The Science behind the Probiotic Strain Bifidobacterium animalis subsp. lactis BB-12®

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
1.1. The Microbiota
1.2. Probiotics
1.3. Strain Level
2. Taxonomy & Characterization
2.1. Taxonomy
2.2. Origin & Selection
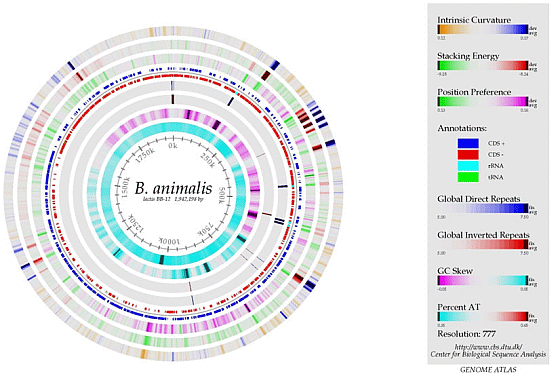
2.3. The Genome
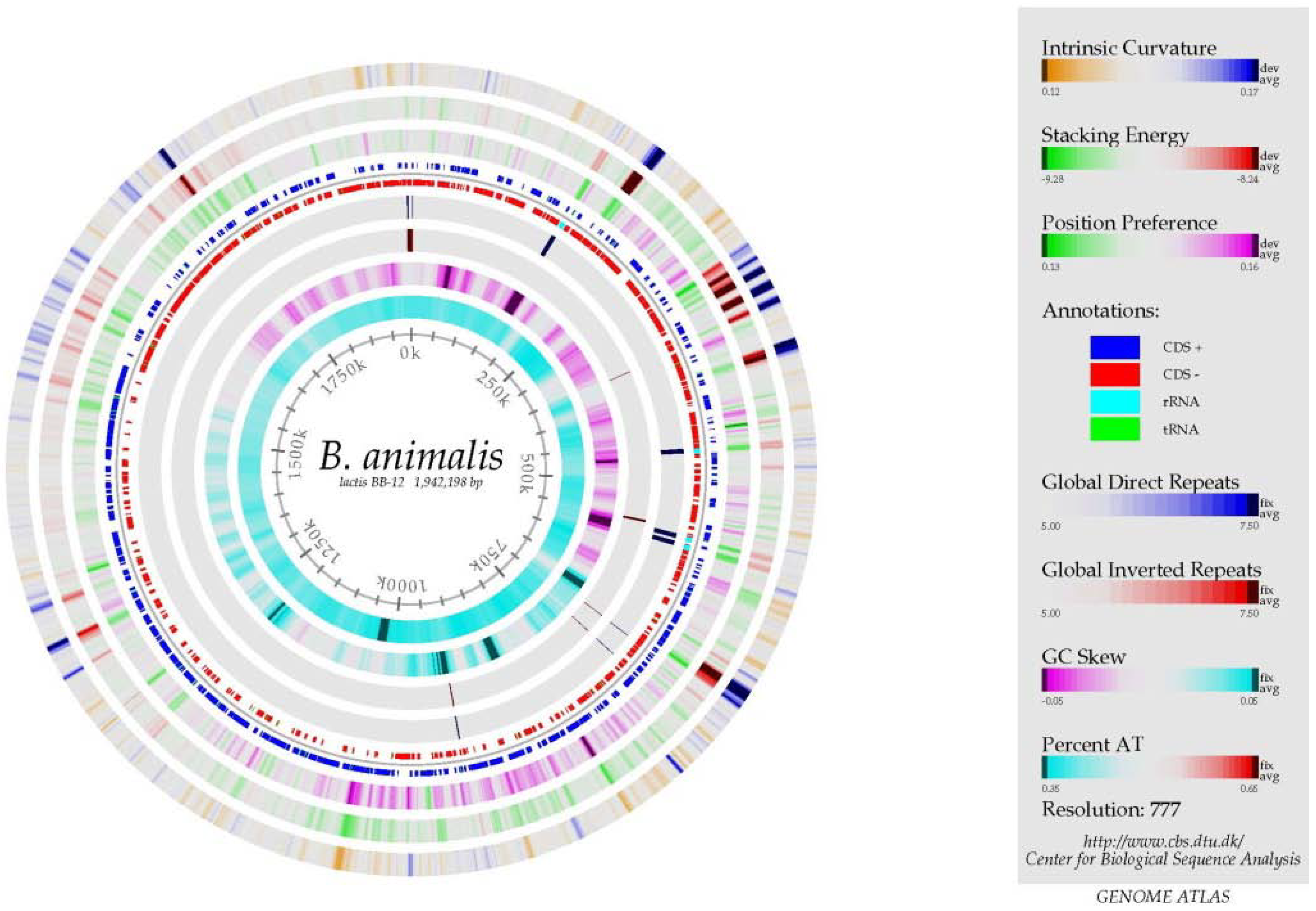
3. Strain Characteristics & Mechanisms
3.1. Acid and Bile Tolerance
3.2. Bile Salt Hydrolase

3.3. Mucus Adhesion
3.4. Pathogen Inhibition
3.5. Barrier Function Enhancement
3.6. Immune Interactions
4. Efficacy
4.1. Proven Efficacy
4.2. Survival in the Gastrointestinal Tract
4.3. Modulation of Intestinal Microbiota
4.4. Gastrointestinal Function
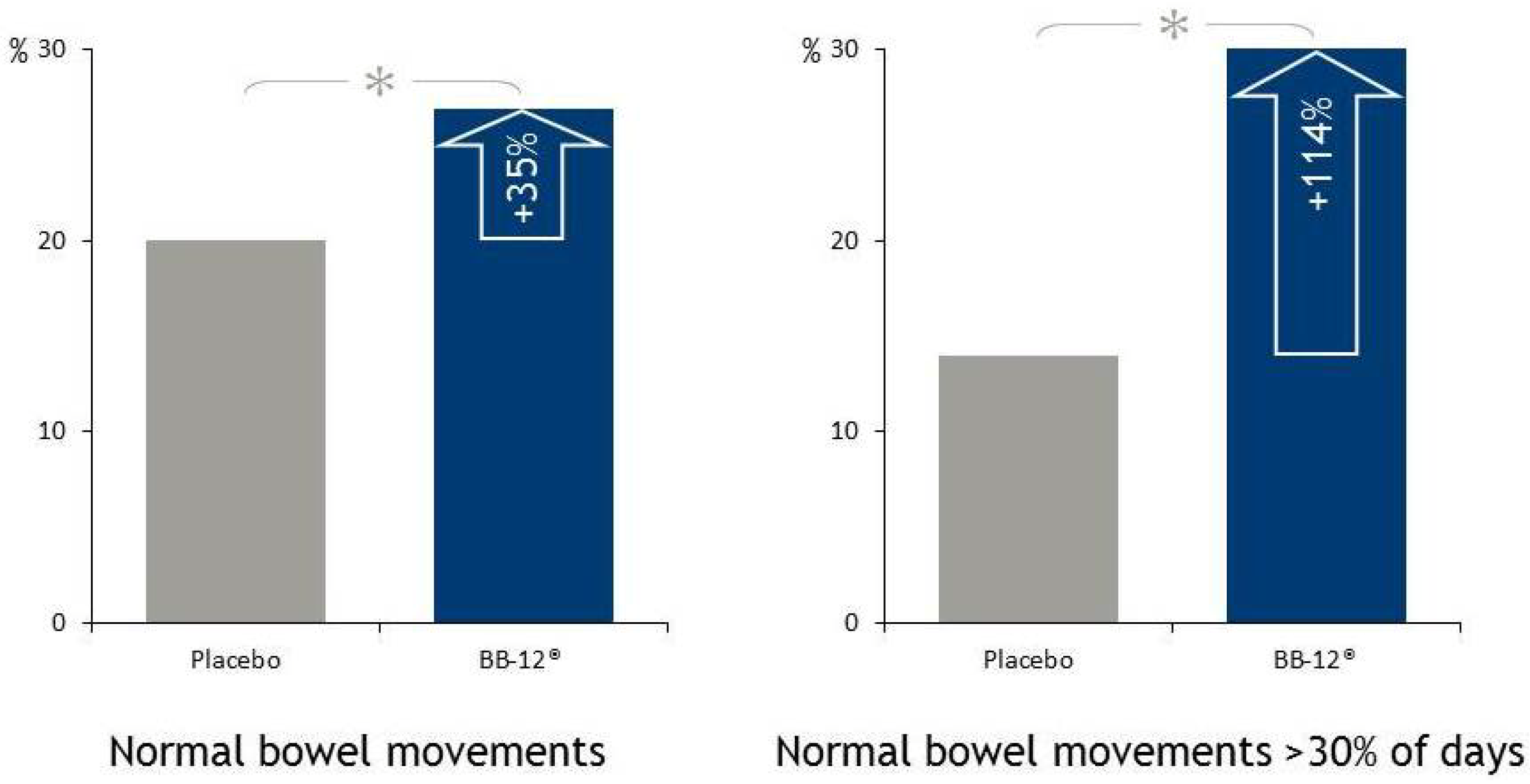
4.4.1. Bowel Function
4.4.2. Diarrhea in Infants and Children
4.4.3. Antibiotic-Associated Diarrhea
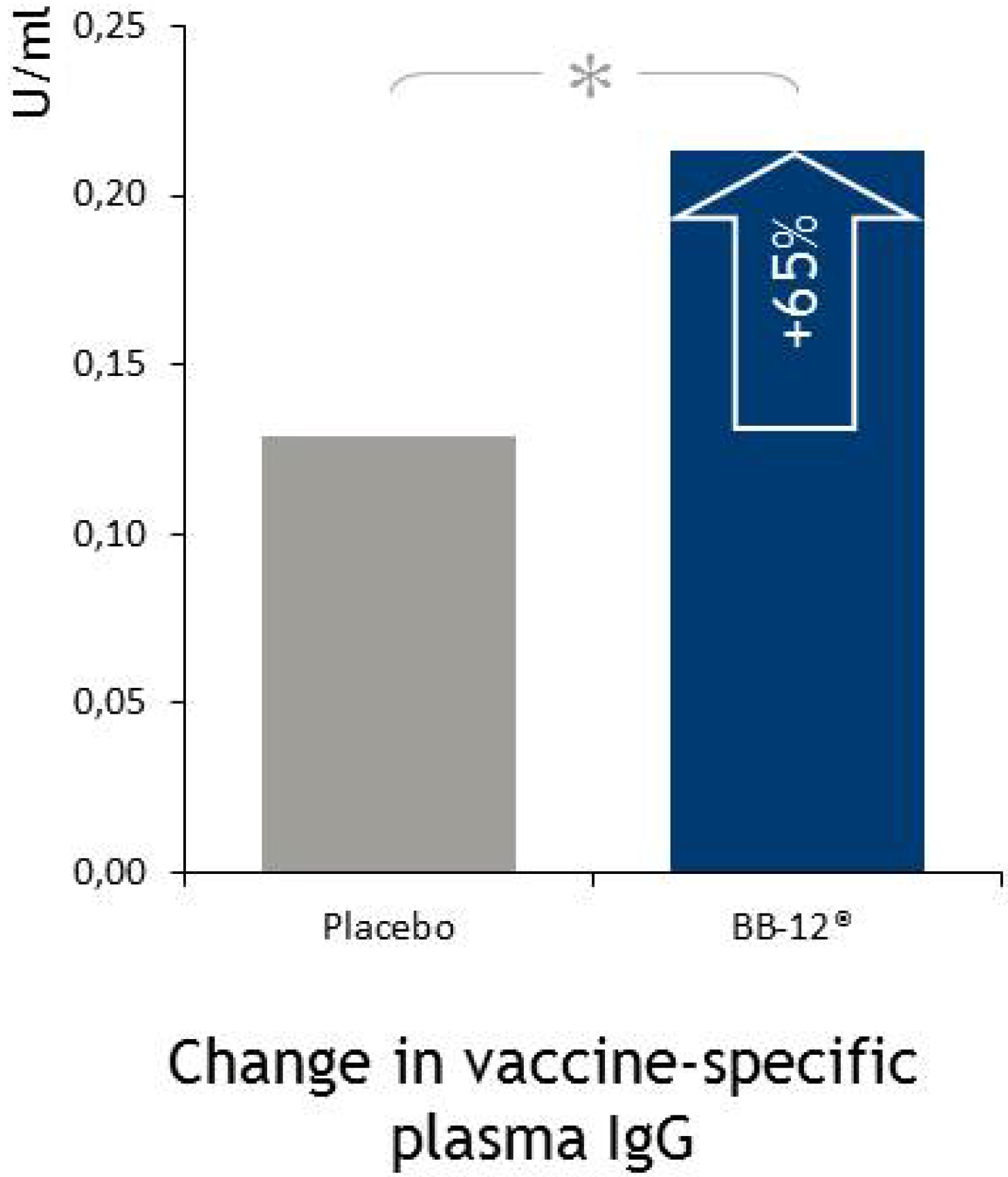
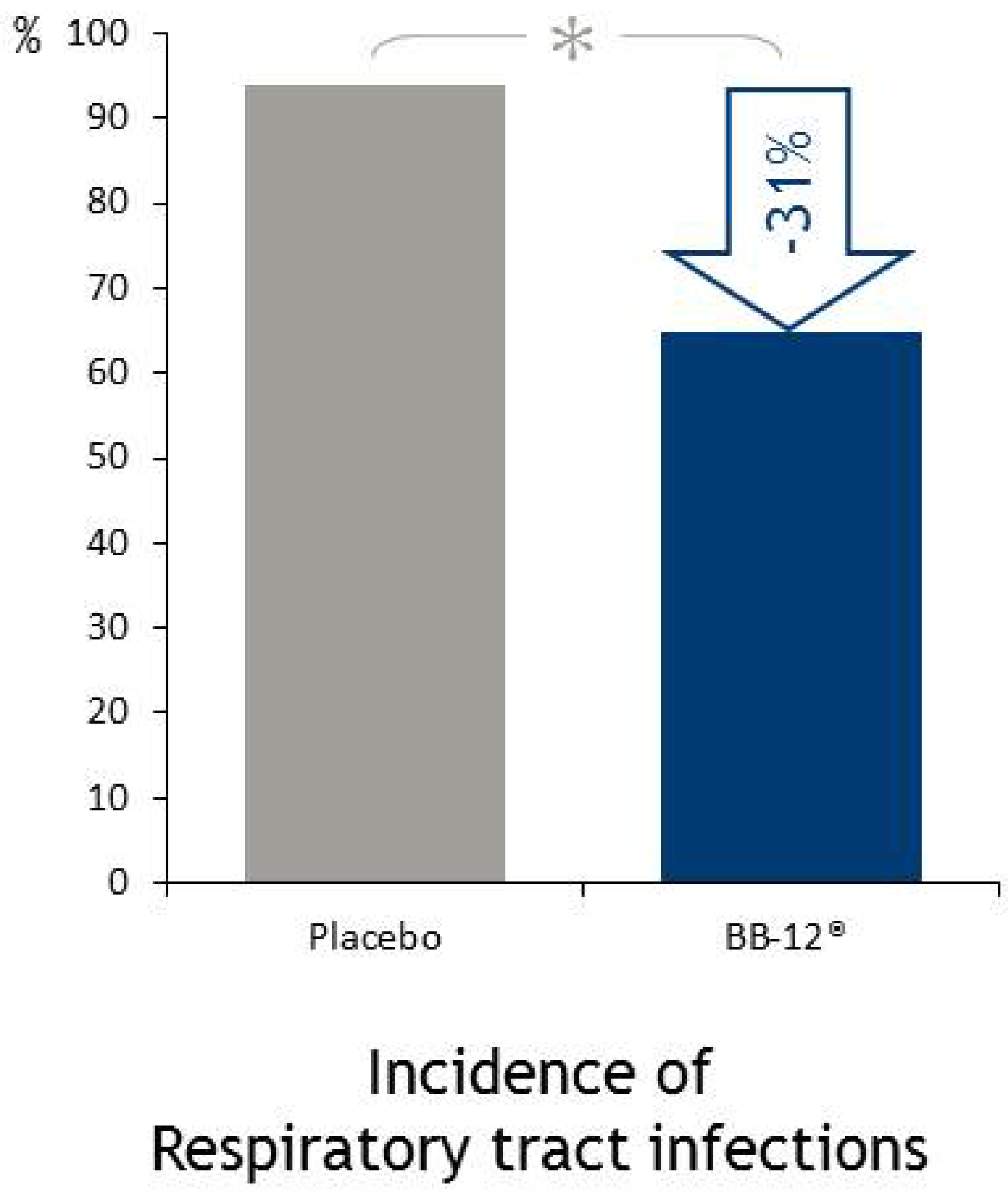
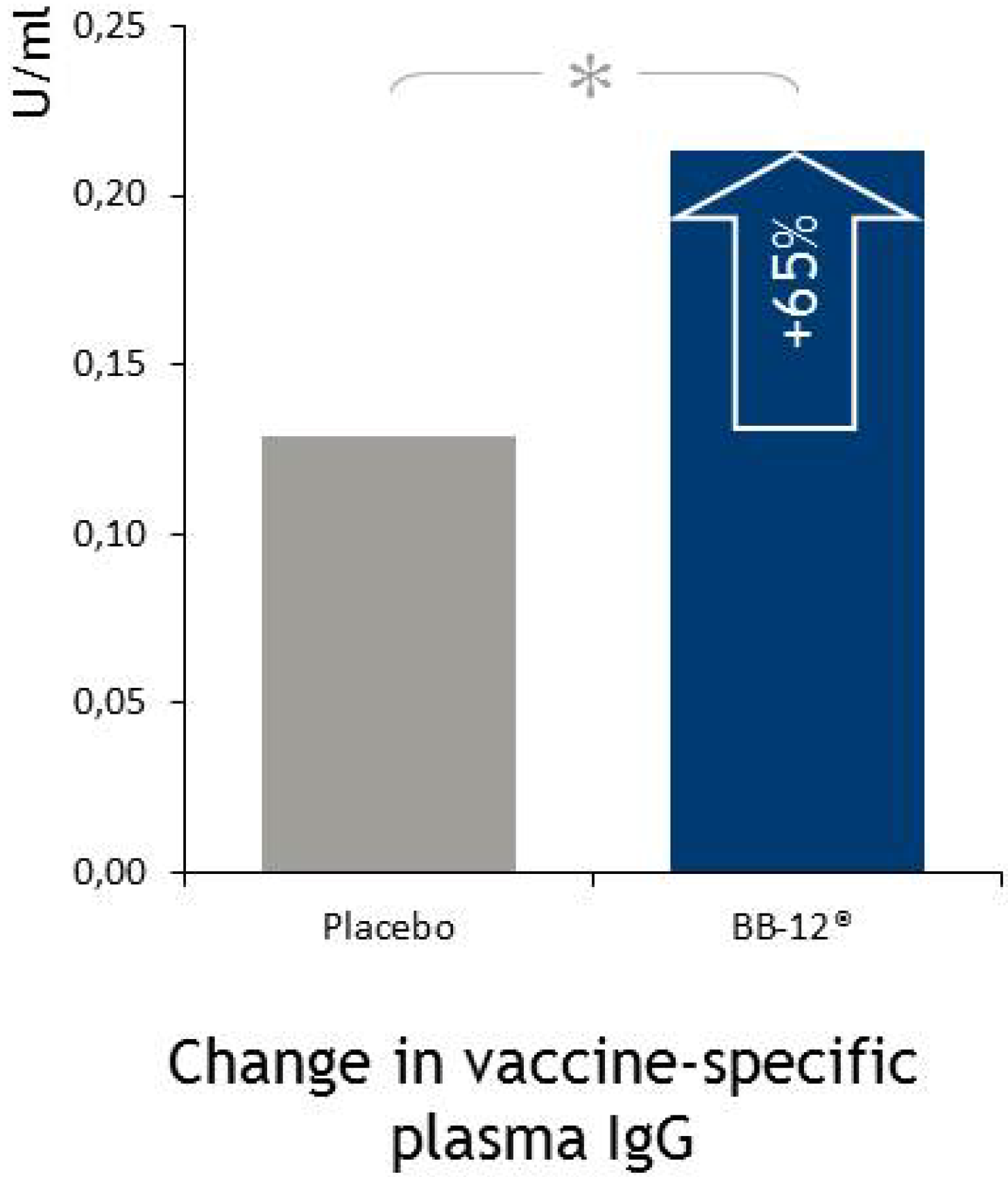
4.5. Immune Function
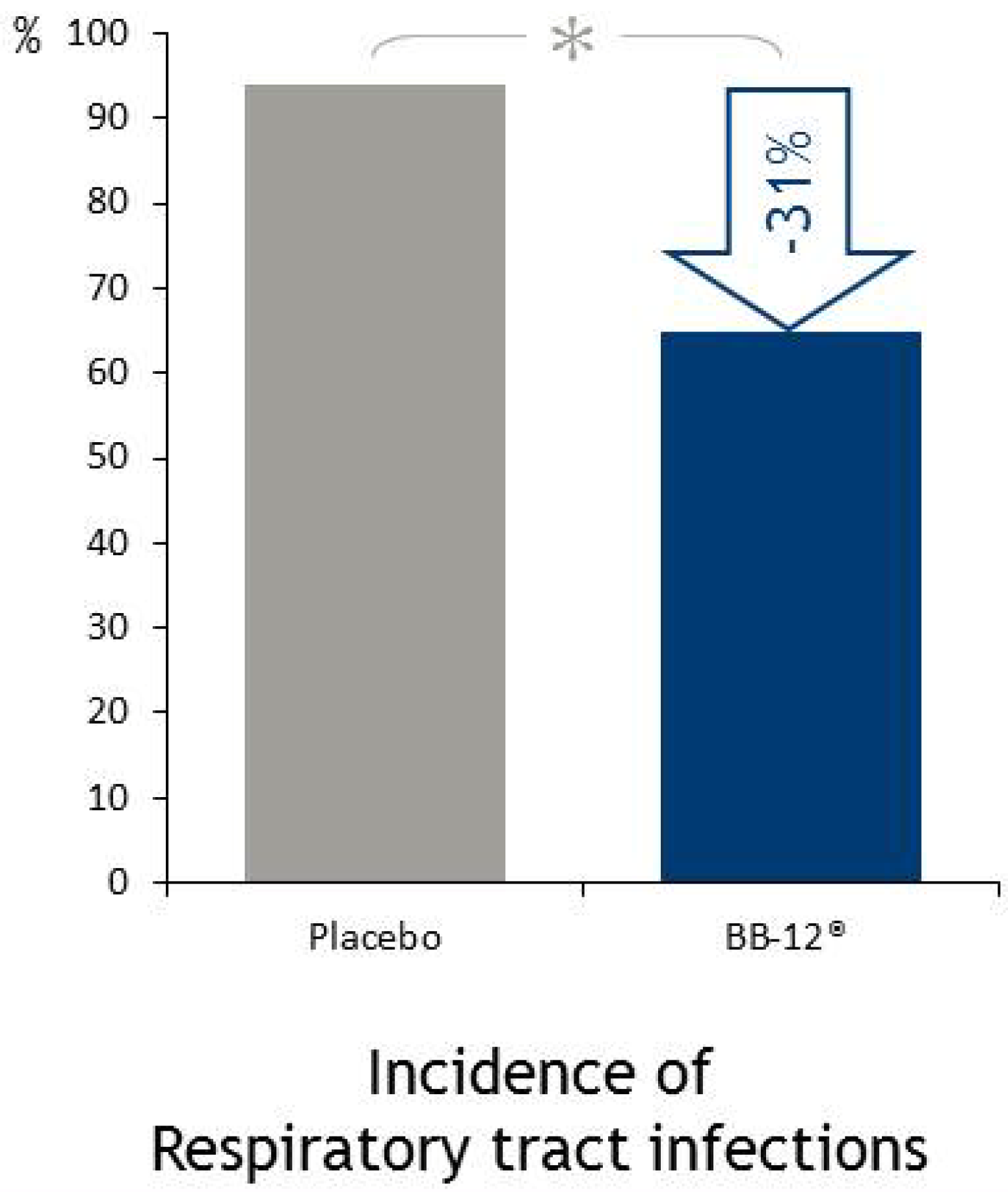
4.5.1. Respiratory Infections
5. Conclusions
Conflicts of Interest
References
- Health and Nutrition Properties of Probiotics in Food including Powder Milk with Live Lactic Acid Bacteria; FAO/WHO: Geneva, Switzerland, 2001.
- Reuter, G. The Lactobacillus and Bifidobacterium microflora of the human intestine: Composition and succession. Curr. Issues Intest. Microbiol. 2001, 2, 43–53. [Google Scholar]
- Garrigues, C.; Stuer-Lauridsen, B.; Johansen, E. Characterisation of Bifidobacterium animalis subsp. lactis BB-12 and other probiotic bacteria using genomics, transcriptomics and proteomics. Aust. J. Dairy Technol. 2005, 60, 84–92. [Google Scholar]
- Garrigues, C.; Johansen, E.; Pedersen, M.B. Complete genome sequence of Bifidobacterium animalis subsp. lactis BB-12, a widely consumed probiotic strain. J. Bacteriol. 2010, 192, 2467–2468. [Google Scholar] [CrossRef]
- Pedersen, M.B.; Iversen, S.L.; Sorensen, K.I.; Johansen, E. The long and winding road from the research laboratory to industrial applications of lactic acid bacteria. FEMS Microbiol. Rev. 2005, 29, 611–624. [Google Scholar] [CrossRef]
- Garrigues, C.; Johansen, E.; Crittenden, R. Pangenomics—An avenue to improved industrial starter cultures and probiotics. Curr. Opin. Biotechnol. 2013, 24, 187–191. [Google Scholar] [CrossRef]
- Vernazza, C.L.; Gibson, G.R.; Rastall, R.A. Carbohydrate preference, acid tolerance and bile tolerance in five strains of Bifidobacterium. J. Appl. Microbiol. 2006, 100, 846–853. [Google Scholar] [CrossRef]
- Matsumoto, M.; Ohishi, H.; Benno, Y. H+-ATPase activity in Bifidobacterium with special reference to acid tolerance. Int. J. Food Microbiol. 2004, 93, 109–113. [Google Scholar] [CrossRef]
- Vinderola, C. Lactic acid starter and probiotic bacteria: A comparative “in vitro” study of probiotic characteristics and biological barrier resistance. Food Res. Int. 2003, 36, 895–904. [Google Scholar] [CrossRef]
- Laparra, J.M.; Sanz, Y. Comparison of in vitro models to study bacterial adhesion to the intestinal epithelium. Lett. Appl. Microbiol. 2009, 49, 695–701. [Google Scholar] [CrossRef]
- Rinkinen, M.; Westermarck, E.; Salminen, S.; Ouwehand, A.C. Absence of host specificity for in vitro adhesion of probiotic lactic acid bacteria to intestinal mucus. Vet. Microbiol. 2003, 97, 55–61. [Google Scholar] [CrossRef]
- He, F.; Ouwehan, A.C.; Hashimoto, H.; Isolauri, E.; Benno, Y.; Salminen, S. Adhesion of Bifidobacterium spp. to human intestinal mucus. Microbiol. Immunol. 2001, 45, 259–262. [Google Scholar] [CrossRef]
- Juntunen, M.; Kirjavainen, P.V.; Ouwehand, A.C.; Salminen, S.J.; Isolauri, E. Adherence of probiotic bacteria to human intestinal mucus in healthy infants and during rotavirus infection. Clin. Diagn. Lab. Immunol. 2001, 8, 293–296. [Google Scholar]
- Martins, F.S.; Silva, A.A.; Vieira, A.T.; Barbosa, F.H.; Arantes, R.M.; Teixeira, M.M.; Nicoli, J.R. Comparative study of Bifidobacterium animalis, Escherichia coli, Lactobacillus casei and Saccharomyces boulardii probiotic properties. Arch. Microbiol. 2009, 191, 623–630. [Google Scholar] [CrossRef]
- Collado, M.C.; Meriluoto, J.; Salminen, S. Role of commercial probiotic strains against human pathogen adhesion to intestinal mucus. Lett. Appl. Microbiol. 2007, 45, 454–460. [Google Scholar] [CrossRef]
- Collado, M.C.; Grzeskowiak, L.; Salminen, S. Probiotic strains and their combination inhibit in vitro adhesion of pathogens to pig intestinal mucosa. Curr. Microbiol. 2007, 55, 260–265. [Google Scholar] [CrossRef]
- Commane, D.M.; Shortt, C.T.; Silvi, S.; Cresci, A.; Hughes, R.M.; Rowland, I.R. Effects of fermentation products of pro- and prebiotics on trans-epithelial electrical resistance in an in vitro model of the colon. Nutr. Cancer 2005, 51, 102–109. [Google Scholar] [CrossRef]
- Lopez, P.; Gueimonde, M.; Margolles, A.; Suarez, A. Distinct Bifidobacterium strains drive different immune responses in vitro. Int. J. Food Microbiol. 2010, 138, 157–165. [Google Scholar] [CrossRef]
- Latvala, S.; Pietila, T.E.; Veckman, V.; Kekkonen, R.A.; Tynkkynen, S.; Korpela, R.; Julkunen, I. Potentially probiotic bacteria induce efficient maturation but differential cytokine production in human monocyte-derived dendritic cells. World J. Gastroenterol. 2008, 14, 5570–5581. [Google Scholar] [CrossRef]
- Matsumoto, M.; Hara, K.; Benno, Y. The influence of the immunostimulation by bacterial cell components derived from altered large intestinal microbiota on probiotic anti-inflammatory benefits. FEMS Immunol. Med. Microbiol. 2007, 49, 387–390. [Google Scholar] [CrossRef]
- Black, F.T. Placebo-Controlled Double-Blind Trial of 4 Lactobacilli Strains (HIP) Used as Prophylactic Agent against Traveller’s Diarrhea (2 Trials); Report by G. Nirnberger; Bioconsult: Vienna, Austria, 1996. [Google Scholar]
- Palaria, A.; Johnson-Kanda, I.; O’Sullivan, D.J. Effect of a synbiotic yogurt on levels of fecal bifidobacteria, clostridia and enterobacteria. Appl. Environ. Microbiol. 2012, 78, 933–940. [Google Scholar] [CrossRef]
- Larsen, C.N.; Nielsen, S.; Kaestel, P.; Brockmann, E.; Bennedsen, M.; Christensen, H.R.; Eskesen, D.C.; Jacobsen, B.L.; Michaelsen, K.F. Dose-response study of probiotic bacteria Bifidobacterium animalis subsp lactis BB-12 and Lactobacillus paracasei subsp. paracasei CRL-341 in healthy young adults. Eur. J. Clin. Nutr. 2006, 60, 1284–1293. [Google Scholar] [CrossRef]
- Matto, J.; Fonden, R.; Tolvanen, T.; Vonwright, A.; Vilpponensalmela, T.; Satokari, R.; Saarela, M. Intestinal survival and persistence of probiotic Lactobacillus and Bifidobacterium strains administered in triple-strain yoghurt. Int. Dairy J. 2006, 16, 1174–1180. [Google Scholar] [CrossRef]
- Alander, M. Effect of galacto-oligosaccharide supplementation on human faecal microflora and on survival and persistence of Bifidobacterium lactis Bb-12 in the gastrointestinal tract. Int. Dairy J. 2001, 11, 817–825. [Google Scholar] [CrossRef]
- Savard, P.; Lamarche, B.; Paradis, M.E.; Thiboutot, H.; Laurin, E.; Roy, D. Impact of Bifidobacterium animalis subsp. lactis BB-12 and, Lactobacillus acidophilus LA-5-containing yoghurt, on fecal bacterial counts of healthy adults. Int. J. Food Microbiol. 2011, 149, 50–57. [Google Scholar] [CrossRef]
- Pitkala, K.H.; Strandberg, T.E.; Finne Soveri, U.H.; Ouwehand, A.C.; Poussa, T.; Salminen, S. Fermented cereal with specific bifidobacteria normalizes bowel movements in elderly nursing home residents. A randomized, controlled trial. J. Nutr. Health Aging 2007, 11, 305–311. [Google Scholar]
- Uchida, K.; Akashi, K.; Kusunoki, I.; Ikeda, T.; Katano, N.; Motoshima, H.; Benno, Y. Effect of fermented milk containing Bifidobacterium lactis BB-12® on stool frequency, defecation, fecal microbiota and safety of excessive ingestion in healthy female students. J. Nutr. Food 2005, 8, 39–51. [Google Scholar]
- Nishida, S.; Gotou, M.; Akutsu, S.; Ono, M.; Hitomi, Y.; Nakamura, T.; Iino, H. Effect of yogurt containing Bifidobacterium lactis BB-12 on improvement of defecation and fecal microflora of healthy female adults. Milk Sci. 2004, 53, 71–80. [Google Scholar]
- Matsumoto, M.; Imai, T.; Hironaka, T.; Kume, H.; Watanabe, M.; Benno, Y. Effect of yoghurt with Bifidobacterium lactis LKM512 in improving fecal microflora and defecation of healthy volunteers. Intest. Microbiol. Mag. 2001, 14, 97–102. [Google Scholar]
- Chouraqui, J.P.; van Egroo, L.D.; Fichot, M.C. Acidified milk formula supplemented with Bifidobacterium lactis: Impact on infant diarrhea in residential care settings. J. Pediatr. Gastroenterol. Nutr. 2004, 38, 288–292. [Google Scholar] [CrossRef]
- Saavedra, J.M.; Bauman, N.A.; Oung, I.; Perman, J.A.; Yolken, R.H. Feeding of Bifidobacterium bifidum and Streptococcus thermophilus to infants in hospital for prevention of diarrhoea and shedding of rotavirus. Lancet 1994, 344, 1046–1049. [Google Scholar] [CrossRef]
- Chatterjee, S.; Kar, P.; Das, T.; Ray, S.; Ganguly, S.; Rajendiran, C.; Mitra, M. Randomised placebo-controlled double blind multicentric trial on efficacy and safety of Lactobacillus acidophilus LA-5® and Bifidobacterium BB-12® for prevention of antibiotic-associated diarrhoea. JAPI 2013, 61, 708–712. [Google Scholar]
- De Vrese, M.; Kristen, H.; Rautenberg, P.; Laue, C.; Schrezenmeir, J. Probiotic lactobacilli and bifidobacteria in a fermented milk product with added fruit preparation reduce antibiotic associated diarrhea and Helicobacter pylori activity. J. Dairy Res. 2011, 78, 396–403. [Google Scholar] [CrossRef]
- Sheu, B.S.; Wu, J.J.; Lo, C.Y.; Wu, H.W.; Chen, J.H.; Lin, Y.S.; Lin, M.D. Impact of supplement with Lactobacillus- and Bifidobacterium-containing yogurt on triple therapy for Helicobacter pylori eradication. Aliment. Pharmacol. Ther. 2002, 16, 1669–1675. [Google Scholar] [CrossRef]
- Wang, K.Y.; Li, S.N.; Liu, C.S.; Perng, D.S.; Su, Y.C.; Wu, D.C.; Jan, C.M.; Lai, C.H.; Wang, T.N.; Wang, W.M. Effects of ingesting Lactobacillus- and Bifidobacterium-containing yogurt in subjects with colonized Helicobacter pylori. Am. J. Clin. Nutr. 2004, 80, 737–741. [Google Scholar]
- Sheu, B.S.; Cheng, H.C.; Kao, A.W.; Wang, S.T.; Yang, Y.J.; Yang, H.B.; Wu, J.J. Pretreatment with Lactobacillus- and Bifidobacterium-containing yogurt can improve the efficacy of quadruple therapy in eradicating residual Helicobacter pylori infection after failed triple therapy. Am. J. Clin. Nutr. 2006, 83, 864–869. [Google Scholar]
- Albers, R.; Antoine, J.M.; Bourdet-Sicard, R.; Calder, P.C.; Gleeson, M.; Lesourd, B.; Samartin, S.; Sanderson, I.R.; van Loo, J.; Vas Dias, F.W.; et al. Markers to measure immunomodulation in human nutrition intervention studies. Br. J. Nutr. 2005, 94, 452–481. [Google Scholar] [CrossRef]
- Burleson, G.R.; Burleson, F.G. Influenza virus host resistance model. Methods 2007, 41, 31–37. [Google Scholar] [CrossRef]
- Rizzardini, G.; Eskesen, D.; Calder, P.C.; Capetti, A.; Jespersen, L.; Clerici, M. Evaluation of the immune benefits of two probiotic strains Bifidobacterium animalis ssp. lactis, BB-12® and Lactobacillus paracasei ssp. paracasei, L. casei 431® in an influenza vaccination model: A randomised, double-blind, placebo-controlled study. Br. J. Nutr. 2012, 107, 876–884. [Google Scholar] [CrossRef]
- Smith, T.J.; Rigassio-Radler, D.; Denmark, R.; Haley, T.; Touger-Decker, R. Effect of Lactobacillus rhamnosus LGG® and Bifidobacterium animalis ssp. lactis BB-12® on health-related quality of life in college students affected by upper respiratory infections. Br. J. Nutr. 2013, 109, 1999–2007. [Google Scholar] [CrossRef]
- Taipale, T.; Pienihakkinen, K.; Isolauri, E.; Larsen, C.; Brockmann, E.; Alanen, P.; Jokela, J.; Soderling, E. Bifidobacterium animalis subsp. lactis BB-12 in reducing the risk of infections in infancy. Br. J. Nutr. 2011, 105, 409–416. [Google Scholar] [CrossRef]
- Rautava, S.; Salminen, S.; Isolauri, E. Specific probiotics in reducing the risk of acute infections in infancy—A randomised, double-blind, placebo-controlled study. Br. J. Nutr. 2009, 101, 1722–1726. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jungersen, M.; Wind, A.; Johansen, E.; Christensen, J.E.; Stuer-Lauridsen, B.; Eskesen, D. The Science behind the Probiotic Strain Bifidobacterium animalis subsp. lactis BB-12®. Microorganisms 2014, 2, 92-110. https://doi.org/10.3390/microorganisms2020092
Jungersen M, Wind A, Johansen E, Christensen JE, Stuer-Lauridsen B, Eskesen D. The Science behind the Probiotic Strain Bifidobacterium animalis subsp. lactis BB-12®. Microorganisms. 2014; 2(2):92-110. https://doi.org/10.3390/microorganisms2020092
Chicago/Turabian StyleJungersen, Mikkel, Anette Wind, Eric Johansen, Jeffrey E. Christensen, Birgitte Stuer-Lauridsen, and Dorte Eskesen. 2014. "The Science behind the Probiotic Strain Bifidobacterium animalis subsp. lactis BB-12®" Microorganisms 2, no. 2: 92-110. https://doi.org/10.3390/microorganisms2020092
APA StyleJungersen, M., Wind, A., Johansen, E., Christensen, J. E., Stuer-Lauridsen, B., & Eskesen, D. (2014). The Science behind the Probiotic Strain Bifidobacterium animalis subsp. lactis BB-12®. Microorganisms, 2(2), 92-110. https://doi.org/10.3390/microorganisms2020092