
Bacillus amyloliquefaciens SQ-2 and Biochar: A Promising Combination for Enhancing Rice Growth in Pb/Al-Contaminated Acidic Soils


Abstract
1. Introduction
2. Materials and Methods
2.1. Strain, Medium, Soil, and Biochar
2.2. Determination of Growth-Promoting Characteristics of SQ-2
2.2.1. Detection of Phosphorus Solubilization Ability of SQ-2
2.2.2. Detection of Potassium-Solubilizing Ability of SQ-2
2.3. Rice Soil Culture Experiment
2.4. Characterization of SQ-2–Biochar Adsorption of Pb2+ and Al3+
2.5. Transcriptome Analysis of SQ-2 Adsorbed Pb2+ and Al3+
2.6. Data Analysis
3. Results
3.1. Phosphorus/Potassium-Solubilizing Capability of B. amyloliquefaciens SQ-2
3.2. SQ-2–Biochar Promotes Rice Growth in Pb/Al Acidic Soil
3.3. Mechanisms of SQ-2–Biochar in Promoting Rice Growth in Pb/Al Acidic Soil
3.3.1. The Effects of SQ-2–Biochar on the Physiological Properties of Rice in Pb/Al Acidic Soil
3.3.2. The Effects of SQ-2–Biochar on the Rhizosphere Soil Nutrients of Rice in Pb/Al Acidic Soil
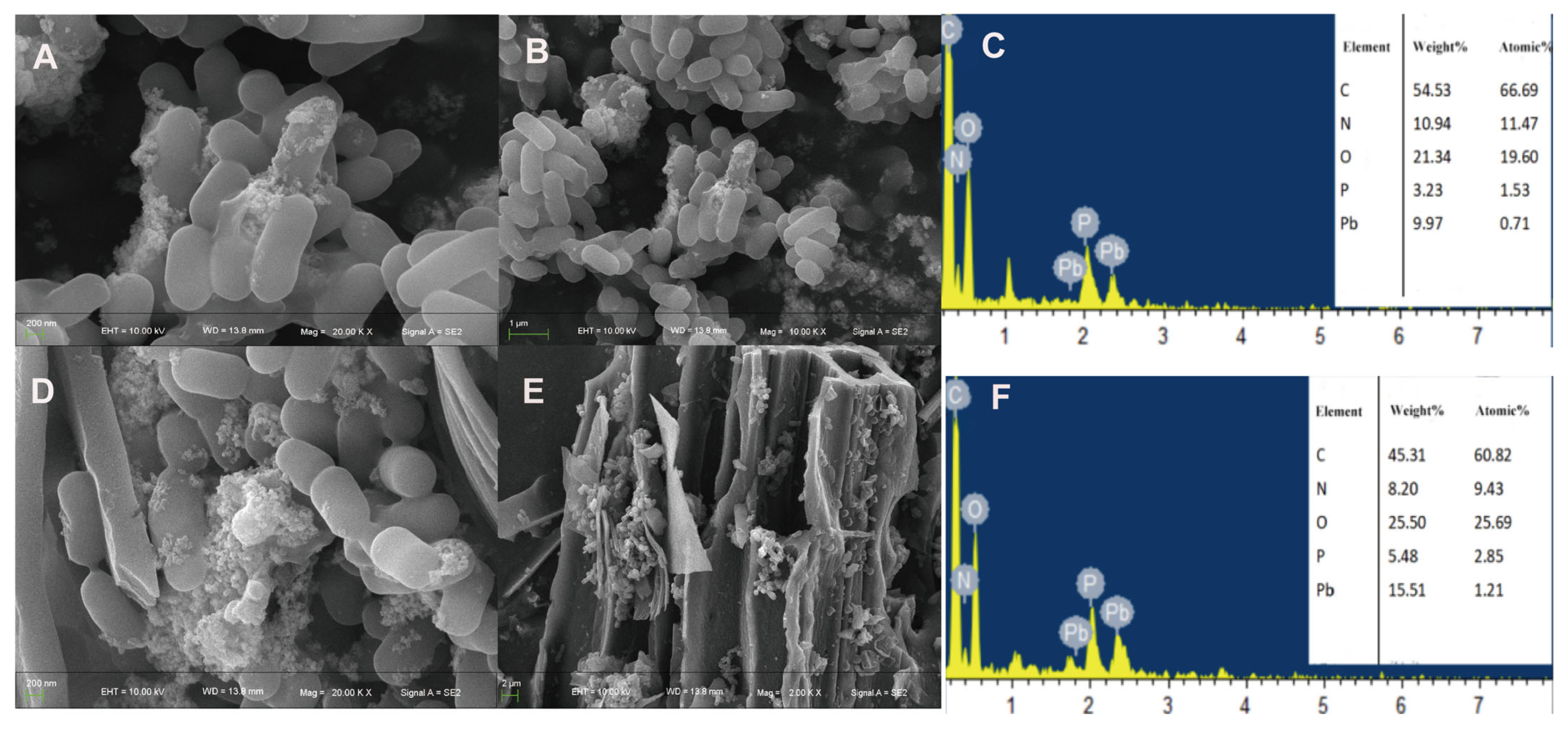
3.3.3. Characterization of Pb2+ and Al3+ Adsorption Capacity by SQ-2
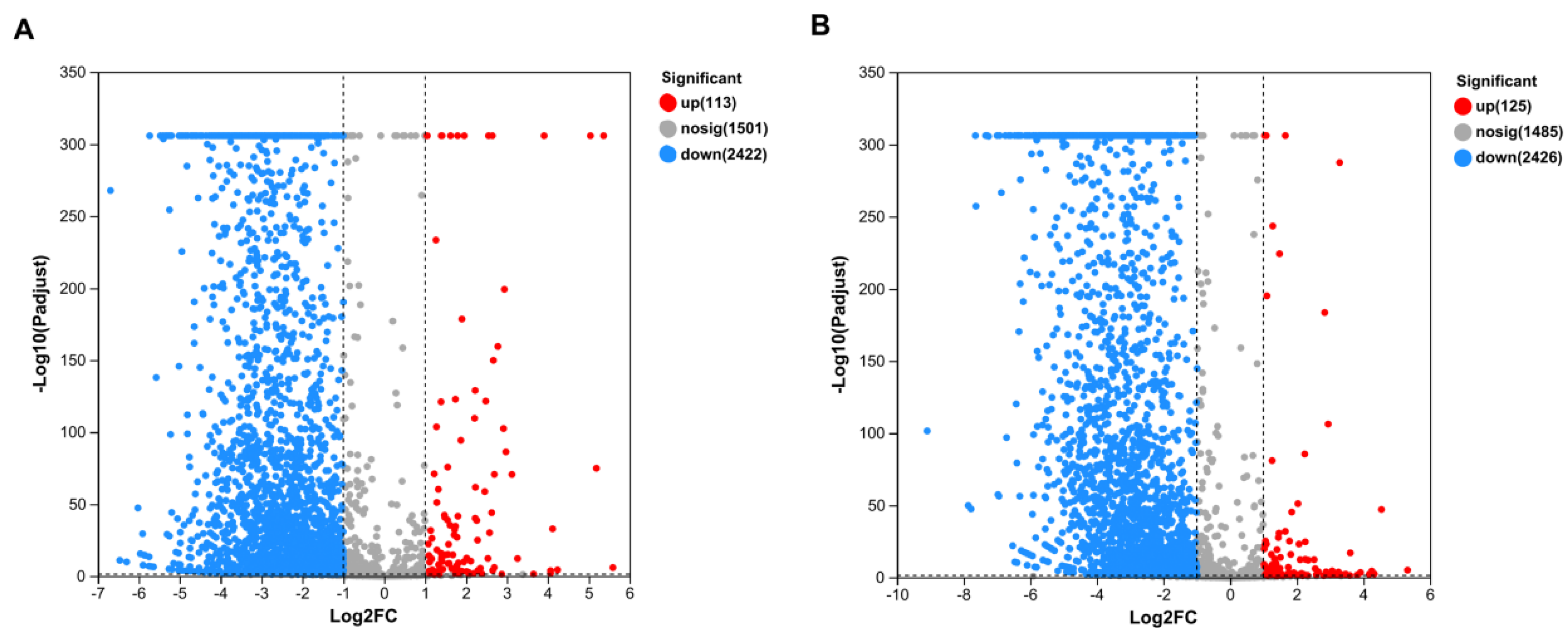
3.4. Effects of Pb2+ and Al3+ on SQ-2 Genome Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Xiang, M.; Li, Y.; Yang, J.; Lei, K.; Li, Y.; Li, F.; Zheng, D.; Fang, X.; Cao, Y. Metal contamination risk assessment and correlation analysis of metal contents in soil and crops. Environ. Pollut. 2021, 278, 116911. [Google Scholar] [CrossRef] [PubMed]
- Fasahat, P. Recent progress in understanding cadmium toxicity and tolerance in rice. Emir. J. Food Agric. 2015, 27, 94–105. [Google Scholar] [CrossRef]
- Meena, R.S.; Kumar, S.; Bohra, J.S.; Lal, R.; Yadav, G.S.; Pandey, A. Response of alley cropping-grown sesame to lime and sulphur on yield and available nutrient status in an acidic soil of Eastern India. Energy Ecol. Environ. 2019, 4, 65–74. [Google Scholar] [CrossRef]
- Shetty, R.; Prakash, N.B. Effect of different biochars on acid soil and growth parameters of rice plants under aluminium toxicity. Sci. Rep. 2020, 10, 12249. [Google Scholar] [CrossRef] [PubMed]
- Rigoletto, M.; Calza, P.; Gaggero, E.; Malandrino, M.; Fabbri, D. Bioremediation methods for the recovery of lead-contaminated soils: A review. Appl. Sci. 2020, 10, 3528. [Google Scholar] [CrossRef]
- Sun, Y.; Li, Y.; Xu, Y.; Liang, X.; Wang, L. In situ stabilization remediation of cadmium (Cd) and lead (Pb) co-contaminated paddy soil using bentonite. Appl. Clay Sci. 2015, 105, 200–206. [Google Scholar] [CrossRef]
- Lin, X.-R.; Chen, H.-B.; Li, Y.-X.; Zhou, Z.-H.; Li, J.-B.; Wang, Y.-Q.; Zhang, H.; Zhang, Y.; Han, Y.-H.; Wang, S.-S. Priestia sp. LWS1 Is a Selenium-Resistant Plant Growth-Promoting Bacterium That Can Enhance Plant Growth and Selenium Accumulation in Oryza sativa L. Agronomy 2022, 12, 1301. [Google Scholar] [CrossRef]
- Khumairah, F.H.; Setiawati, M.R.; Fitriatin, B.N.; Simarmata, T.; Alfaraj, S.; Ansari, M.J.; El Enshasy, H.A.; Sayyed, R.Z.; Najafi, S. Halotolerant plant growth promoting rhizobacteria isolated from saline soil improve nitrogen fixation and alleviate salt stress in rice plants. Front. Microbiol. 2022, 13, 905210. [Google Scholar] [CrossRef]
- Bach, E.; Lisboa, B.B.; Passaglia, L.M.P. Evaluation of biological control and rhizosphere competence of plant growth promoting bacteria. Appl. Soil Ecol. 2016, 99, 141–149. [Google Scholar] [CrossRef]
- Ahemad, M. Implications of bacterial resistance against metals in bioremediation: A review. J. Inst. Integr. Omics Appl. Biotechnol. (IIOAB) 2012, 3, 39–46. [Google Scholar]
- Akinrinlola, R.J.; Yuen, G.Y.; Drijber, R.A.; Adesemoye, A.O. Evaluation of Bacillus strains for plant growth promotion and predictability of efficacy by in vitro physiological traits. Int. J. Microbiol. 2018, 2018, 5686874. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; He, P.; Munir, S.; He, P.; Li, X.; Li, Y.; Wu, J.; Wu, Y.; Yang, L.; He, P.; et al. Efficacy of plant growth promoting bacteria Bacillus amyloliquefaciens B9601-Y2 for biocontrol of southern corn leaf blight. Biol. Control. 2019, 139, 104080. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Adesemoye, A.O. Improvement of crop protection and yield in hostile agroecological conditions with PGPR-based biofertilizer formulations. In Bioformulations: For Sustainable Agriculture; Springer: New Delhi, India, 2016; pp. 199–211. [Google Scholar]
- Govindasamy, V.; Senthilkumar, M.; Magheshwaran, V.; Kumar, U.; Bose, P.; Sharma, V.; Annapurna, K. Bacillus and Paenibacillus spp.: Potential PGPR for sustainable agriculture. In Plant Growth and Health Promoting Bacteria; Springer: Berlin/Heidelberg, Germany, 2011; pp. 333–364. [Google Scholar]
- Ongena, M.; Jacques, P. Bacillus lipopeptides: Versatile weapons for plant disease biocontrol. Trends Microbiol. 2008, 16, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Kavitha, B.; Reddy, P.V.L.; Kim, B.; Lee, S.S.; Pandey, S.K.; Kim, K.-H. Benefits and limitations of biochar amendment in agricultural soils: A review. J. Environ. Manag. 2018, 227, 146–154. [Google Scholar] [CrossRef]
- Qayyum, M.F.; Haider, G.; Raza, M.A.; Mohamed, A.K.S.; Rizwan, M.; El-Sheikh, M.A.; Alyemeni, M.N.; Ali, S. Straw-based biochar mediated potassium availability and increased growth and yield of cotton (Gossypium hirsutum L.). J. Saudi Chem. Soc. 2020, 24, 963–973. [Google Scholar] [CrossRef]
- Alvarez-Campos, O.; Lang, T.A.; Bhadha, J.H.; McCray, J.M.; Glaz, B.; Daroub, S.H. Biochar and mill ash improve yields of sugarcane on a sand soil in Florida. Agric. Ecosyst. Environ. 2018, 253, 122–130. [Google Scholar] [CrossRef]
- Mehmood, A.; Hussain, A.; Irshad, M.; Hamayun, M.; Iqbal, A.; Khan, N. In vitro production of IAA by endophytic fungus Aspergillus awamori and its growth promoting activities in Zea mays. Symbiosis 2019, 77, 225–235. [Google Scholar] [CrossRef]
- Batool, M.; Khan, W.-U.; Hamid, Y.; Farooq, M.A.; Naeem, M.A.; Nadeem, F. Interaction of pristine and mineral engineered biochar with microbial community in attenuating the heavy metals toxicity: A Review. Appl. Soil Ecol. 2022, 175, 104444. [Google Scholar] [CrossRef]
- Wu, C.; Zhi, D.; Yao, B.; Zhou, Y.; Yang, Y.; Zhou, Y. Immobilization of microbes on biochar for water and soil remediation: A Review. Environ. Res. 2022, 212, 113226. [Google Scholar] [CrossRef]
- Bandara, T.; Franks, A.; Xu, J.; Bolan, N.; Wang, H.; Tang, C. Chemical and biological immobilization mechanisms of potentially toxic elements in biochar-amended soils. Crit. Rev. Environ. Sci. Technol. 2020, 50, 903–978. [Google Scholar] [CrossRef]
- Tu, C.; Wei, J.; Guan, F.; Liu, Y.; Sun, Y.; Luo, Y. Biochar and bacteria inoculated biochar enhanced Cd and Cu immobilization and enzymatic activity in a polluted soil. Environ. Int. 2020, 137, 105576. [Google Scholar] [CrossRef]
- Zhao, S.; Zhao, S.; Wang, B. Combined application of biochar and PGPB on crop growth and heavy metals accumulation: A meta-Analysis. Environ. Pollut. 2025, 381, 126626. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Dai, S.; Huang, L.; Cui, Y.; Ying, M. Biocontrol Ability of Strain Bacillus amyloliquefaciens SQ-2 against Table Grape Rot Caused by Aspergillus tubingensis. J. Agric. Food Chem. 2024, 72, 24374–24386. [Google Scholar] [CrossRef]
- Zak, D.; Goldhammer, T.; Cabezas, A.; Gelbrecht, J.; Gurke, R.; Wagner, C.; Reuter, H.; Augustin, J.; Klimkowska, A.; McInnes, R. Top soil removal reduces water pollution from phosphorus and dissolved organic matter and lowers methane emissions from rewetted peatlands. J. Appl. Ecol. 2018, 55, 311–320. [Google Scholar] [CrossRef]
- Liu, T.; Xu, T.; Yu, F.; Yuan, Q.; Guo, Z.; Xu, B. A method combining ELM and PLSR (ELM-P) for estimating chlorophyll content in rice with feature bands extracted by an improved ant colony optimization algorithm. Comput. Electron. Agric. 2021, 186, 106177. [Google Scholar] [CrossRef]
- Quiles, F.A.; Galvez-Valdivieso, G.; Guerrero-Casado, J.; Pineda, M.; Piedras, P. Relationship between ureidic/amidic metabolism and antioxidant enzymatic activities in legume seedlings. Plant Physiol. Biochem. 2019, 138, 1–8. [Google Scholar] [CrossRef]
- Sanchez-Gonzalez, M.E.; Mora-Herrera, M.E.; Wong-Villarreal, A.; De La Portilla-López, N.; Sanchez-Paz, L.; Lugo, J.; Vaca-Paulín, R.; Del Aguila, P.; Yañez-Ocampo, G. Effect of pH and carbon source on phosphate solubilization by bacterial strains in pikovskaya medium. Microorganisms 2022, 11, 49. [Google Scholar] [CrossRef] [PubMed]
- Flores, J.P.M.; Alves, L.A.; de Oliveira Denardin, L.G.; Martins, A.P.; Bortoluzzi, E.C.; Inda, A.V.; de Faccio Carvalho, P.C.; Tiecher, T. Soil K forms and K budget in integrated crop-livestock systems in subtropical paddy fields. Soil Tillage Res. 2021, 213, 105070. [Google Scholar] [CrossRef]
- Nafees, M.; Ullah, S.; Ahmed, I. Modulation of drought adversities in Vicia faba by the application of plant growth promoting rhizobacteria and biochar. Microsc. Res. Tech. 2022, 85, 1856–1869. [Google Scholar] [CrossRef]
- Wang, D.; Gao, Y.; Sun, S.; Lu, X.; Li, Q.; Li, L.; Wang, K.; Liu, J. Effects of salt stress on the antioxidant activity and malondialdehyde, solution protein, proline, and chlorophyll contents of three Malus species. Life 2022, 12, 1929. [Google Scholar] [CrossRef]
- Malik, L.; Sanaullah, M.; Mahmood, F.; Hussain, S.; Shahzad, T. Co-application of biochar and salt tolerant PGPR to improve soil quality and wheat production in a naturally saline soil. Rhizosphere 2024, 29, 100849. [Google Scholar] [CrossRef]
- Deng, S.; Dong, H.; Lv, G.; Jiang, H.; Yu, B.; Bishop, M.E. Microbial dolomite precipitation using sulfate reducing and halophilic bacteria: Results from Qinghai Lake, Tibetan Plateau, NW China. Chem. Geol. 2010, 278, 151–159. [Google Scholar] [CrossRef]
- D’Souza, L.; Devi, P.; Divya Shridhar, M.P.; Naik, C.G. Use of Fourier Transform Infrared(FTIR)Spectroscopy to Study Cadmium-Induced Changes in Padina tetrastromatica (Hauck). Anal. Chem. Insights 2008, 3, 135. [Google Scholar] [CrossRef]
- Naik, M.M.; Pandey, A.; Dubey, S.K. Biological characterization of lead-enhanced exopolysaccharide produced by a lead resistant Enterobacter cloacae strain P2B. Biodegradation 2012, 23, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Hafez, E.M.; Alsohim, A.S.; Farig, M.; Omara, A.E.-D.; Rashwan, E.; Kamara, M.M. Synergistic effect of biochar and plant growth promoting rhizobacteria on alleviation of water deficit in rice plants under salt-affected soil. Agronomy 2019, 9, 847. [Google Scholar] [CrossRef]
- Ye, T.; Li, Y.; Zhang, J.; Hou, W.; Zhou, W.; Lu, J.; Xing, Y.; Li, X. Nitrogen, phosphorus, and potassium fertilization affects the flowering time of rice (Oryza sativa L.). Glob. Ecol. Conserv. 2019, 20, e00753. [Google Scholar] [CrossRef]
- Schmierer, M.; Knopf, O.; Asch, F. Growth and photosynthesis responses of a super dwarf rice genotype to shade and nitrogen supply. Rice Sci. 2021, 28, 178–190. [Google Scholar] [CrossRef]
- Zhao, L.-S.; Li, K.; Wang, Q.-M.; Song, X.-Y.; Su, H.-N.; Xie, B.-B.; Zhang, X.-Y.; Huang, F.; Chen, X.-L.; Zhou, B.-C.; et al. Nitrogen starvation impacts the photosynthetic performance of Porphyridium cruentum as revealed by chlorophyll a fluorescence. Sci. Rep. 2017, 7, 8542. [Google Scholar] [CrossRef]
- Bashir, Z.; Zargar, M.Y.; Vishwakarma, D.K. Potassium-solubilizing microorganisms for sustainable agriculture. Appl. Agric. Pract. Mitigating Clim. Change 2019, 2, 17–28. [Google Scholar]
- Meena, V.S.; Maurya, B.R.; Verma, J.P.; Aeron, A.; Kumar, A.; Kim, K.; Bajpai, V.K. Potassium solubilizing rhizobacteria (KSR): Isolation, identification, and K-release dynamics from waste mica. Ecol. Eng. 2015, 81, 340–347. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.F.; Wu, W.H. Potassium and phosphorus transport and signaling in plants. J. Integr. Plant Biol. 2021, 63, 34–52. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.X.; Weng, X.Y.; Yang, Y. Effect of phosphorus deficiency on the photosynthetic characteristics of rice plants. Russ. J. Plant Physiol. 2007, 54, 741–748. [Google Scholar] [CrossRef]
- Maqsood, M.; Shehzad, M.A.; Wahid, A.; Butt, A.A. Improving Drought Tolerance in Maize (Zea mays) with Potassium Application in Furrow Irrigation Systems. Int. J. Agric. Biol. 2013, 15, 1193–1198. [Google Scholar]
- Gupta, G.; Parihar, S.S.; Ahirwar, N.K.; Snehi, S.K.; Singh, V. Plant growth promoting rhizobacteria (PGPR): Current and future prospects for development of sustainable agriculture. J. Microb. Biochem. Technol. 2015, 7, 96–102. [Google Scholar]
- Fageria, N.K. Potassium requirements of lowland rice. Commun. Soil Sci. Plant Anal. 2015, 46, 1459–1472. [Google Scholar] [CrossRef]
- Geng, A.; Wang, X.; Wu, L.; Wang, F.; Wu, Z.; Yang, H.; Chen, Y.; Wen, D.; Liu, X. Silicon improves growth and alleviates oxidative stress in rice seedlings (Oryza sativa L.) by strengthening antioxidant defense and enhancing protein metabolism under arsanilic acid exposure. Ecotoxicol. Environ. Saf. 2018, 158, 266–273. [Google Scholar] [CrossRef]
- Feng, L.; Li, Q.; Zhou, D.; Jia, M.; Liu, Z.; Hou, Z.; Ren, Q.; Ji, S.; Sang, S.; Lu, S.; et al. B. subtilis CNBG-PGPR-1 induces methionine to regulate ethylene pathway and ROS scavenging for improving salt tolerance of tomato. Plant J. 2024, 117, 193–211. [Google Scholar] [CrossRef]
- Ren, X.; Zhu, J.; Liu, H.; Xu, X.; Liang, C. Response of antioxidative system in rice (Oryza sativa) leaves to simulated acid rain stress. Ecotoxicol. Environ. Saf. 2018, 148, 851–856. [Google Scholar] [CrossRef]
- Bhadwal, S.; Sharma, S.; Singh, D. Interactive effects of selenium and arsenic on phenolic constituents and antioxidant activity in rice (Oryza sativa L.). Chemosphere 2024, 350, 141071. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Alyemeni, M.N.; Rinklebe, J.; Ahmad, P. Citric acid and hydrogen sulfide cooperate to mitigate chromium stress in tomato plants by modulating the ascorbate-glutathione cycle, chromium sequestration, and subcellular allocation of chromium. Environ. Pollut. 2023, 335, 122292. [Google Scholar] [CrossRef]
- Zhi, Y.; Li, X.; Wang, X.; Jia, M.; Wang, Z. Photosynthesis promotion mechanisms of artificial humic acid depend on plant types: A hydroponic study on C3 and C4 plants. Sci. Total Environ. 2024, 917, 170404. [Google Scholar] [CrossRef] [PubMed]
- Rudinskienė, A.; Marcinkevičienė, A.; Velička, R.; Kosteckas, R.; Kriaučiūnienė, Z.; Vaisvalavičius, R. The comparison of soil agrochemical and biological properties in the multi-cropping farming systems. Plants 2022, 11, 774. [Google Scholar] [CrossRef] [PubMed]
- Oleszczuk, P.; Jośko, I.; Futa, B.; Pasieczna-Patkowska, S.; Pałys, E.; Kraska, P. Effect of pesticides on microorganisms, enzymatic activity and plant in biochar-amended soil. Geoderma 2014, 214, 10–18. [Google Scholar] [CrossRef]
- Cao, D.; Chen, X.; Nan, J.; Wang, A.; Li, Z. Biomolecular insights into the inhibition of heavy metals on reductive dechlorination of 2,4,6-trichlorophenol in Pseudomonas sp. CP-1. Water Res. 2023, 247, 120836. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Root Dry Weight (g) | Stem Dry Weight (g) | Root Fresh Weight (g) | Stem Fresh Weight (g) | Stem Thickness (mm) | Plant Height (cm) |
|---|---|---|---|---|---|---|
| CK | 0.101 ± 0.003 a | 0.051 ± 0.002 a | 1.022 ± 0.014 a | 0.48 ± 0.02 a | 0.78 ± 0.01 a | 12.3 ± 0.2 a |
| M | 0.072 ± 0.002 c | 0.032 ± 0.001 b | 0.651 ± 0.015 c | 0.31 ± 0.01 b | 0.72 ± 0.02 b | 9.7 ± 0.2 c |
| S + M | 0.082 ± 0.002 bc | 0.041 ± 0.003 a | 0.773 ± 0.018 bc | 0.43 ± 0.03 a | 0.74 ± 0.01 ab | 11.5 ± 0.2 b |
| S + M + C | 0.091 ± 0.001 bc | 0.042 ± 0.002 a | 0.831 ± 0.012 b | 0.44 ± 0.01 a | 0.74 ± 0.02 ab | 11.7 ± 0.2 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, G.; Li, X.; Ma, J.; Cui, Y.; Ying, M.; Huang, L.; Li, M. Bacillus amyloliquefaciens SQ-2 and Biochar: A Promising Combination for Enhancing Rice Growth in Pb/Al-Contaminated Acidic Soils. Microorganisms 2025, 13, 1556. https://doi.org/10.3390/microorganisms13071556
Gao G, Li X, Ma J, Cui Y, Ying M, Huang L, Li M. Bacillus amyloliquefaciens SQ-2 and Biochar: A Promising Combination for Enhancing Rice Growth in Pb/Al-Contaminated Acidic Soils. Microorganisms. 2025; 13(7):1556. https://doi.org/10.3390/microorganisms13071556
Chicago/Turabian StyleGao, Guohui, Xue Li, Jiajun Ma, Yumeng Cui, Ming Ying, Lei Huang, and Meitong Li. 2025. "Bacillus amyloliquefaciens SQ-2 and Biochar: A Promising Combination for Enhancing Rice Growth in Pb/Al-Contaminated Acidic Soils" Microorganisms 13, no. 7: 1556. https://doi.org/10.3390/microorganisms13071556
APA StyleGao, G., Li, X., Ma, J., Cui, Y., Ying, M., Huang, L., & Li, M. (2025). Bacillus amyloliquefaciens SQ-2 and Biochar: A Promising Combination for Enhancing Rice Growth in Pb/Al-Contaminated Acidic Soils. Microorganisms, 13(7), 1556. https://doi.org/10.3390/microorganisms13071556