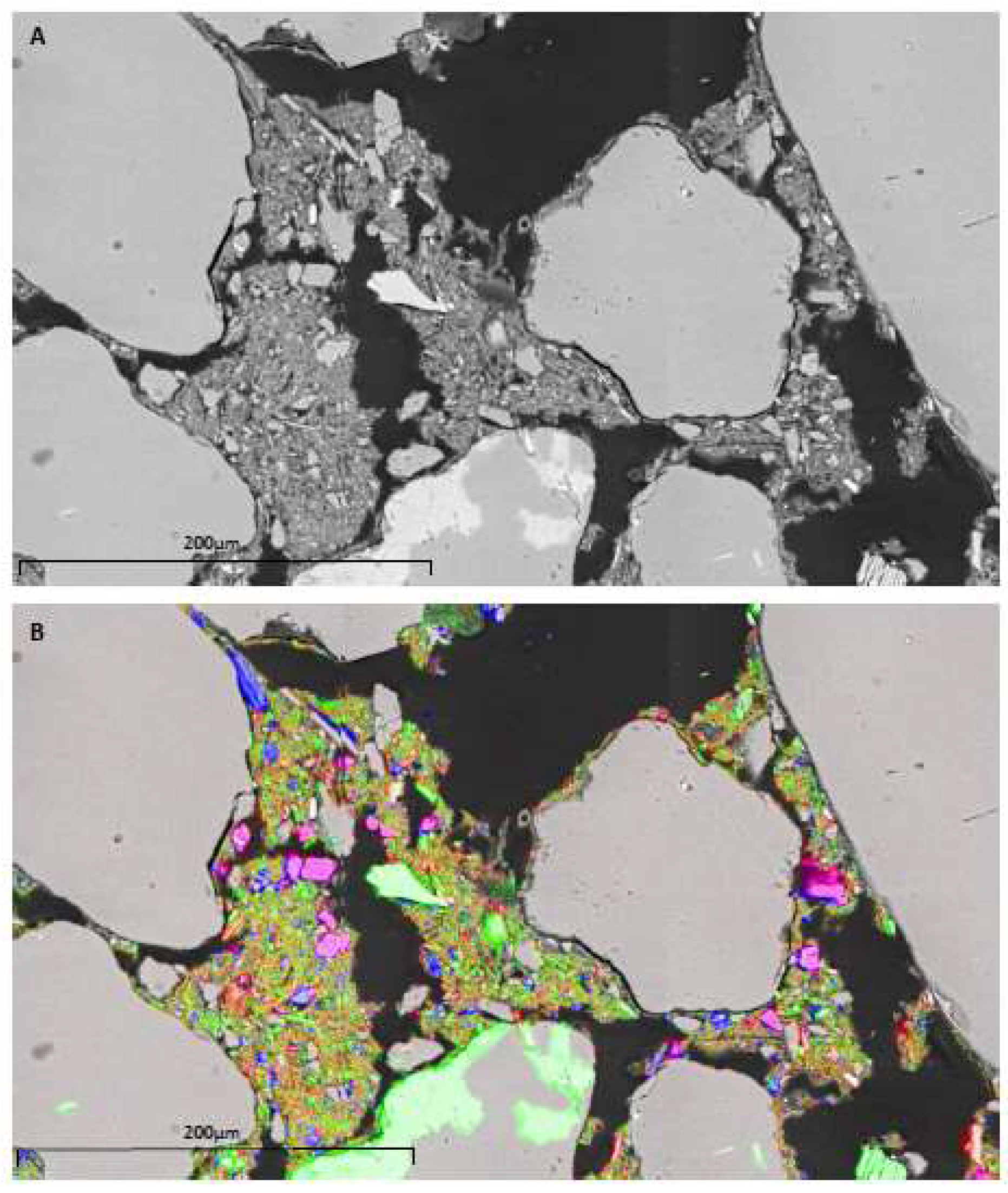
3.2. Mineralogy
3.3. Cell Enumeration
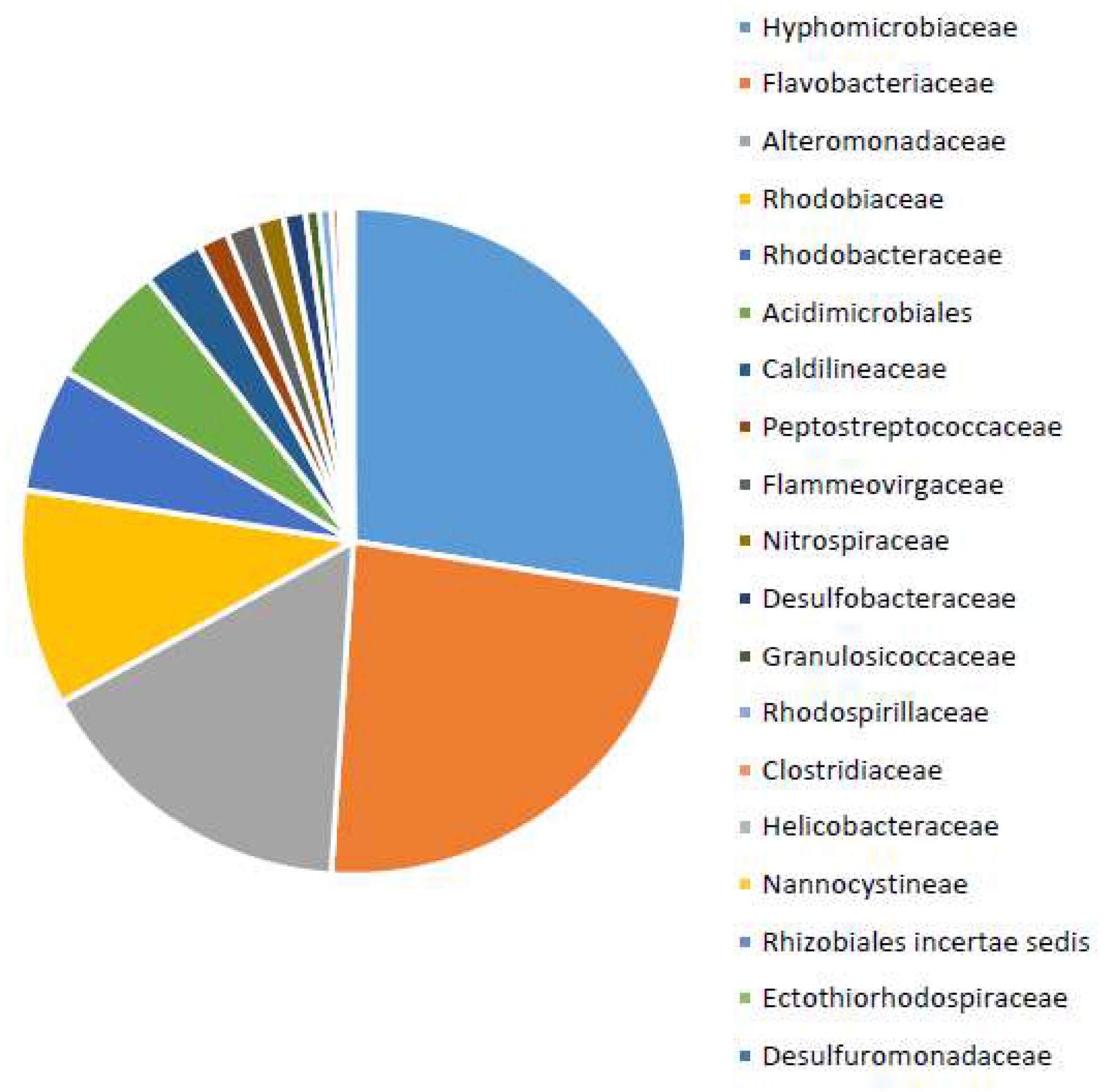
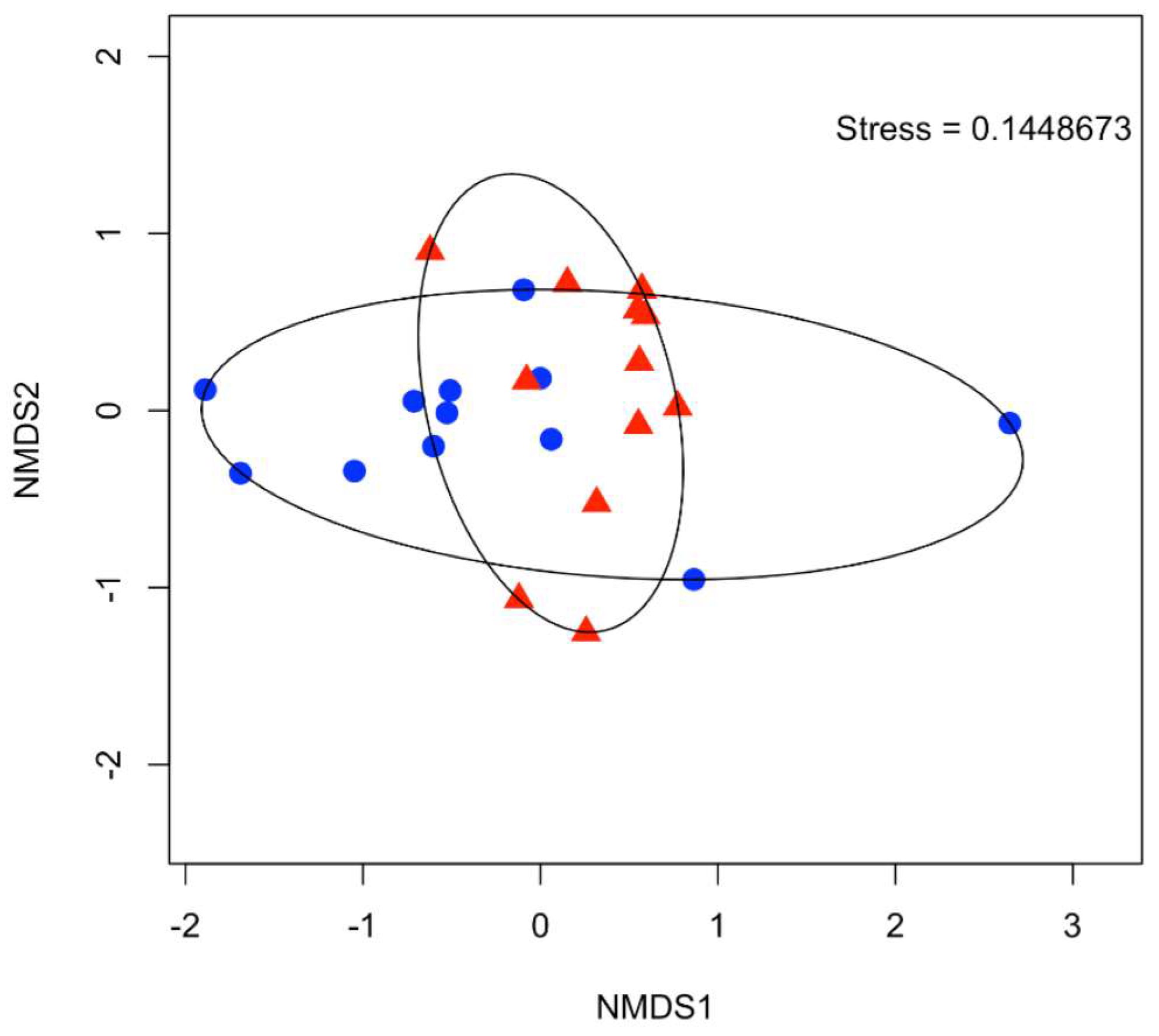
3.4. Community Analysis
3.5. Microbial Isolates
4. Discussion
4.1. An Analogue for the Fluvio-Lacustrine System at Gale Crater?
4.2. An Analogue for Putative Life in Fluvio-Lacustrine System at Gale Crater?
5. Summary and Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Molina-Cuberos, G.J.; Stumptner, W.; Lammer, H.; Komle, N.I. Cosmic ray and UV radiation models on the ancient martian surface. Icarus 2001, 154, 216–222. [Google Scholar] [CrossRef]
- Bibring, J.-P.; Langevin, Y.; Mustard, J.F.; Poulet, F.; Arvidson, R.; Gendrin, A.; Gondet, B.; Mangold, N.; Pinet, P.; Forget, F.; et al. Global mineralogical and aqueous mars history derived from OMEGA/Mars express data. Science 2006, 312, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Kasting, J.F.; Solomon, S.C. Thermal escape of carbon from the early Martian atmosphere. Geophys. Res. Lett. 2009, 36. [Google Scholar] [CrossRef]
- Warner, N.; Gupta, S.; Lin, S.Y.; Kim, J.R.; Muller, J.P.; Morley, J. Late Noachian to Hesperian climate change on Mars: Evidence of episodic warming from transient crater lakes near Ares Vallis. J. Geophys. Res. Planets 2011, 116. [Google Scholar] [CrossRef]
- Malin, M.C.; Edgett, K.S. Evidence for persistent flow and aqueous sediments on early Mars. Science 2003, 302, 1931–1934. [Google Scholar] [CrossRef] [PubMed]
- Irwin, R.P.; Howard, A.D.; Craddock, R.A.; Moore, J.M. An intense terminal epoch of widespread fluvial activity on early Mars: 2. Increased runoff and paleolake development. J. Geophys. Res. Planets 2005, 110. [Google Scholar] [CrossRef]
- Mangold, N.; Kite, E.S.; Kleinhans, M.G.; Newsom, H.; Ansan, V.; Hauber, E.; Kraal, E.; Quantin, C.; Tanaka, K. The origin and timing of fluvial activity at Eberswalde crater, Mars. Icarus 2012, 220, 530–551. [Google Scholar] [CrossRef]
- Williams, R.M.; Grotzinger, J.P.; Dietrich, W.E.; Gupta, S.; Sumner, D.Y.; Wiens, R.C.; Mangold, N.; Malin, M.C.; Edgett, K.S.; Maurice, S.; et al. Martian fluvial conglomerates at Gale crater. Science 2013, 340, 1068–1072. [Google Scholar] [CrossRef] [PubMed]
- Fassett, C.I.; Head, J.W. Fluvial sedimentary deposits on Mars: Ancient deltas in a crater lake in the Nili Fossae region. Geophys. Res. Lett. 2015, 32. [Google Scholar] [CrossRef]
- Grotzinger, J.P.; Sumner, D.Y.; Kah, L.C.; Stack, K.; Gupta, S.; Edgar, L.; Rubin, D.; Lewis, K.; Schieber, J.; Mangold, N.; et al. A habitable fluvio-lacustrine environment at Yellowknife Bay, Gale crater, Mars. Science 2014, 343, 1242777. [Google Scholar] [CrossRef] [PubMed]
- McLennan, S.M.; Anderson, R.B.; Bell, J.F., III; Bridges, J.C.; Calef, F.; Campbell, J.L.; Clark, B.C.; Clegg, S.; Conrad, P.; Cousin, A.; et al. Elemental geochemistry of sedimentary rocks at Yellowknife Bay, Gale crater, Mars. Science 2014, 343, 1244734. [Google Scholar] [CrossRef] [PubMed]
- Grotzinger, J.P.; Gupta, S.; Malin, M.C.; Rubin, D.M.; Schieber, J.; Siebach, K.; Sumner, D.Y.; Stack, K.M.; Vasavada, A.R.; Arvidson, R.E.; et al. Deposition, exhumation, and paleoclimate of an ancient lake deposit, Gale crater, Mars. Science 2015, 350, aac7575. [Google Scholar] [CrossRef] [PubMed]
- Martín-Torres, F.J.; Zorzano, M.-P.; Valentín-Serrano, P.; Harri, A.-M.; Genzer, M.; Kemppinen, O.; Rivera-Valentin, E.G.; Jun, I.; Wray, J.; Madsen, M.B.; et al. Transient liquid water and water activity at Gale crater on Mars. Nat. Geosci. 2015, 8, 357–361. [Google Scholar] [CrossRef]
- Cabrol, N.A.; Wynn-Williams, D.D.; Crawford, D.A.; Grin, E.A. Recent aqueous environments in Martian impact craters: An astrobiological perspective. Icarus 2001, 154, 98–112. [Google Scholar] [CrossRef]
- Schwenzer, S.P.; Abramov, O.; Allen, C.C.; Bridges, J.C.; Clifford, S.; Filiberto, J.; Kring, D.A.; Lasue, J.; McGovern, P.J.; Newsom, H.E.; et al. Gale crater: Formation and post-impact hydrous environments. Planet. Space Sci. 2012, 70, 84–95. [Google Scholar] [CrossRef]
- Thomson, B.J.; Bridges, N.T.; Milliken, R.; Baldrige, A.; Hook, S.J.; Crowley, J.K.; Marion, G.M.; de Souza Filho, C.R.; Brwon, A.J.; Weitz, C.M. Constraints on the origin and evolution of the layered mound in Gale crater, Mars using Mars Reconaissance Orbiter data. Icarus 2011, 214, 413–432. [Google Scholar] [CrossRef]
- Anderson, R.B.; Bell, J.F., III. Geologic mapping and characterization of Gale crater and implications for its potential as a Mars Science Laboratory landing site. Int. J. Mars Sci. Explor. 2005, 4, 76–128. [Google Scholar] [CrossRef]
- Vaniman, D.T.; Bish, D.L.; Ming, D.W.; Bristow, T.F.; Morris, R.V.; Blake, D.F.; Chipera, S.J.; Morrison, S.M.; Treiman, A.H.; Rampe, E.B.; et al. Mineralogy of a mudstone at Yellowknife Bay, Gale crater, Mars. Science 2014, 343, 1243480. [Google Scholar] [CrossRef] [PubMed]
- Bridges, J.C.; Schwenzer, S.P.; Leveille, R.; Westall, F.; Wiens, R.C.; Mangold, N.; Bristow, T.; Edwards, P.; Berger, G. Diagenesis and clay mineral formation at Gale crater, Mars. J. Geophys. Res. Planets 2015, 120, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Leshin, L.A.; Mahaffy, P.R.; Webster, C.R.; Cabane, M.; Coll, P.; Conrad, P.G.; Archer, P.D., Jr.; Atreya, S.K.; Brunner, A.E.; Buch, A.; et al. Volatile, isotope, and organic analysis of martian fines with the Mars Curiosity rover. Science 2013, 341, 1238937. [Google Scholar] [CrossRef] [PubMed]
- Sutter, B.; Mcadam, A.C.; Mahaffy, P.R.; Ming, D.W.; Edgett, K.S.; Rampe, E.B.; Eigenbrode, J.L.; Franz, H.B.; Freissinet, C.; Grotzinger, J.P.; et al. Evolved gas analyses of sedimentary rocks and eolian sediment in Gale crater, Mars: Results of the Curiosity rover’s sample analysis at Mars (SAM) instrument from Yellowknife Bay to the Namib Dune. J. Geophys. Res. Planets 2017, 122, 2574–2609. [Google Scholar] [CrossRef]
- Hurowitz, J.A.; Grotzinger, J.P.; Fischer, W.W.; Mclennan, S.M.; Milliken, R.E.; Stein, N.; Vasavada, A.R.; Dehouck, E.; Eigenbrode, J.L.; Fairén, A.G.; et al. Redox stratification of an ancient lake in Gale crater, Mars. Science 2017, 356, eaah6849. [Google Scholar] [CrossRef] [PubMed]
- Stein, N.; Grotzinger, J.P.; Schieber, J.; Mangold, N.; Newsom, H.; Minitti, M.; Sumner, D.; Edgett, K.S.; Stack, K.; Fedo, C.; et al. Candidate desiccation cracks in the upper murray formation, Gale crater, Mars [abstract 2387]. In 48th Lunar Planetary Science Conference Abstracts; Lunar and Planetary Institute: Houston, TX, USA, 2017. [Google Scholar]
- Yen, A.S.; Ming, D.W.; Vaniman, D.T.; Gellert, R.; Blake, D.F.; Morris, R.V.; Morrison, S.M.; Bristow, T.F.; Chipera, S.J.; Edgett, K.S.; et al. Multiple stages of aqueous alteration along fractures in mudstone and sandstone strata in Gale crater, Mars. Earth Planet. Sci. Lett. 2017, 471, 186–198. [Google Scholar] [CrossRef]
- Eigenbrode, J.L.; Summons, R.E.; Steele, A.; Freissinet, C.; Millan, M.; Navarro-González, R.; Sutter, B.; McAdam, A.C.; Franz, A.; Glavin, D.P.; et al. Organic matter preserved in 3-billion-year-old mudstones at Gale crater, Mars. Science 2018, 360, 1096–1101. [Google Scholar] [CrossRef] [PubMed]
- Banham, S.G.; Gupta, S.; Rubin, D.M.; Watkins, J.A.; Sumner, D.Y.; Edgett, K.S.; Gortzinger, J.P.; Lewis, K.W.; Edgar, L.A.; Stack-Morgan, K.M.; et al. Ancient Martian aeolian processes and palaeomorphology reconstruction from the Stimson formation on the lower slope of Aeolis Mons, Gale crater, Mars. Sedimentology 2018, 65, 993–1042. [Google Scholar] [CrossRef]
- Rampe, E.B.; Ming, D.W.; Blake, D.F.; Bristow, T.F.; Chipera, S.J.; Grotzinger, J.P.; Morris, R.V.; Morrison, S.M.; Vaniman, D.T.; Yen, A.S.; et al. Mineralogy of an ancient lacustrine mudstone succession from the Murray formation, Gale crater, Mars. Earth Planet. Sci. Lett. 2017, 471, 172–185. [Google Scholar] [CrossRef]
- Morris, R.V.; Vaniman, D.T.; Blake, D.F.; Gellert, R.; Chipera, S.J.; Rampe, E.B.; Ming, D.W.; Morrison, S.M.; Downs, R.T.; Treiman, A.H.; et al. High-temperature, possibly silicic, Volcanism on Mars evidenced by tridymite detection in high-SiO2 sedimentary rock at Gale crater. Proc. Natl. Acad. Sci. USA 2016, 113, 7071–7076. [Google Scholar] [CrossRef] [PubMed]
- Frydenvang, J.; Gasda, P.J.; Hurowitz, J.A.; Grotzinger, J.P.; Wiens, R.C.; Newsom, H.E.; Edgett, K.S.; Watkins, J.; Bridges, J.C.; Maurice, S.; et al. Diagenetic silica enrichment and late-stage groundwater activity in Gale crater, Mars. Geophys. Res. Lett. 2017, 44, 4716–4724. [Google Scholar] [CrossRef]
- Schwenzer, S.P.; Bridges, J.C.; Wiens, R.C.; Conrad, P.G.; Kelley, S.P.; Leveille, R.; Mangold, N.; Martin-Torres, J.; Mcadam, A.; Newsom, H.; et al. Fluids during diagenesis and sulfate vein formation in sediments at Gale crater, Mars. Meteorit. Planet. Sci. 2016, 51, 2175–2202. [Google Scholar] [CrossRef]
- Horvath, D.G.; Andrews-Hanna, J.C. Reconstructing the past climate at Gale crater, Mars, from hydrological modeling of late-stage lakes. Geophys. Res. Lett. 2017, 44, 8196–8204. [Google Scholar] [CrossRef]
- Stein, N.; Grotzinger, J.P.; Schieber, J.; Mangold, N.; Hallet, B.; Newsom, H.; Stack, K.; Berger, J.A.; Thompson, L.; Siebach, K.L.; et al. Desiccation cracks provide evidence of lake drying on Mars, Sutton Island member, Murray formation, Gale crater. Geology 2018, 46, 515–518. [Google Scholar] [CrossRef]
- Hausrath, E.M.; Ming, D.W.; Peretyazhko, T.S.; Rampe, E.B. Reactive transport and mass balance modeling of the Stimson sedimentary formation and altered fracture zones constrain diagenetic conditions at Gale crater, Mars. Earth Planet. Sci. Lett. 2018, 491, 1–10. [Google Scholar] [CrossRef]
- Palucis, M.C.; Dietrich, W.E.; Williams, R.M.E.; Hayes, A.G.; Parker, T.; Sumner, D.Y.; Mangold, N.; Lewis, K.; Newsom, H. Sequence and relative timing of large lakes in Gale crater (Mars) after the formation of Mount Sharp. J. Geophys. Res. Planets 2016, 121, 472–496. [Google Scholar] [CrossRef]
- Ehlmann, B.L.; Mustard, J.F.; Murchie, S.L.; Poulet, F.; Bishop, J.L.; Brown, A.J.; Calvin, W.M.; Clark, R.N.; Des Marais, D.J.; Milliken, R.E.; et al. Orbital identification of carbonate-bearing rocks on Mars. Science 2008, 322, 1828–1832. [Google Scholar] [CrossRef] [PubMed]
- Ansan, V.; Loizeau, D.; Mangold, N.; Le Mouelic, S.; Carter, J.; Poulet, F.; Dromart, G.; Lucas, A.; Bibring, J.P.; Gendrin, A.; et al. Stratigraphy, mineralogy, and origin of layered deposits inside Terby crater, Mars. Icarus 2011, 211, 273–304. [Google Scholar] [CrossRef]
- Schon, S.C.; Head, J.W.; Fassett, C.I. An overfilled lacustrine system and progradational delta in Jezero crater, Mars: Implications for Noachian climate. Planet. Space Sci. 2012, 67, 28–45. [Google Scholar] [CrossRef]
- McLennan, S.M.; Bell, J.F.; Calvin, W.M.; Christensen, P.R.; Clark, B.C.; De Souza, P.A.; Farmer, J.; Farrand, W.H.; Fike, D.A.; Gellert, R.; et al. Provenance and diagenesis of the evaporite-bearing Burns formation, Meridiani Planum, Mars. Earth Planet. Sci. Lett. 2005, 240, 95–121. [Google Scholar] [CrossRef]
- Ming, D.W.; Mittlefehldt, D.W.; Morris, R.V.; Golden, D.C.; Gellert, R.; Yen, A.; Clark, B.C.; Squyres, S.W.; Farrand, W.H.; Ruff, S.W.; et al. Geochemical and mineralogical indicators for aqueous processes in the Columbia Hills of Gusev crater, Mars. J. Geophys. Res. Planets 2006, 111. [Google Scholar] [CrossRef]
- Zolotov, M.Y.; Mironenko, M.V. Timing of acid weathering on Mars: A kinetic-thermodynamic assessment. J. Geophys. Res. Planets 2007, 112. [Google Scholar] [CrossRef]
- McAdam, A.C.; Zolotov, M.Y.; Sharp, T.G.; Leshin, L.A. Preferential low-pH dissolution of pyroxene in plagioclase-pyroxene mixtures: Implications for martian surface materials. Icarus 2008, 196, 90–96. [Google Scholar] [CrossRef]
- Bridges, J.C.; Loizeau, D.; Sefton-Nash, E.; Vago, J.; Williams, R.M.E.; Balme, M.; Turner, S.M.R.; Fawdon, P.; Davis, J.M. Selction and characterisation of the ExoMars 2020 rover landing sites [abstract 2378]. In 48th Lunar Planetary Science Conference Abstracts; Lunar and Planetary Institute: Houston, TX, USA, 2017. [Google Scholar]
- Golombek, M.P.; Otero, R.E.; Heverly, M.C.; Ono, M.; Willifor, K.H.; Rothrock, B.; Milkovich, S.; Almeida, E.; Calef, F.; Williams, N.; et al. Characterization of Mars rover 2020 prospective landing sites leading up to the second down selection [abstract 2333]. In Lunar and Planetary Sciences Conference; Lunar and Planerary Institute: Houston, TX, USA, 2017. [Google Scholar]
- Carr, M.H.; Head, J.W., III. Geologic history of Mars. Earth Planet. Sci. Lett. 2010, 284, 185–203. [Google Scholar] [CrossRef]
- Landis, G.A. Martian Water: Are there extant Halobacteria on Mars? Astrobiology 2001, 1, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Remolar, D.; Gomez-Elvira, J.; Gomez, F.; Sebastian, E.; Martiin, J.; Manfredi, J.A.; Torres, J.; Kesler, C.G.; Amils, R. The Tinto River, an extreme acidic environment under control of iron, as an analog of the Terra Meridiani hematite site of Mars. Planet. Space Sci. 2004, 52, 239–248. [Google Scholar] [CrossRef]
- Amils, R.; Gonzalez-Toril, E.; Fernandez-Remolar, D.; Gomez, F.; Aguilera, A.; Rodriguez, N.; Malki, M.; Garcia-Moyano, A.; Fairen, A.G.; De La Fuente, V.; et al. Extreme environments as Mars terrestrial analogs: The Rio Tinto case. Planet. Space Sci. 2007, 55, 370–381. [Google Scholar] [CrossRef]
- Direito, S.O.L.; Ehrenfreund, P.; Marees, A.; Staats, M.; Foing, B.; Roling, W.F.M. A wide variety of putative extremophiles and large beta-diversity at the Mars Desert Research Station (Utah). Int. J. Astrobiol. 2011, 10, 191–207. [Google Scholar] [CrossRef]
- Fox-Powell, M.G.; Hallsworth, J.E.; Cousins, C.R.; Cockell, C.S. Ionic strength is a barrier to the habitability of Mars. Astrobiology 2016, 16, 427–442. [Google Scholar] [CrossRef] [PubMed]
- Klingelhöfer, G.; Morris, R.V.; Bernhardt, B.; Schröder, C.; Rodionov, D.S.; De Souza, P.A.; Yen, A.; Gellert, R.; Evlanov, E.N.; Zubkov, B.; et al. Jarosite and hematite at Meridiani Planum from Opportunity’s Mossbauer spectrometer. Science 2004, 306, 1740–1745. [Google Scholar] [CrossRef] [PubMed]
- Aubrey, A.; Cleaves, H.J.; Chalmers, J.H.; Skelley, A.M.; Mathies, R.A.; Grunthaner, F.J.; Ehrenfreund, P.; Bada, J.L. Sulfate minerals and organic compounds on Mars. Geology 2006, 34, 357–360. [Google Scholar] [CrossRef]
- Cousins, C. Volcanogenic fluvial-lacustrine environments in Iceland and their utility for identifying past habitability on Mars. Life 2015, 5, 568–586. [Google Scholar] [CrossRef] [PubMed]
- Marshall, A.O.; Cestari, N.A. Biomarker analysis of samples visually identified as microbial in the Eocene Green River Formation: An analogue for Mars. Astrobiology 2015, 15, 770–775. [Google Scholar] [CrossRef] [PubMed]
- Treiman, A.H.; Morris, R.V.; Agresti, D.G.; Graff, T.G.; Achilles, C.N.; Rampe, E.B.; Bristow, T.F.; Ming, D.W.; Blake, D.F.; Vaniman, D.T.; et al. Ferrian saponite from the Santa Monica Mountains (California, USA, Earth): Characterization as an analog for clay minerals on Mars with application to Yellowknife Bay in Gale crater. Am. Mineral. 2014, 99, 2234–2250. [Google Scholar] [CrossRef]
- Young, B.W.; Chan, M.A. Gypsum veins in Triassic Moenkopi mudrocks of southern Utah: Analogs to calcium sulfate veins on Mars. J. Geophys. Res. Planets 2017, 122, 150–171. [Google Scholar] [CrossRef]
- Telesh, I.V.; Khlebovich, V.V. Principal processes within the estuarine salinity gradient: A review. Mar. Pollut. Bull. 2010, 61, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Arvidson, R.E.; Squyres, S.W.; Bell, J.F.; Catalano, J.G.; Clark, B.C.; Crumpler, L.S.; De Souza, P.A.; Fairen, A.G.; Farrand, W.H.; Fox, V.K.; et al. Ancient aqueous environments at Endeavour Crater, Mars. Science 2014, 343, 1248097. [Google Scholar] [CrossRef] [PubMed]
- Bristow, T.F.; Bish, D.L.; Vaniman, D.T.; Morris, R.V.; Blake, D.F.; Grotzinger, J.P.; Rampe, E.B.; Crisp, J.A.; Achilles, C.N.; Ming, D.W.; et al. The origin and implications of clay minerals from Yellowknife Bay, Gale crater, Mars. Am. Mineral. 2015, 100, 824–836. [Google Scholar] [CrossRef] [PubMed]
- Ch2M Hill. Dee Estuary. North West Estuaries Processes Reports; Ch2M Hill: Englewood, CO, USA, 2013; Available online: https://www.liverpool.ac.uk/~cmi/dee/Dee_rep2013.pdf (accessed on 15 September 2017).
- Moore, R.D.; Wolf, J.; Souza, A.J.; Flint, S.S. Morphological evolution of the Dee estuary, eastern Irish Sea, UK: A tidal asymmetry approach. Geomorphology 2009, 103, 588–596. [Google Scholar] [CrossRef]
- Google Maps. River Dee Estuary. 2016. Available online: https://www.google.com/maps (accessed on 2 July 2017).
- Summers, S.; Whiteley, A.S.; Kelly, L.C.; Cockell, C.S. Land coverage influences the bacterial community composition in the critical zone of a sub-Arctic basaltic environment. FEMS Microbiol. Ecol. 2013, 86, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Herlemann, D.P.R.; Labrenz, M.; Jurgens, K.; Bertilsson, S.; Waniek, J.J.; Andersson, A.F. Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 2011, 5, 1571–1579. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, J.R.; Sato, T.; Weightman, A.J.; Martin, T.A.; Fry, J.C.; Hiom, S.J.; Dymock, D.; Wade, W.G. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Appl. Environ. Microbiol. 1998, 64, 795–799. [Google Scholar] [PubMed]
- Baker, G.C.; Smith, J.J.; Cowan, D.A. Review and re-analysis of domain-specific 16S primers. J. Microbiol. Methods 2003, 55, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.G.; Peplies, J.; Glockner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2010; Available online: http://www.R-project.org (accessed on 1 June 2015).
- Blackwood, C.B.; Hudleston, D.; Zak, D.; Buyer, J.S. Inrerpreating ecological diversity indices applied to terminal restriction fragment length polymorphism data: Insights from simulated microbial communities. Appl. Environ. Microbiol. 2007, 73, 5276–5283. [Google Scholar] [CrossRef] [PubMed]
- Clarke, K.R.; Warwick, R.M. A further biodiversity index applicable to species lists: Variation in taxonomic distinctness. Mar. Ecol. Prog. Ser. 2001, 216, 265–278. [Google Scholar] [CrossRef]
- Squyres, S.W.; Arvidson, R.E.; Bollen, D.; Bell, J.F.; Brückner, J.; Cabrol, N.A.; Calvin, W.M.; Carr, M.H.; Christensen, P.R.; Clark, B.C.; et al. Overview of the Opportunity Mars Exploration rover mission to Meridiani Planum: Eagle Crater to Purgatory Ripple. J. Geophys. Res. E Planets 2006, 111, 1–19. [Google Scholar] [CrossRef]
- Greeley, R.; Arvidson, R.; Bell, J.F.; Christensen, P.; Foley, D.; Haldemann, A.; Kuzmin, R.O.; Landis, G.; Neakrase, L.D.V.; Neukum, G.; et al. Martian variable features: New insight from the Mars express orbiter and the Mars Exploration Rover Spirit. J. Geophys. Res. E Planets 2005, 110, 1–7. [Google Scholar] [CrossRef]
- McCollom, T.M.; Hynek, B.M. A volcanic environment for bedrock diagenesis at Meridiani Planum on Mars. Nature 2005, 438, 1129–1131. [Google Scholar] [CrossRef] [PubMed]
- Wray, J.J.; Murchie, S.L.; Squyres, S.W.; Seelos, F.P.; Tomabene, L.L. Diverse aqueous environments on ancient Mars revealed in the southern highlands. Geology 2009, 37, 1043–1046. [Google Scholar] [CrossRef]
- Phillips, R.J.; Zuber, M.T.; Solomon, S.C.; Golombek, M.P.; Jakosky, B.M.; Banerdt, W.B.; Smith, D.E.; Williams, R.M.E.; Hynek, B.M.; Aharonson, O.; et al. Ancient geodynamics and global-scale hydrology on Mars. Science 2001, 291, 2587–2591. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.S.; Wiens, R.C.; Vaniman, D.T.; Beegle, L.; Gasnault, O.; Newsom, H.E.; Maurice, S.; Meslin, P.Y.; Clegg, S.; Cousin, A.; et al. ChemCam investigation of the John Klein and Cumberland drill holes and tailings, Gale crater, Mars. Icarus 2016, 277, 330–341. [Google Scholar] [CrossRef]
- Sutter, B.; Eigenbrode, J.L.; Steele, A.; Ming, D.W. The Sample at Mars Analysis (SAM) detections of CO2 and CO in sedimentary material from Gale crater, Mars: Implications for the presence of organic carbon and microbial habitability on Mars. In American Geophysical Union, Fall General Assembly Abstract; American Geophysical Union: Washington, DC, USA, 2016. [Google Scholar]
- Freissinet, C.; Glavin, D.P.; Mahaffy, P.R.; Miller, K.E.; Eigenbrode, J.L.; Summons, R.E.; Brunner, A.E.; Buch, A.; Szopa, C.; Archer, P.D.; et al. Organic molecules in the Sheepbed Mudstone, Gale crater, Mars. J. Geophys. Res. Planets 2015, 120, 495–514. [Google Scholar] [CrossRef] [PubMed]
- Ming, D.W.; Archer, P.D.; Glavin, D.P.; Eigenbrode, J.L.; Franz, H.B.; Sutter, B.; Brunner, A.E.; Stern, J.C.; Freissinet, C.; Mcadam, A.C.; et al. Volatile and organic compositions of sedimentary rocks in Yellowknife Bay, Gale crater, Mars. Science 2014, 343, 1245267. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.E.; Eigenbrode, J.L.; Freissinet, C.; Glavin, D.P.; Kotrc, B.; Francois, P.; Summons, R.E. Potential precursor compounds for chlorohydrocarbons detected in Gale crater, Mars, by the SAM instrument suite on the Curiosity Rover. J. Geophys. Res. Lett. Planets 2016, 121, 296–308. [Google Scholar] [CrossRef]
- Sephton, M.A.; Wright, I.P.; Gilmour, I.; de Leeuw, J.W.; Grady, M.M.; Pillinger, C.T. High molecular weight organic matter in martian meteorites. Planet. Space Sci. 2002, 50, 711–716. [Google Scholar] [CrossRef]
- Steele, A.; Mccubbin, F.M.; Fries, M.; Kater, L.; Boctor, N.Z.; Fogel, M.L.; Conrad, P.G.; Glamoclija, M.; Spencer, M.; Morrow, A.L.; et al. A reduced organic carbon component in Martian basalts. Science 2012, 337, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Wackett, L.P.; Dodge, A.G.; Ellis, L.B.M. Microbial genomics and the periodic table. Appl. Environ. Microbiol. 2004, 70, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Pontefract, A.; Osinski, G.R.; Lindgren, P.; Parnell, J.; Cockell, C.S. The effects of meteorite impacts on the availability of bioessential elements for endolithic organisms. Meteorit. Planet. Sci. 2012, 47, 1681–1691. [Google Scholar] [CrossRef]
- Vandevivere, P.; Welch, S.A.; Ullman, W.J.; Kirchman, D.L. Enhanced dissoluton of silicate minerals by bacteria at near-neutral pH. Microb. Ecol. 1994, 27, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Forni, O.; Gaft, M.; Toplis, K.J.; Clegg, S.M.; Maurice, S.; Wiens, R.C.; Mangold, N.; Gasnault, O.; Sautter, V.; Le Mouélic, S.; et al. First detection of fluorine on Mars: Implications for Gale crater’s geochemistry. Geohpys. Res. Lett. 2015, 42, 1020–1028. [Google Scholar] [CrossRef]
- Blank, J.G.; Ollila, A.M.; Lanza, N.L.; Forni, O.; Mangold, N.; Nachon, M.; Clegg, S.M.; Yen, A.; Maurice, S.; Wiens, R.C. Detection of phosphorous by ChemCam in Gale crater [abstract 2850]. In 46th Lunar and Planetary Science Conference Abstracts; Lunar and Planetary Institute: Houston, TX, USA, 2015. [Google Scholar]
- Lozupone, C.A.; Knight, R. Global patterns in bacterial diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 11436–11440. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Yoshizawa, S.; Inomata, K.; Kogure, K.; Yokota, A. Halioglobus japonicus gen. nov., sp. nov. and Halioglobus pacificus sp. nov., members of the class Gamma proteobacteria isolated from seawater. Int. J. Syst. Evol. Microbiol. 2012, 62, 1784–1789. [Google Scholar] [CrossRef] [PubMed]
- Sekiguchi, Y.; Yamada, T.; Hanada, S.; Ohashi, A.; Harada, H.; Kamagata, Y. Anaerolinea thermophila gen. nov., sp. nov. and Caldilinea aerophila gen. nov., sp. nov., novel filamentous thermophiles that represent a previously uncultured lineage of the domain Bacteria at the subphylum level. Int. J. Syst. Evol. Microbiol. 2003, 53, 1843–1851. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, P.; Bohli, M.; Cayol, J.L.; Joseph, M.; Guasco, S.; Dubourg, K.; Cambar, J.; Michotey, V.; Bonin, P.; Fardeau, M.L.; et al. Caldilinea tarbellica sp. nov., a filamentous, thermophilic, anaerobic bacterium isolated from a deep hot aquifer in the Aquitaine Basin. Int. J. Syst. Evol. Microbiol. 2011, 61, 1436–1441. [Google Scholar] [CrossRef] [PubMed]
- Kale, V.; Bjornsdottir, S.H.; Friodjonsson, O.H.; Petursdottir, S.K.; Omarsdottir, S.; Hreggviosson, G.O. Litorilinea aerophila gen. nov., sp. nov., an aerobic member of the class Caldilineae, phylum Chloroflexi, isolated from an intertidal hot spring. Int. J. Syst. Evol. Microbiol. 2013, 63, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Ng, H.J.; Lopez-Perez, M.; Webb, H.K.; Gomez, D.; Sawabe, T.; Ryan, J.; Vyssotski, M.; Bizet, C.; Malherbe, F.; Mikhailov, V.V.; et al. Marinobacter salarius sp. nov. and Marinobacter similis sp. nov., isolated from sea water. PLoS ONE 2014, 9, e106514. [Google Scholar] [CrossRef] [PubMed]
- Konneke, M.; Bernhard, A.E.; De La Torre, J.R.; Walker, C.B.; Waterbury, J.B.; Stahl, D.A. Isolation of an autotrophic ammonia-oxidizing marine Archaeon. Nature 2005, 437, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Cockell, C.S. Trajectories of martian habitability. Astrobiology 2014, 14, 182–203. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Sekiguchi, Y.; Hanada, S.; Imachi, H.; Ohashii, A.; Harada, H.; Kamagata, Y. Anaerolinea thermolimosa sp. nov., Levilinea saccharolytica gen. nov., sp. nov. and Leptolinea tardivitalis gen. nov., sp. nov., novel filamentous anaerobes, and description of the new classes Anaerolineae classis nov. and Caldilineae classis nov. in the bacterial phylum Chloroflexi. Int. J. Syst. Evol. Microbiol. 2006, 56, 1331–1340. [Google Scholar] [PubMed]
- Zhang, B.; Xu, X.; Zhu, L. Structural and functional of the microbial consortia of activated sludge in typical municipal wastewater treatment plants in winter. Sci. Rep. 2017, 7, 17930. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Temperature (°C) | pH | TOC (%) |
|---|---|---|---|
| 1 | 12.2 | 8.23 | 1.50 |
| 2 | 12.2 | 8.14 | 1.46 |
| 3 | 11.6 | 8.08 | 1.49 |
| 4 | 11.8 | 8.45 | 1.47 |
| 5 | 12.3 | 8.31 | 1.10 |
| 6 | 13.3 | 8.38 | 1.84 |
| 7 | 11.9 | 8.29 | 1.41 |
| 8 | 12.3 | 8.08 | 0.99 |
| ANOVA test | |||
| p value | >0.1 | >0.1 | >0.1 |
| Isolate | Genebank No° | Closest Genebank Relative | Sequence Identity | Class |
|---|---|---|---|---|
| E01 | MH450108 | Acinetobacter johnsonii | 99% | Gammaproteobacteria |
| E02 | MH450105 | Clostridium amygdalinum | 98% | Clostridia |
| E03 | MH450106 | Bacillus toyonensis | 99% | Bacilli |
| John Klein | Cumberland | Lubango | Buckskin | River Dee | |
|---|---|---|---|---|---|
| Plagioclase | 44.8 | 41 | 43.2 | 42.8 | |
| Fe-forsterite | 5.7 | 1.9 | |||
| Augite | 7.6 | 9 | |||
| Pigeonite | 11.3 | 16 | 5.9 | ||
| Orthopyroxene | 6.1 | 9 | 10.4 | ||
| Magnetite | 7.6 | 9 | 11.1 | 6.9 | |
| Anhydrite | 5.3 | 12.3 | 1.8 | ||
| Bassanite | 2.1 | 1.2 | 9.0 | ||
| Quartz/SiO2-phase § | 0.9 | 0.2 | 3.5 | 40.1 | 63 |
| Sanidine | 2.4 | 3.5 | 8.4 | 9 | |
| Haematite † | 1.2 | 1.3 | 2.3 | 5 * | |
| Ilmenite | 1.2 | ||||
| Akaganeite | 2.3 | 3 | |||
| Halite | 0.3 | 0.3 | |||
| Pyrite | 0.6 | ||||
| Pyrrhotite | 2 | 1.9 | |||
| Amorphous $ | 28 | 31 | 73 | 60 | |
| Clay $ | 22 | 18 | 23 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curtis-Harper, E.; Pearson, V.K.; Summers, S.; Bridges, J.C.; Schwenzer, S.P.; Olsson-Francis, K. The Microbial Community of a Terrestrial Anoxic Inter-Tidal Zone: A Model for Laboratory-Based Studies of Potentially Habitable Ancient Lacustrine Systems on Mars. Microorganisms 2018, 6, 61. https://doi.org/10.3390/microorganisms6030061
Curtis-Harper E, Pearson VK, Summers S, Bridges JC, Schwenzer SP, Olsson-Francis K. The Microbial Community of a Terrestrial Anoxic Inter-Tidal Zone: A Model for Laboratory-Based Studies of Potentially Habitable Ancient Lacustrine Systems on Mars. Microorganisms. 2018; 6(3):61. https://doi.org/10.3390/microorganisms6030061
Chicago/Turabian StyleCurtis-Harper, Elliot, Victoria K. Pearson, Stephen Summers, John C. Bridges, Susanne P. Schwenzer, and Karen Olsson-Francis. 2018. "The Microbial Community of a Terrestrial Anoxic Inter-Tidal Zone: A Model for Laboratory-Based Studies of Potentially Habitable Ancient Lacustrine Systems on Mars" Microorganisms 6, no. 3: 61. https://doi.org/10.3390/microorganisms6030061
APA StyleCurtis-Harper, E., Pearson, V. K., Summers, S., Bridges, J. C., Schwenzer, S. P., & Olsson-Francis, K. (2018). The Microbial Community of a Terrestrial Anoxic Inter-Tidal Zone: A Model for Laboratory-Based Studies of Potentially Habitable Ancient Lacustrine Systems on Mars. Microorganisms, 6(3), 61. https://doi.org/10.3390/microorganisms6030061