Fungal Disease Prevention in Seedlings of Rice (Oryza sativa) and Other Grasses by Growth-Promoting Seed-Associated Endophytic Bacteria from Invasive Phragmites australis

 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Bacterial Isolates
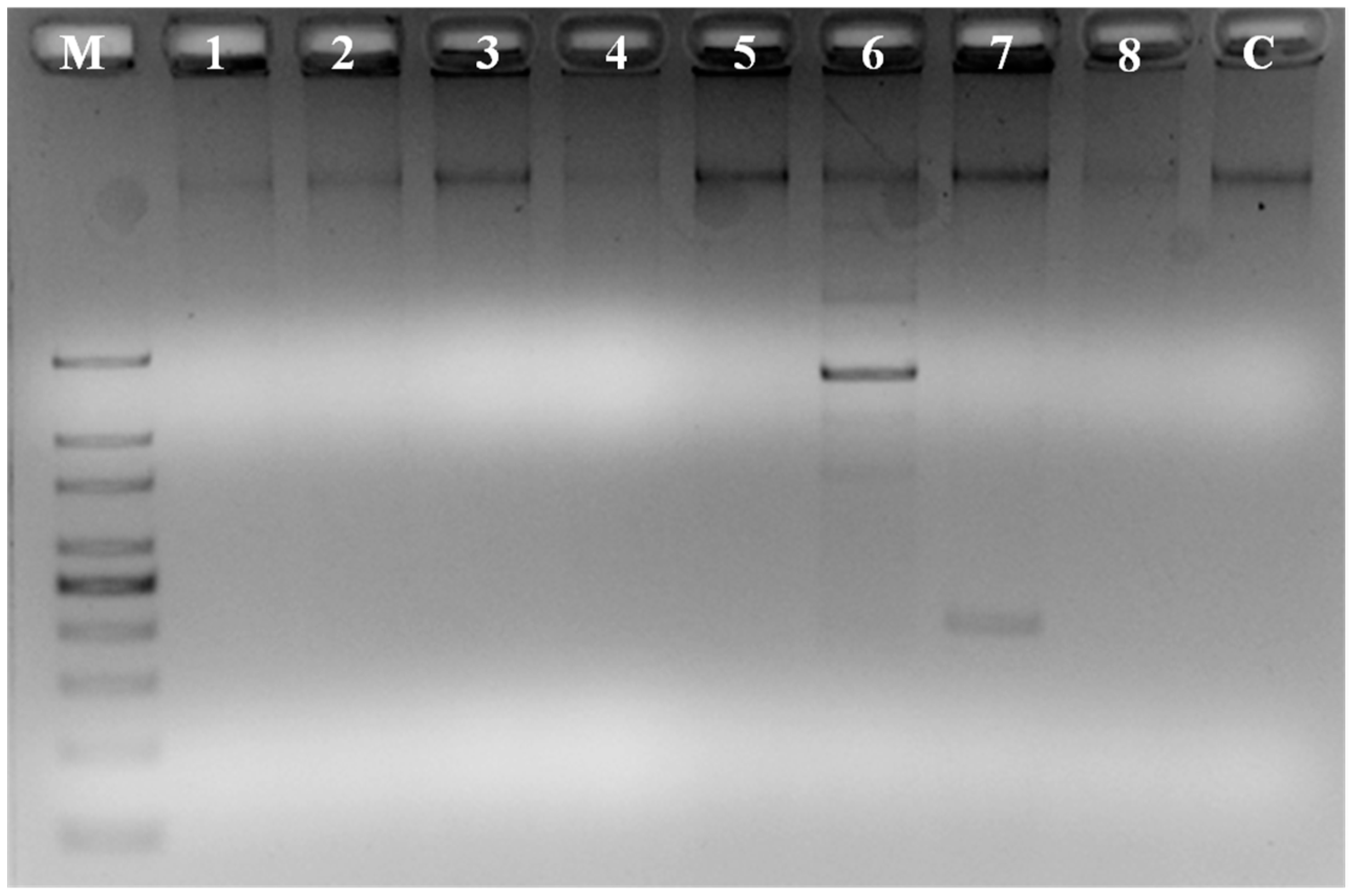
2.3. Molecular Identification of Bacteria
2.4. Cleaning of Rice, Bermuda Grass and Annual Bluegrass Seeds
2.5. Seedling Growth Promotion Experiments in Agarose Plates and Magenta Boxes
2.6. Phosphate Solubilization Activity
2.7. Antifungal Activity of Strain SY1
2.8. Antifungal Activity Assays: Agar Diffusion, Volatiles and HCN Production
2.9. Strain SY1 Antifungal Activity on Rice, Bermuda Grass and Annual Bluegrass Seedlings
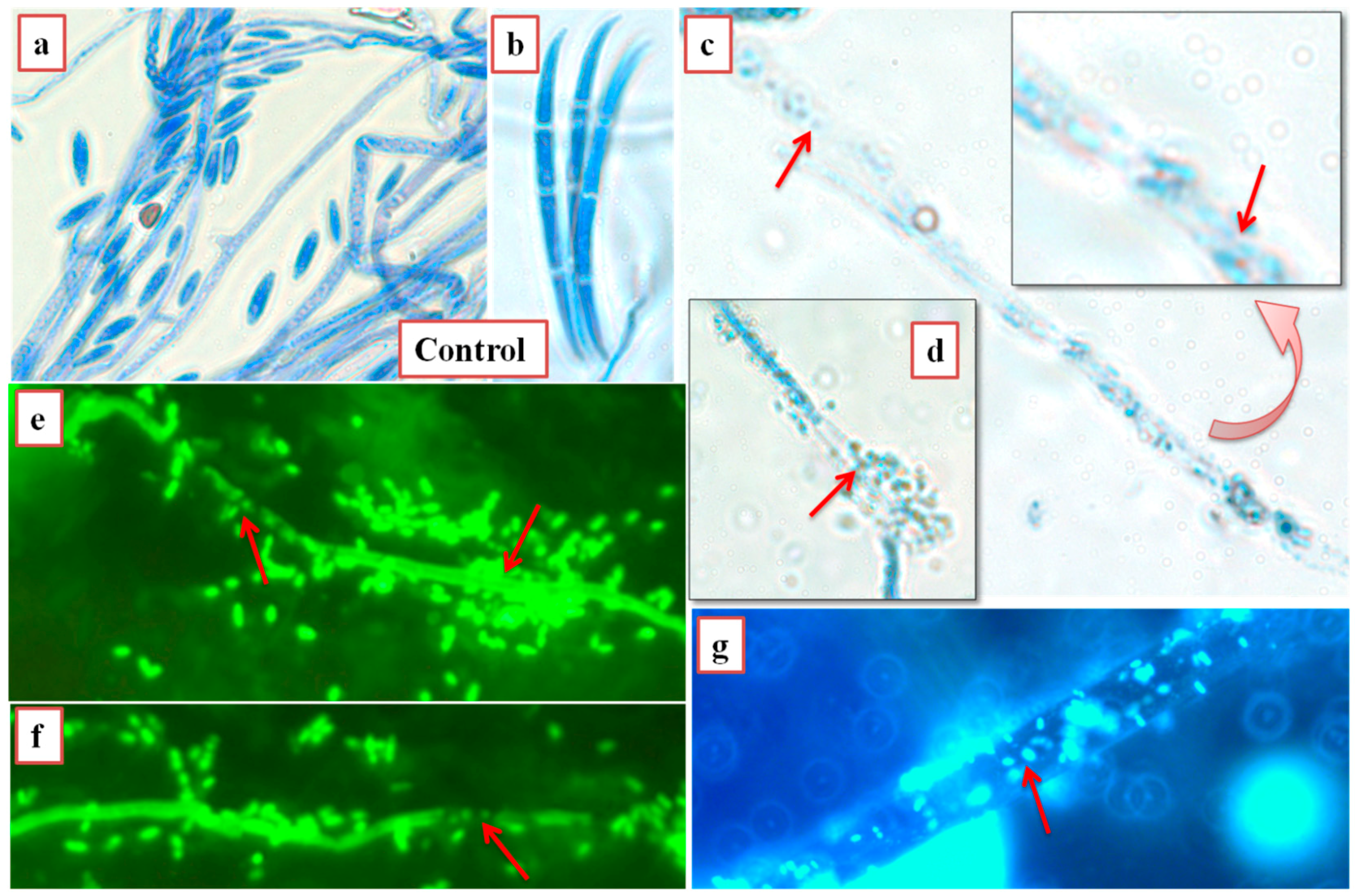
2.10. Reactive Oxygen and SYTO13® Staining to Visualize Bacteria
2.11. Screening for Antifungal Genes in Strain SY1
2.12. Extraction and MALDI-TOF Analysis for Lipopeptides
3. Results
3.1. Effect of Bacteria on Rice and Bermuda Grass Seedling Growth and Development
3.2. Phosphate Solubilization
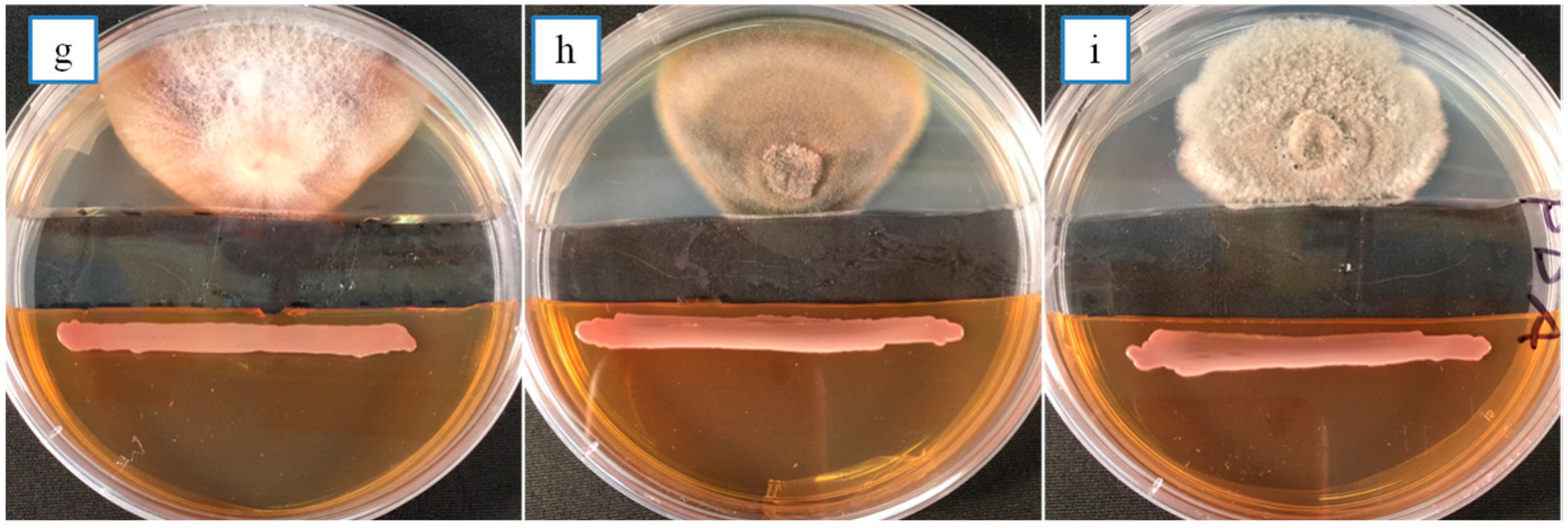
3.3. Antifungal Activity of Bacteria
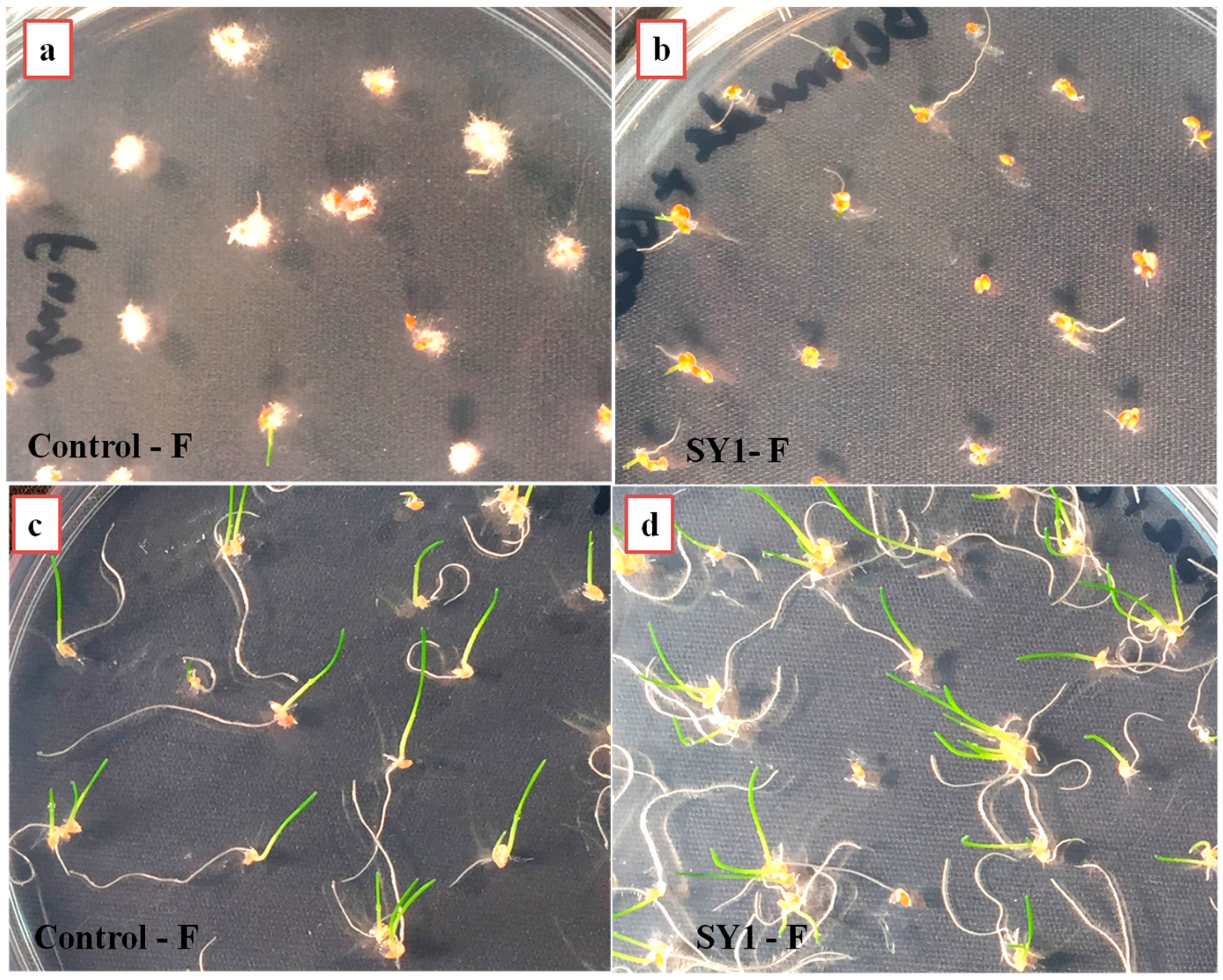
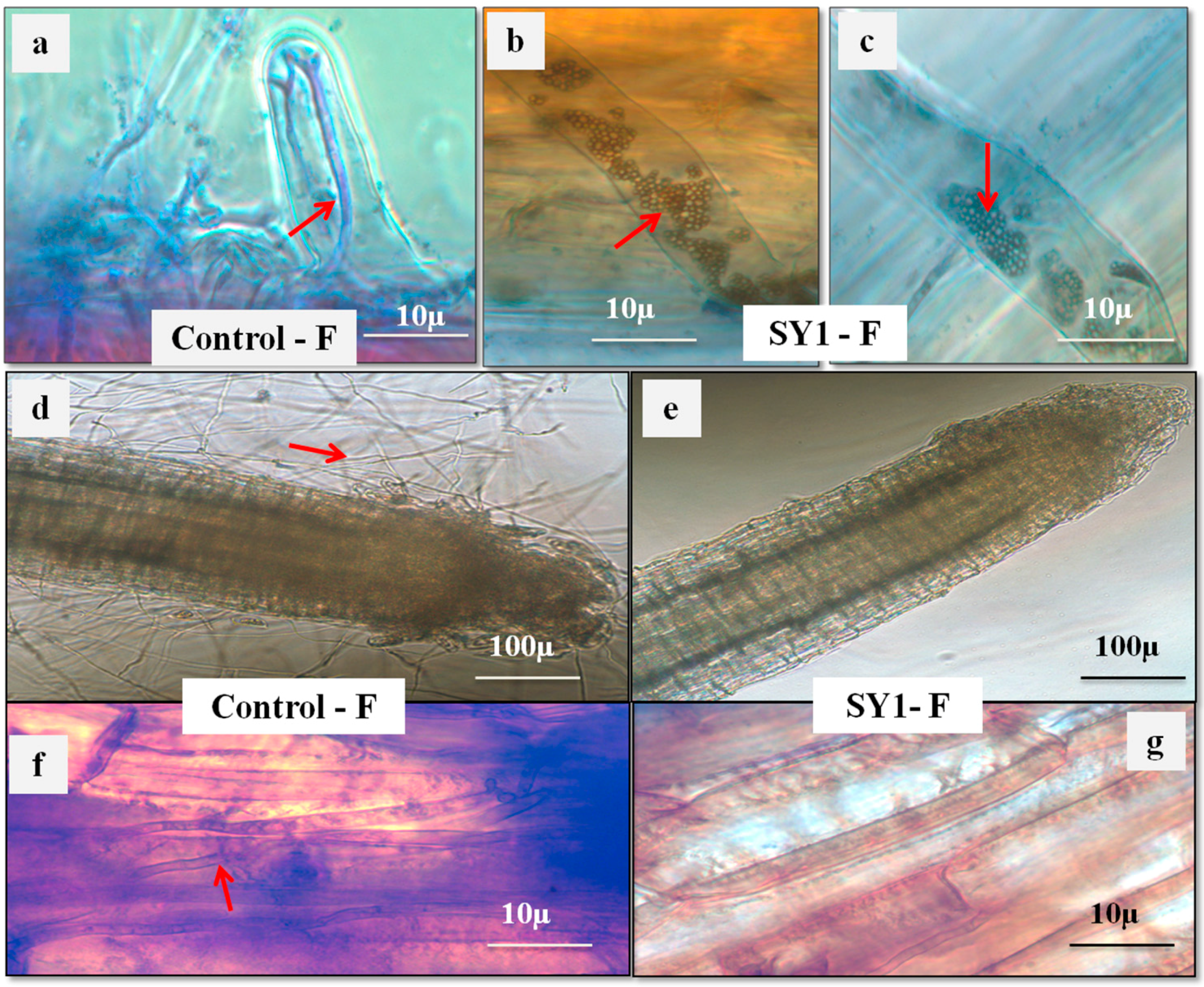
3.4. SY1 Exhibits Antifungal Activity in Grass Seedling Roots against Fusarium oxysporum
3.5. Screening for Antibiotic Genes in SY1
3.6. MALDI-TOF Analysis of SY1
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Imolehin, E.D. Rice seed borne fungi and their effect on seed germination. Plant Dis. 1983, 67, 1334–1336. [Google Scholar] [CrossRef]
- Bigirimana, V.P.; Hua, G.K.H.; Nyamangyoku, O.I.; Höfte, M. Rice heath rot: An emerging ubiquitous destructive disease complex. Front. Plant Sci. 2015, 6, 1066. [Google Scholar] [CrossRef] [PubMed]
- Spence, C.; Alff, E.; Johnson, C.; Ramos, C.; Donofrio, N.; Sundaresan, V.; Bais, H. Natural rice rhizospheric microbes suppress rice blast infections. BMC Plant Biol. 2014, 14, 130. [Google Scholar] [CrossRef] [PubMed]
- Kandel, S.L.; Joubert, P.M.; Doty, L.S. Bacterial endophyte colonization and distribution within plants. Microorganisms 2017, 5, 77. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.K.; Henson, J.; van Volkenburgh, E.; Hoy, M.; Wright, L.; Beckwith, F.; Kim, Y.-O.; Redman, R.S. Stress tolerance in plants via habitat-adapted symbiosis. ISME J. 2008, 2, 404–416. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.; White, J.F.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Puente, M.E.; Lib, C.Y.; Bashan, Y. Endophytic bacteria in cacti seeds can improve the development of cactus seedlings. Environ. Exp. Bot. 2009, 66, 402–408. [Google Scholar] [CrossRef]
- White, J.F.; Kingsley, K.I.; Kowalski, K.P.; Irizarry, I.; Micci, A.; Soares, M.A.; Bergen, M.S. Disease protection and allelopathic interactions of seed-transmitted endophytic Pseudomonads of invasive seed grass (Phragmites australis). Plant Soil 2018, 422, 195–208. [Google Scholar] [CrossRef]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.; Levy, S.B.; Jackson, R.W. Pseudomonas genomes: Diverse and adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef] [PubMed]
- Gond, S.K.; Bergen, M.; Torres, M.S.; White, J.F. Effect of bacterial endophyte on expression of defense genes in Indian popcorn against Fusarium moniliforme. Symbiosis 2015, 66, 133–140. [Google Scholar] [CrossRef]
- Ryu, C.; Farag, M.A.; Hu, C.; Reddy, M.S.; Kloepper, J.W.; Paré, P.W. Bacterial volatiles induce systemic resistance in Arabidopsis. Plant Physiol. 2004, 134, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Prieto, P.; Schilirò, E.; Maldonado-González, M.; Valderrama, R.; Barroso-Albarracín, J.B.; Mercado-Blanco, J. Root hairs play a key role in the endophytic colonization of olive roots by Pseudomonas spp. with biocontrol activity. Microb. Ecol. 2011, 62, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Kastell, A.; Knorr, D.; Smetanska, I. Exudation: An expanding technique for continuous production and release of secondary metabolites from plant cell suspension and hairy root cultures. Plant Cell Rep. 2012, 31, 461–477. [Google Scholar] [CrossRef] [PubMed]
- Pikovskaya, R.I. Mobilization of phosphorus in soil connection with the vital activity of some microbial species. Microbiologiya 1948, 17, 362–370. [Google Scholar]
- Whipps, J.M. Developments in the biological control of soil-borne plant pathogens. Adv. Bot. Res. 1997, 26, 1–134. [Google Scholar]
- Bakker, A.W.; Schippers, B. Microbial cyanide production in the rhizosphere in relation to potato yield reduction and Pseudomonas spp.-mediated plant growth-stimulation. Soil Biol. Biochem. 1987, 19, 451–457. [Google Scholar] [CrossRef]
- White, J.F.; Chen, Q.; Torres, M.S.; Mattera, R.; Irizarry, I.; Tadych, M.; Bergen, M. Collaboration between grass seedlings and rhizobacteria to scavenge organic nitrogen in soils. AoB PLANTS 2015, 7, plu093. [Google Scholar] [CrossRef] [PubMed]
- De Souza, J.T.; Raaijmakers, J.M. Polymor-phisms within the prnD, and pltC genes from pyrrolnitrin and pyoluteorin-producing Pseudomonas and Burkholderia spp. FEMS Microbiol. Ecol. 2003, 43, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Mavrodi, D.V.; Peever, T.L.; Mavrodi, O.V.; Parejko, J.A.; Raaijmakers, J.M.; Lemanceau, P.; Mazurier, S.; Heide, L.; Blankenfeldt, W.; Weller, D.M.; et al. Diversity and evolution of the phenazine biosynthesis pathway. Appl. Environ. Microbiol. 2010, 76, 866–879. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Mele, P.M.; Crowley, D.E. Application of PCR primer sets for detection of Pseudomonas sp. functional genes in the plant rhizosphere. J. Agric. Chem. Environ. 2013, 2, 8–15. [Google Scholar]
- Hurek, T.; Handley, L.L.; Reinhold-Hurek, B.; Piché, Y. Azoarcus grass endo-phytes contribute fixed nitrogen to the plant in an unculturable state. Mol. Plant Microbe Interact. 2002, 15, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Bernard, S.M.; Andersen, G.L.; Chen, W. Developing microbe–plant interactions for applications in plant-growth promotion and disease control, production of useful compounds, remediation and carbon sequestration. Microb. Biotechnol. 2009, 2, 428–440. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.K.; Kingsley, K.; Irizarry, I.; Bergen, M.; Kharwar, R.N.; White, J.F. Seed-vectored endophytic bacteria modulate development of rice seedlings. J. Appl. Microbiol. 2017, 122, 1680–1691. [Google Scholar] [CrossRef] [PubMed]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.; et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 422, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Nestler, J.; Keyes, S.D.; Wissuwa, M. Root hair formation in rice (Oryza sativa L.) differs between root types and is altered in artificial growth conditions. J. Exp. Bot. 2016, 67, 3699–3708. [Google Scholar] [CrossRef] [PubMed]
- Paungfoo-Lonhienne, C.; Schmidt, S.; Webb, R.; Lonhienne, T. Rhizophagy—A new dimension of plant-microbe interactions. In Molecular Microbial Ecology of the Rhizosphere; de Briujn, F.J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013. [Google Scholar]
- Mercado-Blanco, J.; Prieto, P. Bacterial endophytes and root hairs. Plant Soil 2012, 361, 301–306. [Google Scholar] [CrossRef]
- Park, J.Y.; Oh, S.A.; Anderson, A.J.; Neiswender, J.; Kim, J.C.; Kim, Y.C. Production of the antifungal compounds phenazine and pyrrolnitrin from Pseudomonas chlororaphis O6 is differentially regulated by glucose. Lett. Appl. Microbiol. 2011, 52, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Asadhi, S.; Reddy, B.V.B.; Sivaprasad, Y.; Prathyusha, M.; Krishna, T.M.; Kumar, K.V.K.; Reddy, K.R. Characterization, genetic diversity and antagonistic potential of 2,4-diacetylphloroglucinol producing Pseudomonas fluorescens isolates in groundnut-based cropping systems of Andhra Pradesh, India. Arch. Phytopathol. Plant Prot. 2013, 46, 1966–1977. [Google Scholar] [CrossRef]
- Mandal, S.M.; Sharma, S.; Pinnaka, A.K.; Kumari, A.; Korpole, S. Isolation and characterization of diverse antimicrobial lipopeptides produced by Citrobacter and Enterobacter. BMC Microbiol. 2013, 13, 152. [Google Scholar] [CrossRef] [PubMed]
- Fernando, W.G.D.; Ramarathnam, R.K.A.S.; Savchuk, S.C. Identification and use of potential bacterial organic antifungal volatiles in biocontrol. Soil Biol. Biochem. 2005, 37, 955–964. [Google Scholar] [CrossRef]
- Uroz, S.; Oger, P.M.; Chapelle, E.; Adeline, M.T.; Faure, D.; Dessaux, Y. A Rhodococcus qsdA-encoded enzyme defines a novel class of large-spectrum quorum-quenching lactonases. Appl. Environ. Microbiol. 2008, 74, 1357–1366. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Geotropic Response (%) | Root Length (Mean ± SD cm) | Root Hairs and Branching * | Shoot Length (Mean ± SD cm) | Phosphate Solubilization |
|---|---|---|---|---|---|
| No bacteria | 10 | 1.46 ± 0.53 | + | 1.45 ± 0.51 | − |
| West9 | 20 | 2.28 ± 0.86 | ++ | 2.13 ± 0.75 | + |
| SLB4 | 40 | 2.39 ± 0.91 | +++ | 2.37 ± 0.90 | + |
| Microbac | 20 | 2.30 ± 0.84 | ++ | 2.18 ± 0.79 | + |
| SY1 | 25 | 2.23 ± 083 | +++ | 2.19 ± 0.76 | + |
| SY5 | 10 | 1.47 ± 0.62 | None | 1.47 ± 0.65 | + |
| SY4 | 0.0 | 1.37 ± 0.60 | None | 1.44 ± 0.63 | + |
| SLB6 | 25 | 1.33 ± 0.56 | ++ | 1.25 ± 0.42 | + |
| RiY3 | 0.0 | 2.50 ± 0.73 | None | 2.68 ± 0.74 | + |
| ROLB13w | 10 | 0.93 ± 0.58 | None | 1.03 ± 0.53 | + |
| Treatment | Geotropic Response (%) | Root Length (Mean ± sd cm) | Root Hairs and Branching * | Shoot Length (Mean ± sd cm) |
|---|---|---|---|---|
| No bacteria | 35 | 1.53 ± 0.35 | + | 1.58 ± 0.37 |
| West9 | 35 | 1.87 ± 0.58 | +++ | 1.88 ± 0.59 |
| SLB4 | 50 | 2.23 ± 0.58 | +++ | 2.43 ± 0.92 |
| Microbac | 40 | 1.65 ± 0.83 | + | 1.36 ± 0.67 |
| SY1 | 40 | 2.45 ± 0.55 | +++ | 2.43 ± 0.70 |
| SLB6 | 60 | 1.99 ± 0.59 | +++ | 2.24 ± 0.47 |
| Treatments | % Inhibition | ||
|---|---|---|---|
| F. oxysporum | Curvularia sp. | Alternaria sp. | |
| Antagonism | 85.71 | 86.33 | 82.14 |
| Agar diffusion | 52.38 | 63.63 | 55.55 |
| Treatments | % Infection of Seedlings | ||
|---|---|---|---|
| Rice | Bermuda Grass | Annual Bluegrass | |
| Control (Fungi) | 51.50 | 100.00 | 90.00 |
| Treatment (SY1 + Fungi) | 18.75 | 15.00 | 10.00 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verma, S.K.; Kingsley, K.L.; Bergen, M.S.; Kowalski, K.P.; White, J.F. Fungal Disease Prevention in Seedlings of Rice (Oryza sativa) and Other Grasses by Growth-Promoting Seed-Associated Endophytic Bacteria from Invasive Phragmites australis. Microorganisms 2018, 6, 21. https://doi.org/10.3390/microorganisms6010021
Verma SK, Kingsley KL, Bergen MS, Kowalski KP, White JF. Fungal Disease Prevention in Seedlings of Rice (Oryza sativa) and Other Grasses by Growth-Promoting Seed-Associated Endophytic Bacteria from Invasive Phragmites australis. Microorganisms. 2018; 6(1):21. https://doi.org/10.3390/microorganisms6010021
Chicago/Turabian StyleVerma, Satish K., Kathryn L. Kingsley, Marshall S. Bergen, Kurt P. Kowalski, and James F. White. 2018. "Fungal Disease Prevention in Seedlings of Rice (Oryza sativa) and Other Grasses by Growth-Promoting Seed-Associated Endophytic Bacteria from Invasive Phragmites australis" Microorganisms 6, no. 1: 21. https://doi.org/10.3390/microorganisms6010021
APA StyleVerma, S. K., Kingsley, K. L., Bergen, M. S., Kowalski, K. P., & White, J. F. (2018). Fungal Disease Prevention in Seedlings of Rice (Oryza sativa) and Other Grasses by Growth-Promoting Seed-Associated Endophytic Bacteria from Invasive Phragmites australis. Microorganisms, 6(1), 21. https://doi.org/10.3390/microorganisms6010021