Bacteriophages as New Human Viral Pathogens


Abstract
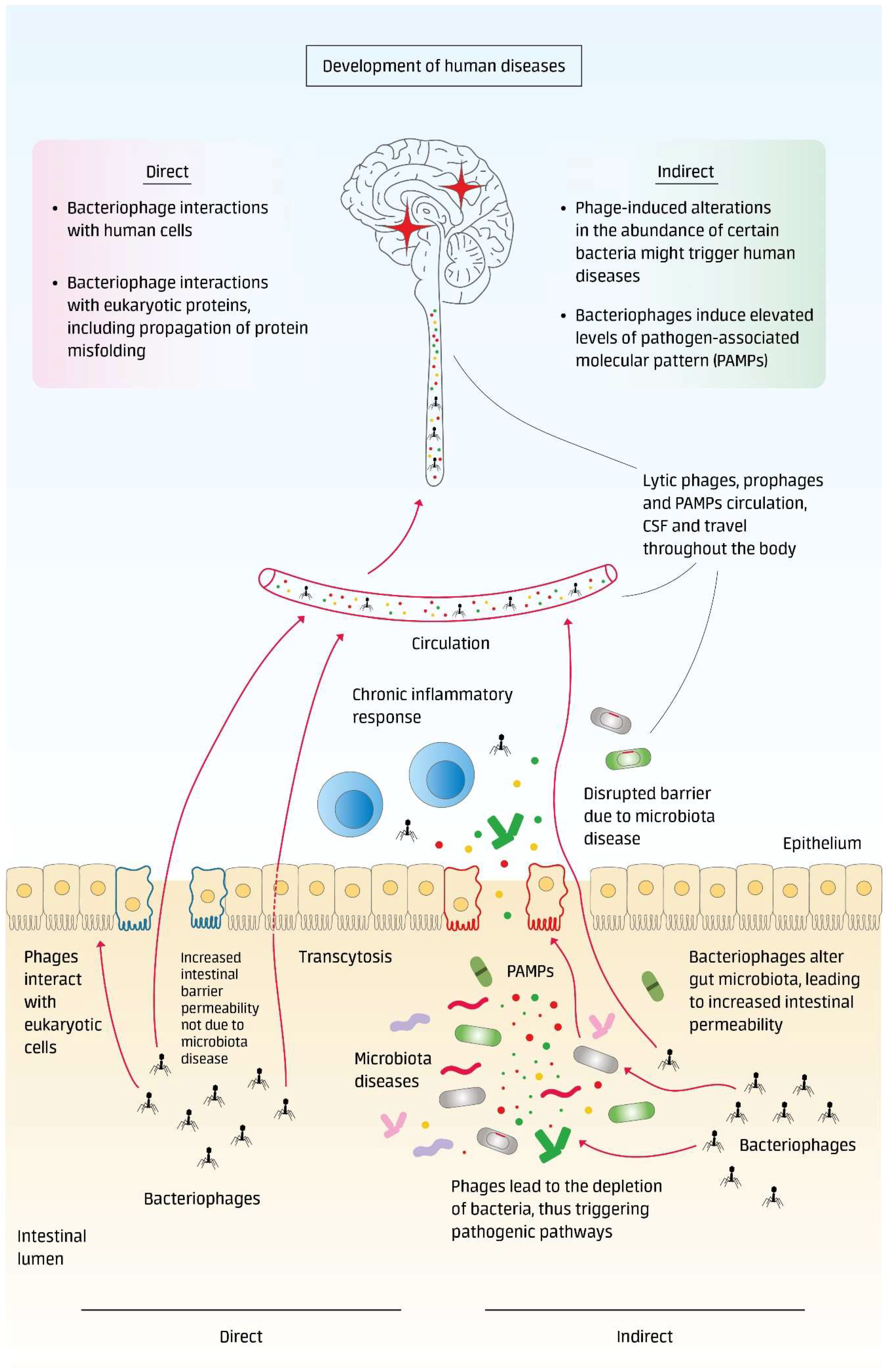
1. Introduction
- Direct interaction of phages with the host macroorganism (eukaryotic cells and proteins);
- Indirect interaction with the host macroorganism by causing harmful microbiota alterations and microbiota-related diseases (Figure 1).
1.1. Phages as Human Pathogens: Direct Actions of Phages
1.1.1. Model 1. Bacteriophage Interactions with Human Cells
1.1.2. Bacteriophage Interactions with Eukaryotic Proteins, Including Propagation of Protein Misfolding
1.2. Phages as Human Pathogens: Indirect Interactions through Microbiota Alterations and Microbiota Diseases
1.2.1. Phage-Induced Alterations in the Abundances of Certain Bacteria Might Trigger Human Diseases
1.2.2. Bacteriophages Induce Elevated Levels of Pathogen-Associated Molecular Pattern (PAMPs)
2. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clemente, J.; Ursell, L.; Parfrey, L.; Knight, R. The Impact of the Gut Microbiota on Human Health: An Integrative View. Cell 2012, 148, 1258–1270. [Google Scholar] [CrossRef] [PubMed]
- Heijtz, R.; Wang, S.; Anuar, F.; Qian, Y.; Bjorkholm, B.; Samuelsson, A.; Hibberd, M.; Forssberg, H.; Pettersson, S. Normal gut microbiota modulates brain development and behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 3047–3052. [Google Scholar] [CrossRef] [PubMed]
- Sommer, F.; Bäckhed, F. The gut microbiota—Masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Nauta, A.; Ben Amor, K.; Knippels, L.; Knol, J.; Garssen, J. Early life: Gut microbiota and immune development in infancy. Benef. Microbes 2010, 1, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.; Dinan, T. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.; Murphy, K.; Stanton, C.; Ross, R.; Kober, O.; Juge, N.; Avershina, E.; Rudi, K.; Narbad, A.; Jenmalm, M.; et al. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb. Ecol. Health Dis. 2015, 26, 26050. [Google Scholar] [CrossRef] [PubMed]
- Funkhouser, L.; Bordenstein, S. Mom Knows Best: The Universality of Maternal Microbial Transmission. PLoS Biol. 2013, 11, e1001631. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Bello, M.; Costello, E.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [PubMed]
- Faith, J.; Guruge, J.; Charbonneau, M.; Subramanian, S.; Seedorf, H.; Goodman1, A.; Clemente, J.; Knight, R.; Heath, A.; Leibel, R.; et al. The Long-Term Stability of the Human Gut Microbiota. Science 2013, 341, 1237439. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.; Rho, M.; Song, Y.; Lee, K.; Sung, J.; Ko, G. Stability of Gut Enterotypes in Korean Monozygotic Twins and Their Association with Biomarkers and Diet. Sci. Rep. 2014, 4, 7348. [Google Scholar] [CrossRef] [PubMed]
- Dalmasso, M.; Hill, C.; Ross, R. Exploiting gut bacteriophages for human health. Trends Microbiol. 2014, 22, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Hobbs, Z.; Abedon, S. Diversity of phage infection types and associated terminology: The problem with ‘Lytic or lysogenic’. FEMS Microbiol. Lett. 2016, 363, fnw047. [Google Scholar] [CrossRef] [PubMed]
- Clokie, M.; Millard, A.; Letarov, A.; Heaphy, S. Phages in nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, S.; Rashel, M.; Uchiyama, J.; Sakurai, S.; Ujihara, T.; Kuroda, M.; Imai, S.; Ikeuchi, M.; Tani, T.; Fujieda, M.; et al. Bacteriophage therapy: A revitalized therapy against bacterial infectious diseases. J. Infect. Chemother. 2005, 11, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.; Abedon, S.; Loc-Carrillo, C. Phage cocktails and the future of phage therapy. Future Microbiol. 2013, 8, 769–783. [Google Scholar] [CrossRef] [PubMed]
- Echols, H. Developmental Pathways for the Temperate Phage: Lysis VS Lysogeny. Annu. Rev. Genet. 1972, 6, 157–190. [Google Scholar] [CrossRef] [PubMed]
- Ptashne, M. Principles of a switch. Nat. Chem. Biol. 2011, 7, 484–487. [Google Scholar] [CrossRef] [PubMed]
- Payet, J.; Suttle, C. To kill or not to kill: The balance between lytic and lysogenic viral infection is driven by trophic status. Limnol. Oceanogr. 2013, 58, 465–474. [Google Scholar] [CrossRef]
- Tetz, G.; Tetz, V. Bacteriophage infections of microbiota can lead to leaky gut in an experimental rodent model. Gut Pathog. 2016, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Tetz, G.; Ruggles, K.; Zhou, H.; Heguy, A.; Tsirigos, A.; Tetz, V. Bacteriophages as potential new mammalian pathogens. Sci. Rep. 2017, 7, 7043. [Google Scholar] [CrossRef] [PubMed]
- Lepage, P.; Colombet, J.; Marteau, P.; Sime-Ngando, T.; Dore, J.; Leclerc, M. Dysbiosis in inflammatory bowel disease: A role for bacteriophages? Gut 2008, 57, 424–425. [Google Scholar] [CrossRef] [PubMed]
- Norman, J.; Handley, S.; Baldridge, M.; Droit, L.; Liu, C.; Keller, B.; Kambal, A.; Monaco, C.; Zhao, G.; Fleshner, P.; et al. Disease-Specific Alterations in the Enteric Virome in Inflammatory Bowel Disease. Cell 2015, 160, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Manrique, P.; Dills, M.; Young, M. The Human Gut Phage Community and Its Implications for Health and Disease. Viruses 2017, 9, 141. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Rodriguez, T.; Naidu, M.; Abeles, S.; Boehm, T.; Ly, M.; Pride, D. Transcriptome analysis of bacteriophage communities in periodontal health and disease. BMC Genom. 2015, 16, 549. [Google Scholar] [CrossRef] [PubMed]
- Ly, M.; Abeles, S.; Boehm, T.; Robles-Sikisaka, R.; Naidu, M.; Santiago-Rodriguez, T.; Pride, D. Altered Oral Viral Ecology in Association with Periodontal Disease. mBio 2014, 5, e01133-14. [Google Scholar] [CrossRef] [PubMed]
- Richmond, P. American Attitudes toward the Germ Theory of Disease (1860–1880). J. Hist. Med. Allied Sci. 1954, IX, 428–454. [Google Scholar] [CrossRef]
- Manolio, T.; Collins, F.; Cox, N.; Goldstein, D.; Hindorff, L.; Hunter, D.; McCarthy, M.; Ramos, E.; Cardon, L.; Chakravarti, A.; et al. Finding the missing heritability of complex diseases. Nature 2009, 461, 747–753. [Google Scholar] [CrossRef] [PubMed]
- McClellan, J.; King, M. Genetic Heterogeneity in Human Disease. Cell 2010, 141, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, K. Tobacco smoke: Involvement of reactive oxygen species and stable free radicals in mechanisms of oxidative damage, carcinogenesis and synergistic effects with other respirable particles. Int. J. Environ. Res. Public Health 2009, 6, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Sarasin, A. An overview of the mechanisms of mutagenesis and carcinogenesis. Mutat. Res. Rev. Mutat. Res. 2003, 544, 99–106. [Google Scholar] [CrossRef]
- Hernandez, L.; Blazer, D. Genes, Behavior, and the Social Environment; National Academies Press: Washington, DC, USA, 2006. [Google Scholar]
- Bulik-Sullivan, B.; Finucane, H.; Anttila, V.; Gusev, A.; Day, F.; Loh, P.; Duncan, L.; Perry, J.; Patterson, N.; Robinson, E.; et al. An atlas of genetic correlations across human diseases and traits. Nat. Genet. 2015, 47, 1236–1241. [Google Scholar] [CrossRef] [PubMed]
- Worboys, M. Spreading Germs; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Farley, J. Parasites and the Germ Theory of Disease. Hosp. Pract. 1992, 27, 175–196. [Google Scholar] [CrossRef]
- Łusiak-Szelachowska, M.; Weber-Dąbrowska, B.; Jończyk-Matysiak, E.; Wojciechowska, R.; Górski, A. Bacteriophages in the gastrointestinal tract and their implications. Gut Pathog. 2017, 9, 44. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.; Baker, K.; Padman, B.; Patwa, R.; Dunstan, R.; Weston, T.; Schlosser, K.; Bailey, B.; Lithgow, T.; Lazarou, M.; et al. Bacteriophage Transcytosis Provides a Mechanism to Cross Epithelial Cell Layers. mBio 2017, 8, e01874-17. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, L.; Harmon, W.; Strom, T.; Bunn, H. In utero detection of T7 phage after systemic administration to pregnant mice. Biotechniques 2004, 37, 81–83. [Google Scholar]
- Barr, J. A bacteriophages journey through the human body. Immunol. Rev. 2017, 279, 106–122. [Google Scholar] [CrossRef] [PubMed]
- Carlton, R.; Noordman, W.; Biswas, B.; de Meester, E.; Loessner, M. Bacteriophage P100 for control of Listeria monocytogenes in foods: Genome sequence, bioinformatic analyses, oral toxicity study, and application. Regul. Toxicol. Pharmacol. 2005, 43, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.; Viñas, I.; Colàs, P.; Anguera, M.; Usall, J.; Abadias, M. Effectiveness of a bacteriophage in reducing Listeria monocytogenes on fresh-cut fruits and fruit juices. Food Microbiol. 2014, 38, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Geier, M.; Trigg, M.; Merril, C. Fate of Bacteriophage Lambda in Non-immune Germ-free Mice. Nature 1973, 246, 221–223. [Google Scholar] [CrossRef] [PubMed]
- Merril, C.; Scholl, D.; Adhya, S. Timeline: The prospect for bacteriophage therapy in Western medicine. Nat. Rev. Drug Disc. 2003, 2, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Dabrowska, K.; Switala-Jelen, K.; Opolski, A.; Weber-Dabrowska, B.; Gorski, A. Bacteriophage penetration in vertebrates. J. Appl. Microbiol. 2005, 98, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Merril, C.; Biswas, B.; Carlton, R.; Jensen, N.; Creed, G.; Zullo, S.; Adhya, S. Long-circulating bacteriophage as antibacterial agents. Proc. Natl. Acad. Sci. USA 1996, 93, 3188–3192. [Google Scholar] [CrossRef] [PubMed]
- Gόrski, A.; Krotkiewski, H.; Zimecki, M. Inflammation; Springer: Dordrecht, The Netherlands, 2001. [Google Scholar]
- Çetin, E.; Kaya, S.; Demirci, M.; Aridogan, B. Comparison of the BACTEC blood culture system versus conventional methods for culture of normally sterile body fluids. Adv. Ther. 2007, 24, 1271–1277. [Google Scholar] [CrossRef] [PubMed]
- Radstrom, P.; Backman, A.; Qian, N.; Kragsbjerg, P.; Pallson, C.; Olcen, P. Detection of bacterial DNA in cerebrospinal fluid by an assay for simultaneous detection of Neisseria meningitidis, Haemophilus influenzae, and streptococci using a seminested PCR strategy. J. Clin. Microbiol. 1994, 32, 2738–2744. [Google Scholar] [PubMed]
- Schuurman, T.; de Boer, R.; Kooistra-Smid, A.; van Zwet, A. Prospective Study of Use of PCR Amplification and Sequencing of 16S Ribosomal DNA from Cerebrospinal Fluid for Diagnosis of Bacterial Meningitis in a Clinical Setting. J. Clin. Microbiol. 2004, 42, 734–740. [Google Scholar] [CrossRef] [PubMed]
- Ihekwaba, U.; Kudesia, G.; McKendrick, M. Clinical Features of Viral Meningitis in Adults: Significant Differences in Cerebrospinal Fluid Findings among Herpes Simplex Virus, Varicella Zoster Virus, and Enterovirus Infections. Clin. Infect. Dis. 2008, 47, 783–789. [Google Scholar] [CrossRef] [PubMed]
- Piacentini, R.; De Chiara, G.; Li Puma, D.; Ripoli, C.; Marcocci, M.; Garaci, E.; Palamara, A.; Grassi, C. HSV-1 and Alzheimer’s disease: More than a hypothesis. Front. Pharmacol. 2014, 5, 97. [Google Scholar] [CrossRef] [PubMed]
- Itzhaki, R.; Lathe, R.; Balin, B.; Ball, M.; Bearer, E.; Braak, H.; Bullido, M.; Carter, C.; Clerici, M.; Cosby, S.; et al. Microbes and Alzheimer’s Disease. J. Alzheimer’s Dis. 2016, 51, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Miranda-Saksena, M.; Saksena, N. Viruses and neurodegeneration. Virol. J. 2013, 10, 172. [Google Scholar] [CrossRef] [PubMed]
- McManus, R.; Heneka, M. Role of neuroinflammation in neurodegeneration: New insights. Alzheimer’s Res. Ther. 2017, 9, 14. [Google Scholar] [CrossRef] [PubMed]
- Fasano, A. Leaky Gut and Autoimmune Diseases. Clin. Rev. Allergy Immunol. 2011, 42, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Minagar, A.; Alexander, J. Blood-brain barrier disruption in multiple sclerosis. Mult. Scler. J. 2003, 9, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Cardenas, J.; Manzano-Agugliaro, F. The metagenomics worldwide research. Curr. Genet. 2017, 63, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Tetz, G.; Tetz, V. Prion-Like Domains in Phagobiota. Front. Microbiol. 2017, 8, 2239. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, G.; Greig, N.; Khan, T.; Hassan, I.; Tabrez, S.; Shakil, S.; Sheikh, I.; Zaidi, S.; Akram, M.; Jabir, N.; et al. Protein Misfolding and Aggregation in Alzheimer’s Disease and Type 2 Diabetes Mellitus. CNS Neurol. Disord. Drug Targets 2014, 13, 1280–1293. [Google Scholar] [CrossRef] [PubMed]
- Wemheuer, W.; Wrede, A.; Schulz-Schaeffer, W. Types and Strains: Their Essential Role in Understanding Protein Aggregation in Neurodegenerative Diseases. Front. Aging Neurosci. 2017, 9, 187. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Nonaka, T.; Masuda-Suzukake, M. Prion-like mechanisms and potential therapeutic targets in neurodegenerative disorders. Pharmacol. Ther. 2017, 172, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Bolton, D.; McKinley, M.; Prusiner, S. Identification of a protein that purifies with the scrapie prion. Science 1982, 218, 1309–1311. [Google Scholar] [CrossRef] [PubMed]
- Telling, G.; Scott, M.; Mastrianni, J.; Gabizon, R.; Torchia, M.; Cohen, F.; DeArmond, S.; Prusiner, S. Prion propagation in mice expressing human and chimeric PrP transgenes implicates the interaction of cellular PrP with another protein. Cell 1995, 83, 79–90. [Google Scholar] [CrossRef]
- Cheng, P.; Liu, C.; Zhao, M.; Eisenberg, D.; Nowick, J. Amyloid β-sheet mimics that antagonize protein aggregation and reduce amyloid toxicity. Nat. Chem. 2012, 4, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Messing, J. Phage M13 for the treatment of Alzheimer and Parkinson disease. Gene 2016, 583, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Bäuerl, C.; Collado, M.; Diaz Cuevas, A.; Viña, J.; Pérez Martínez, G. Shifts in gut microbiota composition in an APP/PSS1 transgenic mouse model of Alzheimer’s disease during lifespan. Lett. Appl. Microbiol. 2018, 66, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.; Perez-Chanona, E.; Muhlbauer, M.; Tomkovich, S.; Uronis, J.; Fan, T.; Campbell, B.; Abujamel, T.; Dogan, B.; Rogers, A.; et al. Intestinal Inflammation Targets Cancer-Inducing Activity of the Microbiota. Science 2012, 338, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Ghaisas, S.; Maher, J.; Kanthasamy, A. Gut microbiome in health and disease: Linking the microbiome–gut–brain axis and environmental factors in the pathogenesis of systemic and neurodegenerative diseases. Pharmacol. Ther. 2016, 158, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Tetz, G.; Brown, S.; Hao, Y.; Tetz, V. Parkinsons disease and bacteriophages as its overlooked contributors. bioRxiv 2018. [Google Scholar] [CrossRef]
- Asano, Y.; Hiramoto, T.; Nishino, R.; Aiba, Y.; Kimura, T.; Yoshihara, K.; Koga, Y.; Sudo, N. Critical role of gut microbiota in the production of biologically active, free catecholamines in the gut lumen of mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G1288–G1295. [Google Scholar] [CrossRef] [PubMed]
- Scheperjans, F. Gut microbiota, 1013 new pieces in the Parkinson’s disease puzzle. Curr. Opin. Neurol. 2016, 29, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Rub, U.; Gai, W.; Del Tredici, K. Idiopathic Parkinson’s disease: Possible routes by which vulnerable neuronal types may be subject to neuroinvasion by an unknown pathogen. J. Neural Transm. 2003, 110, 517–536. [Google Scholar] [CrossRef] [PubMed]
- Chopin, M.; Chopin, A.; Bidnenko, E. Phage abortive infection in lactococci: Variations on a theme. Curr. Opin. Microbiol. 2005, 8, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Samson, J.; Belanger, M.; Moineau, S. Effect of the Abortive Infection Mechanism and Type III Toxin/Antitoxin System AbiQ on the Lytic Cycle of Lactococcus lactis Phages. J. Bacteriol. 2013, 195, 3947–3956. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.; Serino, M.; Tilg, H.; Watson, A.; Wells, J. Intestinal permeability—A new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef] [PubMed]
- Fang, X. Potential role of gut microbiota and tissue barriers in Parkinson’s disease and amyotrophic lateral sclerosis. Int. J. Neurosci. 2015, 126, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, G.; Lee, S.; Mazmanian, S. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2015, 14, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Sursal, T.; Stearns-Kurosawa, D.J.; Itagaki, K.; Oh, S.Y.; Sun, S.; Kurosawa, S.; Hauser, C.J. Plasma Bacterial and Mitochondrial DNA Distinguish Bacterial Sepsis from Sterile SIRS and Quantify Inflammatory Tissue Injury in Nonhuman Primates. Shock 2013, 39, 55. [Google Scholar] [CrossRef] [PubMed]
- Tetz, G.; Artemenko, N.; Tetz, V. Effect of DNase and Antibiotics on Biofilm Characteristics. Antimicrob. Agents Chemother. 2008, 53, 1204–1209. [Google Scholar] [CrossRef] [PubMed]
- Anunobi, R.; Boone, B.; Cheh, N.; Tang, D.; Kang, R.; Loux, T.; Lotze, M.; Zeh, H. Extracellular DNA promotes colorectal tumor cell survival after cytotoxic chemotherapy. J. Surg. Res. 2018, 226, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Gallo, P.; Rapsinski, G.; Wilson, R.; Oppong, G.; Sriram, U.; Goulian, M.; Buttaro, B.; Caricchio, R.; Gallucci, S.; Tükel, Ç. Amyloid-DNA Composites of Bacterial Biofilms Stimulate Autoimmunity. Immunity 2015, 42, 1171–1184. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Llorente, C.; Hartmann, P.; Yang, A.; Chen, P.; Schnabl, B. Methods to determine intestinal permeability and bacterial translocation during liver disease. J. Immunol. Methods 2015, 421, 44–53. [Google Scholar] [CrossRef] [PubMed]
- De Paepe, M.; Leclerc, M.; Tinsley, C.; Petit, M. Bacteriophages: An underestimated role in human and animal health? Front. Cell. Infect. Microbiol. 2014, 4, 39. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Cai, Z.; Sanchez, A.; Zhang, T.; Wen, S.; Wang, J.; Yang, J.; Fu, S.; Zhang, D. A Novel Toll-like Receptor That Recognizes Vesicular Stomatitis Virus. J. Biol. Chem. 2010, 286, 4517–4524. [Google Scholar] [CrossRef] [PubMed]
- Mandl, J.; Barry, A.; Vanderford, T.; Kozyr, N.; Chavan, R.; Klucking, S.; Barrat, F.; Coffman, R.; Staprans, S.; Feinberg, M. Divergent TLR7 and TLR9 signaling and type I interferon production distinguish pathogenic and nonpathogenic AIDS virus infections. Nat. Med. 2008, 14, 1077–1087. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, R.; Guo, H.; Coulson, R.; Smyth, D.; Pekalski, M.; Burren, O.; Cutler, A.; Doecke, J.; Flint, S.; McKinney, E.; et al. A Type I Interferon Transcriptional Signature Precedes Autoimmunity in Children Genetically at Risk for Type 1 Diabetes. Diabetes 2014, 63, 2538–2550. [Google Scholar] [CrossRef] [PubMed]
- Rönnblom, L.; Alm, G. An etiopathogenic role for the type I IFN system in SLE. Trends Immunol. 2001, 22, 427–431. [Google Scholar] [CrossRef]
- Mills, S.; Shanahan, F.; Stanton, C.; Hill, C.; Coffey, A.; Ross, R. Movers and shakers. Gut Microbes 2013, 4, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Sobsey, M. Survival of prototype strains of somatic coliphage families in environmental waters and when exposed to UV low-pressure monochromatic radiation or heat. Water Res. 2011, 45, 3723–3734. [Google Scholar] [CrossRef] [PubMed]
- Riesenman, P.; Nicholson, W. Role of the Spore Coat Layers in Bacillus subtilis Spore Resistance to Hydrogen Peroxide, Artificial UV-C, UV-B, and Solar UV Radiation. Appl. Environ. Microbiol. 2000, 66, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Tetz, G.; Tetz, V. Introducing the sporobiota and sporobiome. Gut Pathog. 2017, 9, 38. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, G.; Moineau, S. Evolution of Lactococcus lactis Phages within a Cheese Factory. Appl. Environ. Microbiol. 2009, 75, 5336–5344. [Google Scholar] [CrossRef] [PubMed]
- Ogorzaly, L.; Gantzer, C. Development of real-time RT-PCR methods for specific detection of F-specific RNA bacteriophage genogroups: Application to urban raw wastewater. J. Virol. Methods 2006, 138, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Garneau, J.; Moineau, S. Bacteriophages of lactic acid bacteria and their impact on milk fermentations. Microb. Cell Fact. 2011, 10, S20. [Google Scholar] [CrossRef] [PubMed]
- Chehoud, C.; Dryga, A.; Hwang, Y.; Nagy-Szakal, D.; Hollister, E.; Luna, R.; Versalovic, J.; Kellermayer, R.; Bushman, F. Transfer of Viral Communities between Human Individuals during Fecal Microbiota Transplantation. mBio 2016, 7, e00322-16. [Google Scholar] [CrossRef] [PubMed]
- Michel, J.; Yeh, P.; Chait, R.; Moellering, R.; Kishony, R. Drug interactions modulate the potential for evolution of resistance. Proc. Natl. Acad. Sci. USA 2008, 105, 14918–14923. [Google Scholar] [CrossRef] [PubMed]
- Stanton, T.; Humphrey, S.; Sharma, V.; Zuerner, R. Collateral Effects of Antibiotics: Carbadox and Metronidazole Induce VSH-1 and Facilitate Gene Transfer among Brachyspira hyodysenteriae Strains. Appl. Environ. Microbiol. 2008, 74, 2950–2956. [Google Scholar] [CrossRef] [PubMed]
- Migliore, L.; Copped, F. Genetic and environmental factors in cancer and neurodegenerative diseases. Mutat. Res. Rev. Mutat. Res. 2002, 512, 135–153. [Google Scholar] [CrossRef]


{kind=link}
| Main Causes of Bacteriophages Infections | Comments |
|---|---|
| Infection by lytic phages | Primary bacteriophage infection due to environmental bacterial viruses |
| Infection by bacteria carrying prophages in their DNA | Primary infection by environmental bacteria harboring prophages |
| Induction of prophages | Can be due to a variety of different internal and external triggers, leading to progeny release, an increase in the number of free phages leading to significant microbiota alterations [87] |
| Increased translocation of phages to biological fluids | Can be due to increased intestinal and blood-brain barrier permeability [43,81] |
| Alteration of the microbiota | Alteration of microbiota composition (following emergence of new bacteria, antibiotics treatment, diet, etc.) leading to a shift in numbers of prophages or lytic phages |
| Mutations in prophages leading to Abi-escape phage mutants | Phages able to overcome bacterial defense systems [72,73] |
| Changes in macroorganism sensitivity to direct and indirect bacteriophage effects | Human sensitivity to bacteriophage infection can be altered through a variety of internal factors or factors associated with the microbiota |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tetz, G.; Tetz, V. Bacteriophages as New Human Viral Pathogens. Microorganisms 2018, 6, 54. https://doi.org/10.3390/microorganisms6020054
Tetz G, Tetz V. Bacteriophages as New Human Viral Pathogens. Microorganisms. 2018; 6(2):54. https://doi.org/10.3390/microorganisms6020054
Chicago/Turabian StyleTetz, George, and Victor Tetz. 2018. "Bacteriophages as New Human Viral Pathogens" Microorganisms 6, no. 2: 54. https://doi.org/10.3390/microorganisms6020054
APA StyleTetz, G., & Tetz, V. (2018). Bacteriophages as New Human Viral Pathogens. Microorganisms, 6(2), 54. https://doi.org/10.3390/microorganisms6020054