
Complete Genome Analysis and Antimicrobial Mechanism of Burkholderia gladioli ZBSF BH07 Reveal Its Dual Role in the Biocontrol of Grapevine Diseases and Growth Promotion in Grapevines

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains, Culture Conditions, and Plant Material
2.2. Isolation of Strains and In Vitro Antifungal Activity Assays
2.3. Whole-Genome Sequencing and Assembly
2.4. Gene Prediction and Functional Annotation
2.5. Phylogenetic Tree Construction
2.6. Comparative Genomics Analysis and Mining for Genes Related to Plant-Beneficial Traits
2.7. Morphological, Physiological, and Biochemical Tests
2.8. Measurement of Siderophore Production and Phosphate Solubilization
2.9. Assessment of Biocontrol Activity and Plant Growth Promotion
2.10. Statistical Analysis
3. Results
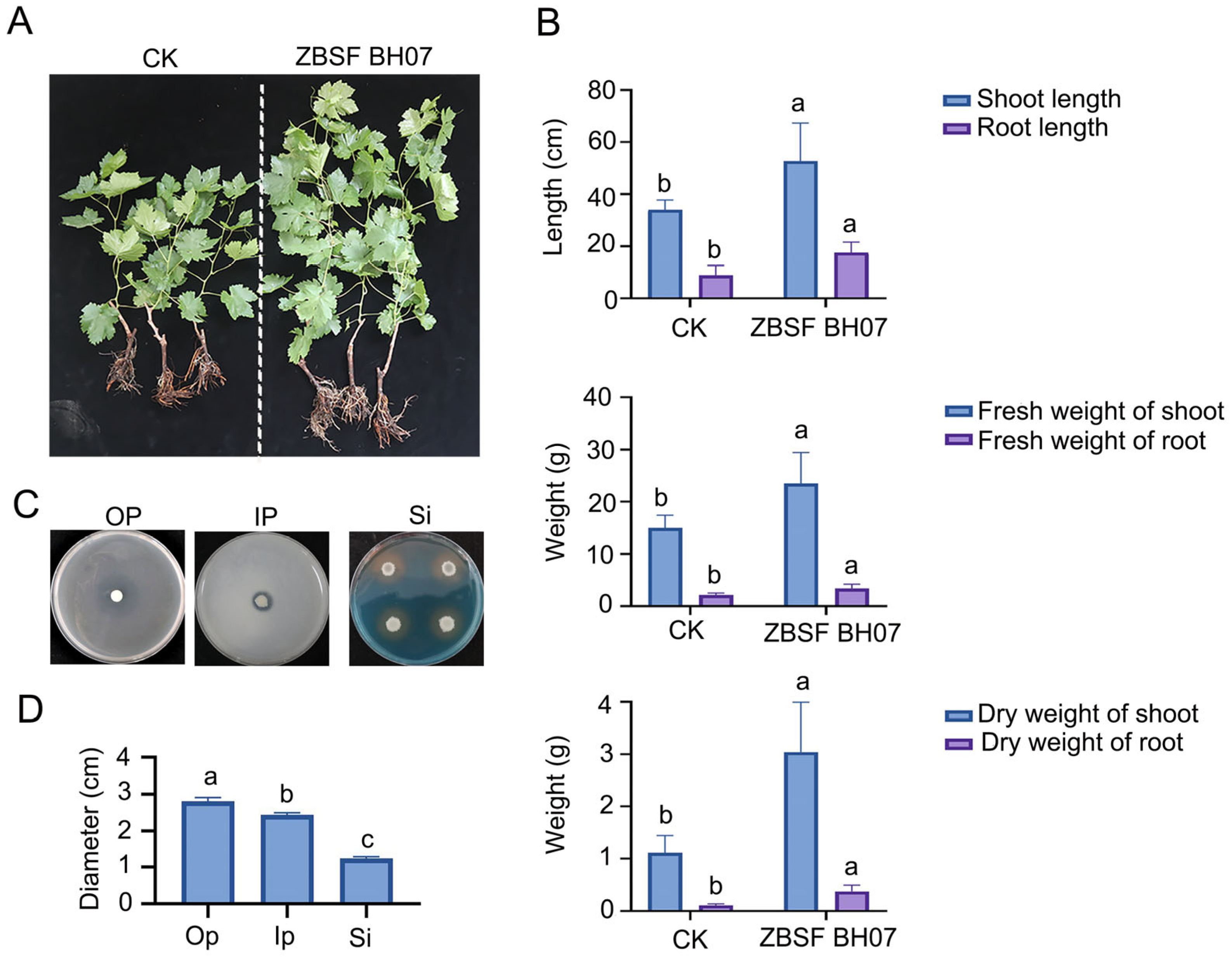
3.1. Biocontrol Effect of Isolate ZBSF BH07 Against Plant Pathogens
3.2. Efficacy of ZBSF BH07 in Controlling Grape White Rot
3.3. Effects of ZBSF BH07 on Plant Growth Promotion
3.4. General Genomic Features of B. gladioli ZBSF BH07
3.5. Morphological Identification and Phylogenetic Analysis of B. gladioli ZBSF BH07
3.6. Comparison of the B. gladioli ZBSF BH07 Genome with Those of Other Completely Sequenced B. gladioli Strains
3.6.1. Comparison of ZBSF BH07 with the Genomic Information of Other B. gladioli
3.6.2. ANI and DDH Analysis
3.6.3. Genetic Basis for Promoting Plant Growth
3.7. Genes/Gene Cluster for Antibiotic Synthesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of Plant Growth Promoting Rhizobacteria in Agricultural Sustainability-A Review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [PubMed]
- Burkholder, W.H. Three bacterial plant pathogens: Phytomonas earyophylli sp. n., Phytomonas alliicola sp. n., and Phytomonas manihotis (Arthaud-Berthet et Sondar) Viégas. Phytopathology 1942, 32, 141–149. [Google Scholar]
- Yabuuchi, E.; Kosako, Y.; Oyaizu, H.; Yano, I.; Hotta, H.; Hashimoto, Y.; Arakawa, M. Proposal of Burkholderia gen. nov. and transfer of seven species of the genus Pseudomonas homology group II to the new genus, with the type species Burkholderia cepacia (Palleroni and Holmes 1981) comb. nov. Microbiol. Immunol. 1992, 36, 1251–1275. [Google Scholar] [CrossRef] [PubMed]
- Coenye, T.; Vandamme, P. Diversity and significance of Burkholderia species occupying diverse ecological niches. Environ. Microbiol. 2003, 5, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Ait Barka, E.; Nowak, J.; Clément, C. Enhancement of chilling resistance of inoculated grapevine plantlets with a plant growth-promoting rhizobacterium, Burkholderia phytofirmans strain PsJN. Appl. Environ. Microbiol. 2006, 72, 7246–7252. [Google Scholar] [CrossRef] [PubMed]
- An, C.; Ma, S.; Liu, C.; Ding, H.; Xue, W. Burkholderia ambifaria XN08: A plant growth-promoting endophytic bacterium with biocontrol potential against sharp eyespot in wheat. Front. Microbiol. 2022, 13, 906724. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Luo, W.Z.; Zhang, D.D.; Li, R.; Kong, Z.Q.; Song, J.; Dai, X.F.; Alkan, N.; Chen, J.Y. Insights into the Biocontrol Function of a Burkholderia gladioli Strain against Botrytis cinerea. Microbiol. Spectr. 2023, 11, e0480522. [Google Scholar] [CrossRef] [PubMed]
- Valvano, M.A.; Keith, K.E.; Cardona, S.T. Survival and persistence of opportunistic Burkholderia species in host cells. Curr. Opin. Microbiol. 2005, 8, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Lee, D.; Lee, S.B.; Lim, G.H.; Kim, S.W.; Kim, T.J.; Park, D.S.; Seo, Y.S. Understanding Burkholderia glumae BGR1 Virulence through the Application of Toxoflavin-Degrading Enzyme, TxeA. Plants 2023, 12, 3934. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Park, J.; Choi, O.; Kim, J.; Seo, Y.S. Investigation of quorum sensing-dependent gene expression in Burkholderia gladioli BSR3 through RNA-seq analyses. J. Microbiol. Biotechnol. 2014, 24, 1609–1621. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Deng, H.; Wang, Y.; Zhang, C.; Zhu, J. Detection methods and control measures of Burkholderia gladioli and its toxins: A review. J. Food Sci. 2025, 90, e17668. [Google Scholar] [CrossRef] [PubMed]
- Bach, E.; Passaglia, L.M.P.; Jiao, J.; Gross, H. Burkholderia in the genomic era: From taxonomy to the discovery of new antimicrobial secondary metabolites. Crit. Rev. Microbiol. 2022, 48, 121–160. [Google Scholar] [CrossRef] [PubMed]
- Tian, P.; Huang, Q.; Peng, W.; Guo, X.; Sun, L.; Jian, W.; Xie, C.; Yang, X. Complete genome sequence of Burkholderia gladioli BK04 with biocontrol potential against cotton verticillium wilt (Verticillium dahliae). Mol. Plant Microbe Interact. 2022, 35, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- Cui, G.; Yin, K.; Lin, N.; Liang, M.; Huang, C.; Chang, C.; Xi, P.; Deng, Y.Z. Burkholderia gladioli CGB10: A novel strain biocontrolling the sugarcane smut disease. Microorganisms 2020, 8, 1943. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.T.; Lee, C.C.; Leu, W.M.; Wu, J.J.; Huang, Y.C.; Meng, M. Fungicidal activity of volatile organic compounds emitted by Burkholderia gladioli strain BBB-01. Molecules 2021, 26, 745. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Carlson, R.; Tharpe, W.; Schell, M.A. Characterization of genes involved in biosynthesis of a novel antibiotic from Burkholderia cepacia BC11 and their role in biological control of Rhizoctonia solani. Appl. Environ. Microbiol. 1998, 64, 3939–3947. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Jenner, M.; Masschelein, J.; Jones, C.; Bull, M.J.; Harris, S.R.; Hartkoorn, R.C.; Vocat, A.; Romero-Canelon, I.; Coupland, P.; et al. Discovery and biosynthesis of gladiolin: A Burkholderia gladioli antibiotic with promising activity against Mycobacterium tuberculosis. J. Am. Chem. Soc. 2017, 139, 7974–7981. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, A.; Yadav, S.K.; Jha, G. Glycosyltransferase-Like Toxin of Burkholderia gladioli Strain NGJ1 Is a Potent Antifungal Protein with Potential for Control of Sheath Blight Disease in Rice. Phytopathology 2025, 115, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Penttinen, P.; Zhang, X.; Ao, X.; Liu, M.; Yu, X.; Chen, Q. Maize rhizosphere in Sichuan, China, hosts plant growth promoting Burkholderia cepacia with phosphate solubilizing and antifungal abilities. Microbiol. Res. 2014, 169, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.; Khan, S.; Iqbal, S.; Mirza, M.S.; Khan, Q.M. Inoculation method affects colonization and activity of Burkholderia phytofirmans PsJN during phytoremediation of diesel-contaminated soil. Int. Biodeterior. Biodegrad. 2013, 85, 331–336. [Google Scholar] [CrossRef]
- Baghel, V.; Thakur, J.K.; Yadav, S.S.; Manna, M.C.; Mandal, A.; Shirale, A.O.; Patra, A.K. Phosphorus and potassium solubilization from rock minerals by endophytic Burkholderia sp. strain FDN2–1 in soil and shift in diversity of bacterial endophytes of corn root tissue with crop growth stage. Geomicrobiol. J. 2020, 37, 550–563. [Google Scholar] [CrossRef]
- Malviya, M.K.; Li, C.N.; Solanki, M.K.; Singh, R.K.; Htun, R.; Singh, P.; Verma, K.K.; Yang, L.T.; Li, Y.R. Comparative analysis of sugarcane root transcriptome in response to the plant growth-promoting Burkholderia anthina MYSP113. PLoS ONE 2020, 15, e0231206. [Google Scholar] [CrossRef] [PubMed]
- Sabir, A.; Yazici, M.A.; Kara, Z.; Sahin, F. Growth and mineral acquisition response of grapevine rootstocks (Vitis spp.) to inoculation with different strains of plant growth-promoting rhizobacteria (PGPR). J. Sci. Food Agric. 2012, 92, 2148–2153. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Lin, H.; Shan, Y.; Song, J.; Zhang, D.D.; Dai, X.F.; Han, D.; Chen, J.Y. The potential of Burkholderia gladioli KRS027 in plant growth promotion and biocontrol against Verticillium dahliae revealed by dual transcriptome of pathogen and host. Microbiol. Res. 2024, 287, 127836. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Li, T.; Jiang, X.; Tang, X.; Zhang, J.; Yuan, L.; Wei, Y. Suppression of Grape White Rot Caused by Coniella vitis Using the Potential Biocontrol Agent Bacillus velezensis GSBZ09. Pathogens 2022, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Jiang, H.; Jiang, X.; Li, T.; Lu, P.; Yin, X.; Wei, Y. Comparative genomic and functional analyses of Paenibacillus peoriae ZBSF16 with biocontrol potential against grapevine diseases, provide insights into its genes related to plant growth-promoting and biocontrol mechanisms. Front. Microbiol. 2002, 13, 975344. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, A.; Au, K.F. PacBio sequencing and its applications. Genom. Proteom. Bioinform. 2015, 13, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Larralde, M. Pyrodigal: Python bindings and interface to Prodigal, an efficient method for gene prediction in prokaryotes. J. Open Source Softw. 2022, 7, 4296. [Google Scholar] [CrossRef]
- Chan, P.P.; Lin, B.Y.; Mak, A.J.; Lowe, T.M. tRNAscan-SE 2.0: Improved detection and functional classification of transfer RNA genes. Nucleic Acids Res. 2021, 49, 9077–9096. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Glaeser, S.P.; Kämpfer, P. Multilocus sequence analysis (MLSA) in prokaryotic taxonomy. Syst. Appl. Microbiol. 2015, 38, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Darling, A.C.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [PubMed]
- Alikhan, N.F.; Petty, N.K.; Zakour, N.L.; Ben Beatson, S.A. BLAST ring image generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Acharya, V.; Mukhia, S.; Singh, D.; Kumar, S. Complete genome sequence of Pseudomonas frederiksbergensis ERDD5: 01 revealed genetic bases for survivability at high altitude ecosystem and bioprospection potential. Genomics 2019, 111, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Vader, L.; Szenei, J.; Reitz, Z.L.; Augustijn, H.E.; Cediel-Becerra, J.D.D.; de Crécy-Lagard, V.; Koetsier, R.A.; Williams, S.E.; et al. antiSMASH 8.0: Extended gene cluster detection capabilities and analyses of chemistry, enzymology, and regulation. Nucleic Acids Res. 2025, 53, W32–W38. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, D.D.; Li, R.; Zhu, H.; Song, J.; Kong, Z.Q.; Dai, X.F.; Han, D.F.; Chen, J.Y. Genome Sequence Resource of Burkholderia gladioli KRS027: A Strain with Antifungal Activity Against Plant Pathogens. PhytoFrontiers 2023, 3, 874–878. [Google Scholar] [CrossRef]
- Seo, Y.S.; Lim, J.; Choi, B.S.; Kim, H.; Goo, E.; Lee, B.; Lim, J.S.; Choi, I.Y.; Moon, J.S.; Kim, J.; et al. Complete genome sequence of Burkholderia gladioli BSR3. J. Bacteriol. 2011, 193, 3149. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Wang, Z.; Wan, J.; Qi, T.; Zou, L. Burkholderia gladioli strain KJ-34 exhibits broad-spectrum antifungal activity. Front. Plant Sci. 2023, 14, 1097044. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, F.X.; Rossi, M.J.; Glick, B.R. Ethylene and 1-aminocyclopropane-1-carboxylate (ACC) in plant-bacterial interactions. Front. Plant Sci. 2018, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Zhuang, Y.; Zhang, L.; Li, S.; Ruan, S.; Zhang, Q. Preferred hierarchical control strategy of phosphorus from nonPoint source pollution at regional scale. Environ. Sci. Pollut. Res. Int. 2021, 28, 60111–60121. [Google Scholar] [CrossRef] [PubMed]
- de Andrade, L.A.; Santos, C.H.B.; Frezarin, E.T.; Sales, L.R.; Rigobelo, E.C. Plant growth-promoting rhizobacteria for sustainable agricultural production. Microorganisms 2023, 11, 1088. [Google Scholar] [CrossRef] [PubMed]
- de Werra, P.; Péchy-Tarr, M.; Keel, C.; Maurhofer, M. Role of gluconic acid production in the regulation of biocontrol traits of Pseudomonas fluorescens CHA0. Appl. Environ. Microbiol. 2009, 75, 4162–4174. [Google Scholar] [CrossRef] [PubMed]
- Nikata, T.; Sakai, Y.; Shibat, K.; Kato, J.; Kuroda, A.; Ohtake, H. Molecular analysis of the phosphate-specific transport (pst) operon of Pseudomonas aeruginosa. Mol. Gen. Genet. 1996, 250, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Chan, F.Y.; Torriani, A.J. PstB protein of the phosphate-specific transport system of Escherichia coli is an ATPase. J. Bacteriol. 1996, 178, 3974–3977. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Cai, B. Phosphate-solubilizing bacteria: Advances in their physiology, molecular mechanisms and microbial community effects. Microorganisms 2023, 11, 2904. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Jiao, Z.; Hale, L.; Wu, W.; Gao, Y. Disruption of gene pqqA or pqqB reduces plant growth promotion activity and biocontrol of grown gall disease by Rahnella aquatilis HX2. PLoS ONE 2014, 9, e115010. [Google Scholar] [CrossRef]
- Estrada-De Los Santos, P.; Bustillos-Cristales, R.; Caballero-Mellado, J. Burkholderia, a genus rich in plant-associated nitrogen fixers with wide environmental and geographic distribution. Appl. Environ. Microbiol. 2001, 67, 2790–2798. [Google Scholar] [CrossRef] [PubMed]
- Perin, L.; Martínez-Aguilar, L.; Paredes-Valdez, G.; Baldani, J.I.; Santos, P.E.D.L.; Reis, V.M.; Caballero-Mellado, J. Burkholderia silvatlantica sp. nov., a diazotrophic bacterium associated with sugar cane and maize. Int. J. Syst. Evol. Microbiol. 2006, 56, 1931–1937. [Google Scholar] [CrossRef] [PubMed]
- Estrada, P.; Mavingui, P.; Cournoyer, B.; Fontaine, F.; Balandreau, J.; Caballero-Mellado, J. A N2-fixing endophytic Burkholderia sp. associated with maize plants cultivated in Mexico. Can. J. Microbiol. 2002, 48, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Geudens, N.; Kieu, N.P.; Sinnaeve, D.; Ongena, M.; Martins, J.C.; Höfte, M. Biosynthesis, chemical structure, and structure-activity relationship of orfamide lipopeptides produced by Pseudomonas protegens and related species. Front. Microbiol. 2016, 7, 382. [Google Scholar] [CrossRef] [PubMed]
- D’aes, J.; Kieu, N.P.; Léclère, V.; Tokarski, C.; Olorunleke, F.E.; De Maeyer, K.; Jacques, P.; Höfte, M.; Ongena, M. To settle or to move? The interplay between two classes of cyclic lipopeptides in the biocontrol strain Pseudomonas CMR12a. Environ. Microbiol. 2014, 16, 2282–2300. [Google Scholar] [CrossRef] [PubMed]
- Hermenau, R.; Mehl, J.L.; Ishida, K.; Dose, B.; Pidot, S.J.; Stinear, T.P.; Hertweck, C. Genomics-driven discovery of nO-donating diazeniumdiolate siderophores in diverse plant-associated bacteria. Angew. Chem. Int. Ed. Engl. 2019, 58, 13024–13029. [Google Scholar] [CrossRef] [PubMed]
- Dose, B.; Niehs, S.P.; Scherlach, K.; Shahda, S.; Flórez, L.V.; Kaltenpoth, M.; Hertweck, C. Biosynthesis of sinapigladioside, an antifungal isothiocyanate from Burkholderia Symbionts. ChemBioChem 2021, 22, 1920–1924. [Google Scholar] [CrossRef] [PubMed]
- Petrova, Y.D.; Zhao, J.; Webster, G.; Mullins, A.J.; Williams, K.; Alswat, A.S.; Challis, G.L.; Bailey, A.M.; Mahenthiralingam, E. Cloning and expression of Burkholderia polyyne biosynthetic gene clusters in Paraburkholderia hosts provides a strategy for biopesticide development. Microb. Biotechnol. 2022, 15, 2547–2561. [Google Scholar] [CrossRef] [PubMed]
- Flórez, L.V.; Scherlach, K.; Gaube, P.; Ross, C.; Sitte, E.; Hermes, C.; Rodrigues, A.; Hertweck, C.; Kaltenpoth, M. Antibiotic-producing symbionts dynamically transition between plant pathogenicity and insect-defensive mutualism. Nat. Commun. 2017, 8, 15172. [Google Scholar] [CrossRef] [PubMed]
- Dose, B.; Niehs, S.P.; Scherlach, K.; Flórez, L.V.; Kaltenpoth, M.; Hertweck, C. Unexpected bacterial origin of the antibiotic icosalide: Two-tailed depsipeptide assembly in multifarious Burkholderia Symbionts. ACS Chem. Biol. 2018, 13, 2414–2420. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Bai, X.; Sun, T.; Wang, X.; Zhang, Y.; Bian, X.; Zhou, H. The genomic-driven discovery of glutarimide-containing derivatives from Burkholderia gladioli. Molecules 2023, 28, 6937. [Google Scholar] [CrossRef] [PubMed]
- Bode, H.B.; Brachmann, A.O.; Jadhav, K.B.; Seyfarth, L.; Dauth, C.; Fuchs, S.W.; Kaiser, M.; Waterfield, N.R.; Sack, H.; Heinemann, S.H.; et al. Structure elucidation and activity of kolossin A, the D-/L-pentadecapeptide product of a giant nonribosomal peptide synthetase. Angew. Chem. Int. Ed. Engl. 2015, 54, 10352–10355. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Chen, J.; Chen, W.; Wang, Y. Bongkrekic acid and Burkholderia gladioli pathovar cocovenenans: Formidable foe and ascending threat to food safety. Foods 2023, 12, 3926. [Google Scholar] [CrossRef] [PubMed]
- Choi, O.; Lee, Y.; Kang, B.; Kim, J. Toxoflavin contamination in rice samples from rice processing complexes in South Korea. Int. J. Food Microbiol. 2023, 385, 110014. [Google Scholar] [CrossRef] [PubMed]
- Tien, D.T.K.; Giang, V.Q.; Luc, P.V.; Vy, K.T.T.; Thong, H.T.; Hau, L.V.; Lefeuvre, P.; Nga, N.T.T. Complete and nearly complete genome sequences of seven strains of Burkholderia glumae and B. gladioli isolated from rice panicles in Vietnam. Microbiol. Resour. Announc. 2025, 14, e0038825. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Liu, D.; Lin, R.; Zhu, Y.; Zhu, P.; Jiang, X.; Mao, J.; Cao, Y.; Peng, J.; Zhao, T.; et al. Bongkrekic Acid and Its Novel Isomers: Separation, Identification, and Determination in Food Matrices. Toxins 2025, 17, 223. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Features | B. gladioli ZBSF BH07 | B. gladioli CGB10 | B. gladioli KRS027 | B. gladioli BK04 | B. gladioli BBB-01 | B. gladioli BSR3 |
|---|---|---|---|---|---|---|
| Biosample | SAMN48577572 | SAMN15158960 | SAMN34105703 | SAMN28690464 | SAMN17301201 | SAMN02603164 |
| Chromosome Number | 2 | 2 | 2 | 2 | 2 | 2 |
| Plasmid Number | 1 | 1 | 1 | 0 | 1 | 4 |
| Size (bp) | 8,558,021 | 8,423,203 | 8,257,358 | 8,039,389 | 8,201,484 | 9,052,299 |
| GC content (%) | 68.06 | 68.13 | 68.67 | 68.24 | 68.19 | 67.40 |
| Total genes | 7431 | 7462 | 8844 | 6928 | 7090 | 7757 |
| Predicted protein-coding genes | 7262 | 6981 | 8781 | 6768 | 6941 | 7410 |
| Ribosomal RNA | 15 | 15 | 15 | 15 | 15 | 15 |
| tRNA | 68 | 67 | 66 | 67 | 66 | 68 |
| CRISPR | 25 | 6 | / | / | 6 | / |
| Pseudogene | 84 | 395 | / | 74 | 64 | 264 |
| Origin | grape rhizosphere soil | sugarcane leaves | Cotton rhizosphere soil | Catalpa ovata rhizosphere soil | rice shoot | Rice |
| Reference | This study | [16] | [42] | [3] | [17] | [43] |
| Gene Name | Product | ZBSF BH07 | CGB10 | KRS027 | BK04 | BBB01 | BSR3 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ProteinID | ProteinID | Homology (%) | ProteinID | Homology (%) | ProteinID | Homology (%) | ProteinID | Homology (%) | ProteinID | Homology (%) | ||
| Phosphate solubilization genes | ||||||||||||
| pstS | phosphate ABC transporter substrate-binding | WP_013697368.1 | WP_013697368.1 | 100.00 | NA | 99.66 | WP_250804138.1 | 99.42 | WP_013697368.1 | 100.00 | WP_013697368.1 | 100.00 |
| pstC | phosphate ABC transporter permease PstC | WP_013697369.1 | WP_013697369.1 | 100.00 | NA | 100.00 | WP_013697369.1 | 100.00 | WP_013697369.1 | 100.00 | WP_013697369.1 | 100 |
| pstA | phosphate ABC transporter permease PstA | WP_105851495.1 | WP_096749337.1 | 99.33 | NA | 99.66 | WP_036034067.1 | 99.66 | WP_036034067.1 | 99.66 | WP_013697370.1 | 99.33 |
| pstB | phosphate ABC transporter ATP-binding protein PstB | WP_013697371.1 | WP_013697371.1 | 100.00 | NA | 100.00 | WP_105826707.1 | 99.29 | WP_013697371.1 | 100.00 | WP_013697371.1 | 100.00 |
| phoU | phosphate signaling complex protein PhoU | WP_013697372.1 | WP_013697372.1 | 100.00 | NA | 100.00 | WP_013697372.1 | 100.00 | WP_013697372.1 | 100.00 | WP_013697372.1 | 100.00 |
| phoB | phosphate regulon transcriptional regulator PhoB | WP_025099650.1 | WP_025099650.1 | 100.00 | NA | 100.00 | WP_025099650.1 | 100.00 | WP_025099650.1 | 100.00 | WP_013697373.1 | 99.57 |
| phoR | phosphate regulon sensor histidine kinase PhoR | WP_036052035.1 | WP_036052035.1 | 100.00 | NA | 99.69 | WP_036052035.1 | 100.00 | WP_036052035.1 | 100.00 | WP_013697374.1 | 99.54 |
| inorganic phosphate transporter | WP_017921247.1 | WP_017921247.1 | 100.00 | NA | 99.70 | WP_017921247.1 | 100.00 | WP_017921247.1 | 100.00 | WP_013698297.1 | 99.70 | |
| gcd | glucose/quinate/shikimate family membrane-bound PQQ-dependent dehydrogenase | WP_036056478.1 | WP_096751037.1 | 99.88 | NA | 99.88 | WP_036056478.1 | 100.00 | WP_186039311.1 | 99.76 | WP_013690155.1 | 99.76 |
| pqqA | pyrroloquinoline quinone precursor peptide PqqA | WP_013691668.1 | WP_013691668.1 | 100.00 | NA | 100.00 | WP_013691668.1 | 100.00 | WP_013691668.1 | 100.00 | WP_013691668.1 | 100.00 |
| pqqB | pyrroloquinoline quinone biosynthesis protein PqqB | WP_036028557.1 | WP_046574788.1 | 99.34 | NA | 99.34 | WP_013691667.1 | 99.67 | WP_201446796.1 | 99.34 | WP_013691667.1 | 99.67 |
| pqqC | pyrroloquinoline-quinone synthase PqqC | WP_145756981.1 | WP_145756981.1 | 100.00 | NA | 98.79 | WP_250805862.1 | 99.20 | WP_060001136.1 | 99.60 | WP_043217028.1 | 98.80 |
| pqqD | pyrroloquinoline quinone biosynthesis peptide chaperone PqqD | WP_025098424.1 | WP_025098424.1 | 100.00 | NA | 100.00 | WP_025098424.1 | 100.00 | WP_025098424.1 | 100.00 | WP_013691665.1 | 98.90 |
| pqqE | pyrroloquinoline quinone biosynthesis protein PqqE | WP_186086773.1 | WP_025098425.1 | 99.75 | NA | 99.25 | WP_025098425.1 | 99.75 | WP_105851184.1 | 99.25 | WP_013691664.1 | 98.74 |
| phnX | phosphonoacetaldehyde hydrolase | WP_431018100.1 | WP_260865992.1 | 99.58 | NA | 99.07 | WP_250805634.1 | 98.95 | WP_186020012.1 | 98.60 | WP_013691467.1 | 98.25 |
| phnA | phosphonoacetate hydrolase | WP_431018012.1 | WP_186036997.1 | 99.31 | NA | 98.71 | WP_250804705.1 | 98.17 | WP_201446705.1 | 98.17 | WP_013689373.1 | 98.40 |
| phnV | 2-aminoethylphosphonate ABC transport system, membrane component PhnV | WP_017920137.1 | WP_046582114.1 | 99.65 | NA | 99.30 | WP_017920137.1 | 100.00 | WP_036054457.1 | 99.65 | WP_013689372.1 | 98.95 |
| phnU | 2-aminoethylphosphonate ABC transporter permease subunit | WP_186104004.1 | WP_043217526.1 | 99.68 | NA | 99.15 | WP_186078140.1 | 99.04 | WP_043217526.1 | 99.68 | WP_013689371.1 | 98.39 |
| phnT | 2-aminoethylphosphonate ABC transport system ATP-binding subunit PhnT | WP_036054458.1 | WP_036054458.1 | 100.00 | NA | 100.00 | WP_036054458.1 | 100.00 | WP_036054458.1 | 100.00 | WP_013689370.1 | 99.46 |
| phnS | 2-aminoethylphosphonate ABC transporter substrate-binding protein | WP_017920134.1 | WP_017920134.1 | 100.00 | NA | 100.00 | WP_250804704.1 | 99.72 | WP_017920134.1 | 100.00 | WP_013689369.1 | 99.17 |
| Indole-3-acetic acid biosynthesis genes | ||||||||||||
| trpC | indole-3-glycerol phosphate synthase TrpC | WP_046581210.1 | WP_096749797.1 | 98.85 | NA | 98.85 | WP_096749797.1 | 98.85 | WP_046581210.1 | 100.00 | WP_013696524.1 | 98.47 |
| trpD | anthranilate phosphoribosyltransferase | WP_025100550.1 | WP_025100550.1 | 100.00 | NA | 99.42 | WP_250803631.1 | 99.42 | WP_025100550.1 | 100.00 | WP_013696525.1 | 99.42 |
| anthranilate synthase component II | WP_013696526.1 | WP_013696526.1 | 100.00 | NA | 100.00 | WP_013696526.1 | 100.00 | WP_025100549.1 | 99.49 | WP_013696526.1 | 100.00 | |
| trpE | anthranilate synthase component I | WP_013696527.1 | WP_013696527.1 | 100.00 | NA | 100.00 | WP_013696527.1 | 100.00 | WP_013696527.1 | 100.00 | WP_013696527.1 | 100.00 |
| trpB | tryptophan synthase subunit beta | WP_013690578.1 | WP_013690578.1 | 100.00 | NA | 100.00 | WP_186236507.1 | 99.75 | WP_013690578.1 | 100.00 | WP_013690578.1 | 100.00 |
| trpA | tryptophan synthase subunit alpha | WP_046580979.1 | WP_036055918.1 | 99.26 | NA | 99.26 | WP_036055918.1 | 99.26 | WP_201446989.1 | 98.89 | WP_013690576.1 | 97.78 |
| trpS | tryptophan--tRNA ligase | WP_046582086.1 | WP_046582086.1 | 100.00 | NA | 99.44 | WP_105857686.1 | 99.16 | WP_046582086.1 | 100.00 | WP_013689344.1 | 99.44 |
| iaaH | indoleacetamide hydrolase | WP_160294375.1 | WP_186271478.1 | 99.23 | NA | NA | NA | NA | NA | NA | NA | NA |
| aroC | chorismate synthase | WP_036051822.1 | WP_013697745.1 | 99.73 | NA | 99.18 | WP_250804326.1 | 99.43 | WP_013697745.1 | 99.43 | WP_013697745.1 | 99.43 |
| aroA | 3-phosphoshikimate 1-carboxyvinyltransferase | WP_105859154.1 | WP_036052403.1 | 99.31 | NA | NA | WP_145757446.1 | 99.54 | WP_036052403.1 | 99.31 | WP_013696997.1 | 99.08 |
| shikimate kinase | WP_047837134.1 | WP_080757405.1 | 99.46 | NA | 99.46 | WP_047837134.1 | 100.00 | WP_047837134.1 | 100.00 | WP_047837134.1 | 100.00 | |
| aroE | shikimate dehydrogenase | WP_046576987.1 | WP_186049681.1 | 99.32 | NA | 99.32 | WP_059999883.1 | 99.66 | WP_186049681.1 | 99.32 | WP_013696600.1 | 97.60 |
| aroQ | type II 3-dehydroquinate dehydratase | WP_013696594.1 | WP_013696594.1 | 100.00 | NA | 100.00 | WP_013696594.1 | 100.00 | WP_013696594.1 | 100.00 | WP_013696594.1 | 100.00 |
| aroB | 3-dehydroquinate synthase | WP_017921100.1 | WP_166915270.1 | 99.72 | NA | 99.72 | WP_250803588.1 | 99.72 | WP_017921100.1 | 100.00 | WP_013696404.1 | 99.72 |
| aroG | 3-deoxy-7-phosphoheptulonate synthase AroG | WP_013696710.1 | WP_036048186.1 | 100.00 | NA | 99.72 | WP_036048186.1 | 100.00 | WP_105829091.1 | 99.72 | WP_013696710.1 | 100.00 |
| Cluster | Type | From | To | Size (bp) | Most Similar Known Cluster | Similarity Confidence |
|---|---|---|---|---|---|---|
| Chromosome 1 | ||||||
| Cluster1 | terpene-precursor | 541,153 | 562,151 | 20,998 | ||
| Cluster2 | T1PKS | 741,213 | 788,865 | 47,652 | ||
| Cluster3 | NRPS-like, NRPS | 1,524,680 | 1,579,818 | 55,138 | sulfazecin | High |
| Cluster4 | RiPP-like, NRPS | 1,606,244 | 1,670,249 | 64,005 | ||
| Cluster5 | isocyanide | 2,037,150 | 2,078,985 | 41,835 | sinapigladioside | High |
| Cluster6 | polyyne | 2,114,617 | 2,141,003 | 26,386 | caryoynencin | High |
| Cluster7 | NRPS, T1PKS | 2,248,353 | 2,388,098 | 139,745 | orfamide B | Low |
| Cluster8 | terpene | 2,644,052 | 2,664,891 | 20,839 | ||
| Chromosome 2 | ||||||
| Cluster1 | redox-cofactor | 37,427 | 59,647 | 22,220 | ||
| Cluster2 | terpene | 138,663 | 159,733 | 21,070 | ||
| Cluster3 | transAT-PKS | 554,651 | 640,680 | 86,029 | gladiostatinA | High |
| Cluster4 | terpene-precursor | 1,125,693 | 1,146,577 | 20,884 | ||
| Cluster5 | NRP-metallophore, NRPS | 1,693,445 | 1,777,734 | 84,289 | plantaribactin | High |
| Cluster6 | NRPS | 1,798,931 | 1,855,874 | 56,943 | ||
| Cluster7 | hydrogen-cyanide | 1,887,604 | 1,900,630 | 13,026 | ||
| Cluster8 | RiPP-like | 1,939,225 | 1,950,241 | 11,016 | ||
| Cluster9 | phosphonate | 2,212,326 | 2,246,799 | 34,473 | ||
| Cluster10 | hserlactone | 2,510,928 | 2,531,542 | 20,614 | ||
| Cluster11 | NRPS | 2,539,442 | 2,602,838 | 63,396 | kolossin | High |
| Cluster12 | transAT-PKS, PKS-like, T3PKS | 2,804,582 | 2,944,789 | 140,207 | lagriene | High |
| Cluster13 | terpene, betalactone, RiPP-like | 3,602,604 | 3,644,478 | 41,874 | barbamide | Low |
| Cluster14 | NRPS | 3,775,254 | 3,830,304 | 55,050 | icosalide A/icosalide B | High |
| Cluster15 | ectoine | 3,831,482 | 3,841,868 | 10,386 | ||
| Cluster16 | terpene | 3,872,495 | 3,893,547 | 21,052 | ||
| Cluster17 | terpene | 4,008,965 | 4,032,489 | 23,524 | ||
| Plasmid | ||||||
| Cluster1 | NRPS, T1PKS | 170,127 | 222,396 | 52,269 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, X.; Wang, C.; Yuan, L.; Wei, Y.; Li, T.; Liu, Q.; Han, X.; Wu, X.; Wang, C.; Jiang, X. Complete Genome Analysis and Antimicrobial Mechanism of Burkholderia gladioli ZBSF BH07 Reveal Its Dual Role in the Biocontrol of Grapevine Diseases and Growth Promotion in Grapevines. Microorganisms 2025, 13, 1756. https://doi.org/10.3390/microorganisms13081756
Yin X, Wang C, Yuan L, Wei Y, Li T, Liu Q, Han X, Wu X, Wang C, Jiang X. Complete Genome Analysis and Antimicrobial Mechanism of Burkholderia gladioli ZBSF BH07 Reveal Its Dual Role in the Biocontrol of Grapevine Diseases and Growth Promotion in Grapevines. Microorganisms. 2025; 13(8):1756. https://doi.org/10.3390/microorganisms13081756
Chicago/Turabian StyleYin, Xiangtian, Chundong Wang, Lifang Yuan, Yanfeng Wei, Tinggang Li, Qibao Liu, Xing Han, Xinying Wu, Chaoping Wang, and Xilong Jiang. 2025. "Complete Genome Analysis and Antimicrobial Mechanism of Burkholderia gladioli ZBSF BH07 Reveal Its Dual Role in the Biocontrol of Grapevine Diseases and Growth Promotion in Grapevines" Microorganisms 13, no. 8: 1756. https://doi.org/10.3390/microorganisms13081756
APA StyleYin, X., Wang, C., Yuan, L., Wei, Y., Li, T., Liu, Q., Han, X., Wu, X., Wang, C., & Jiang, X. (2025). Complete Genome Analysis and Antimicrobial Mechanism of Burkholderia gladioli ZBSF BH07 Reveal Its Dual Role in the Biocontrol of Grapevine Diseases and Growth Promotion in Grapevines. Microorganisms, 13(8), 1756. https://doi.org/10.3390/microorganisms13081756