
Biosurfactant Produced by Bacillus subtilis UCP 1533 Isolated from the Brazilian Semiarid Region: Characterization and Antimicrobial Potential

 ,
, 
 , ,
, ,  and
and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Soil Collection and Microorganism Identification
2.1.1. Microorganism Identification by MALDI-TOF MS
2.1.2. Gram Staining
2.1.3. Spore Staining by Wirtz-Conklin Technique
2.1.4. Catalase Test
2.2. Preparation and Growth of the Inoculum
2.3. Selection of Cultivation Conditions for Biosurfactant Production
2.4. Biosurfactant Production
2.5. Determination of Surface Tension and Critical Micelle Concentration (CMC)
2.6. Determination of Emulsifying Activity
2.7. Evaluation of Biosurfactant Stability (Effects of pH, Temperature, and NaCl)
2.8. Biosurfactant Isolation
2.9. Physicochemical Characterization of the Biosurfactant
2.10. Ionic Charge Determination
2.11. Toxicological Analysis
2.11.1. Toxicity Assessment of the Biosurfactant in Tenebrio molitor
2.11.2. Phytotoxicity in Solanum lycopersicon and Lactuca sativa
2.12. Disk Diffusion Assay of the Biosurfactant
2.13. Statistical Analysis
3. Results and Discussion
3.1. Microorganism Identification
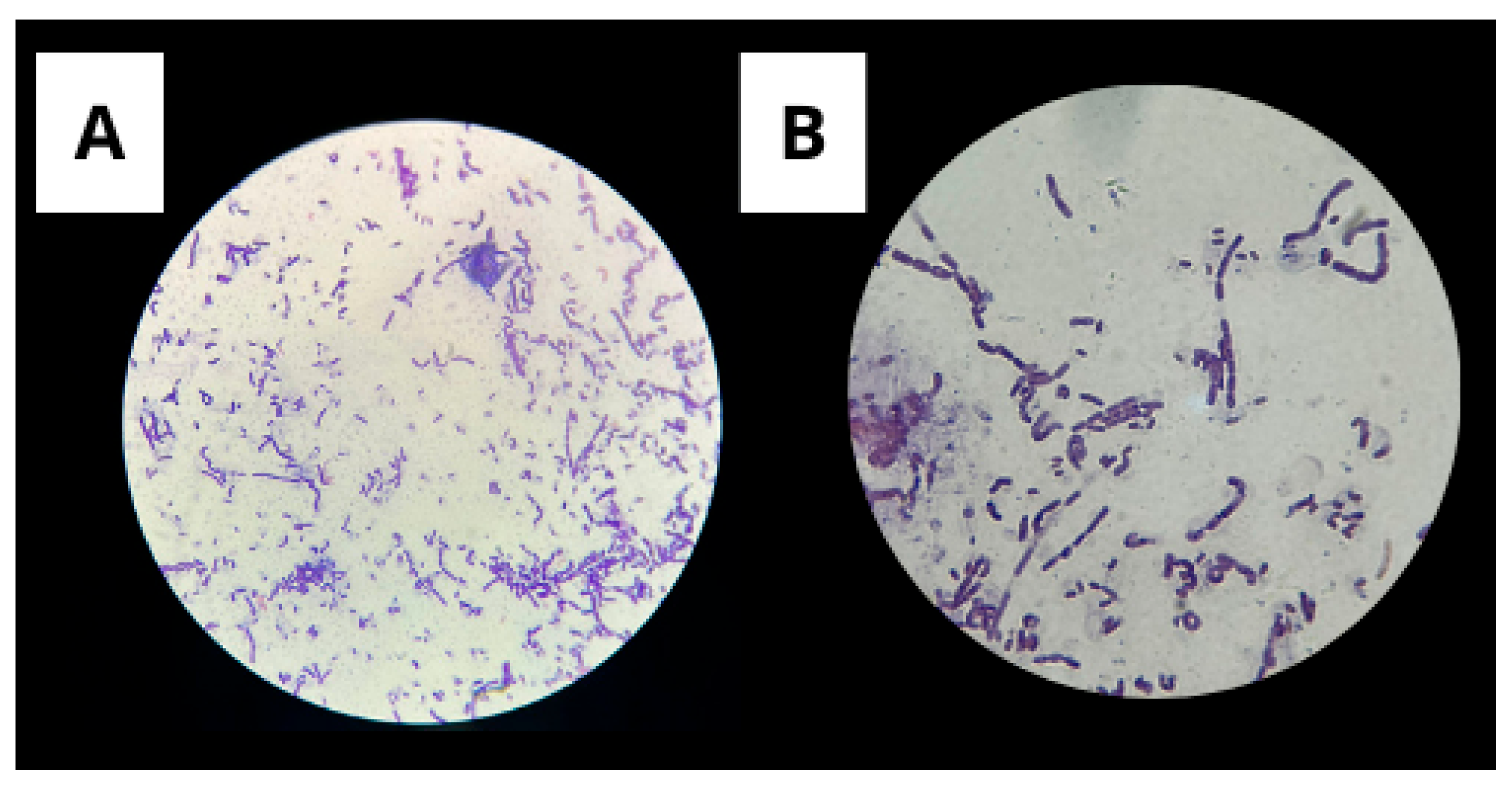
3.1.1. Gram Staining
3.1.2. Spore Staining by the Wirtz-Conklin Technique
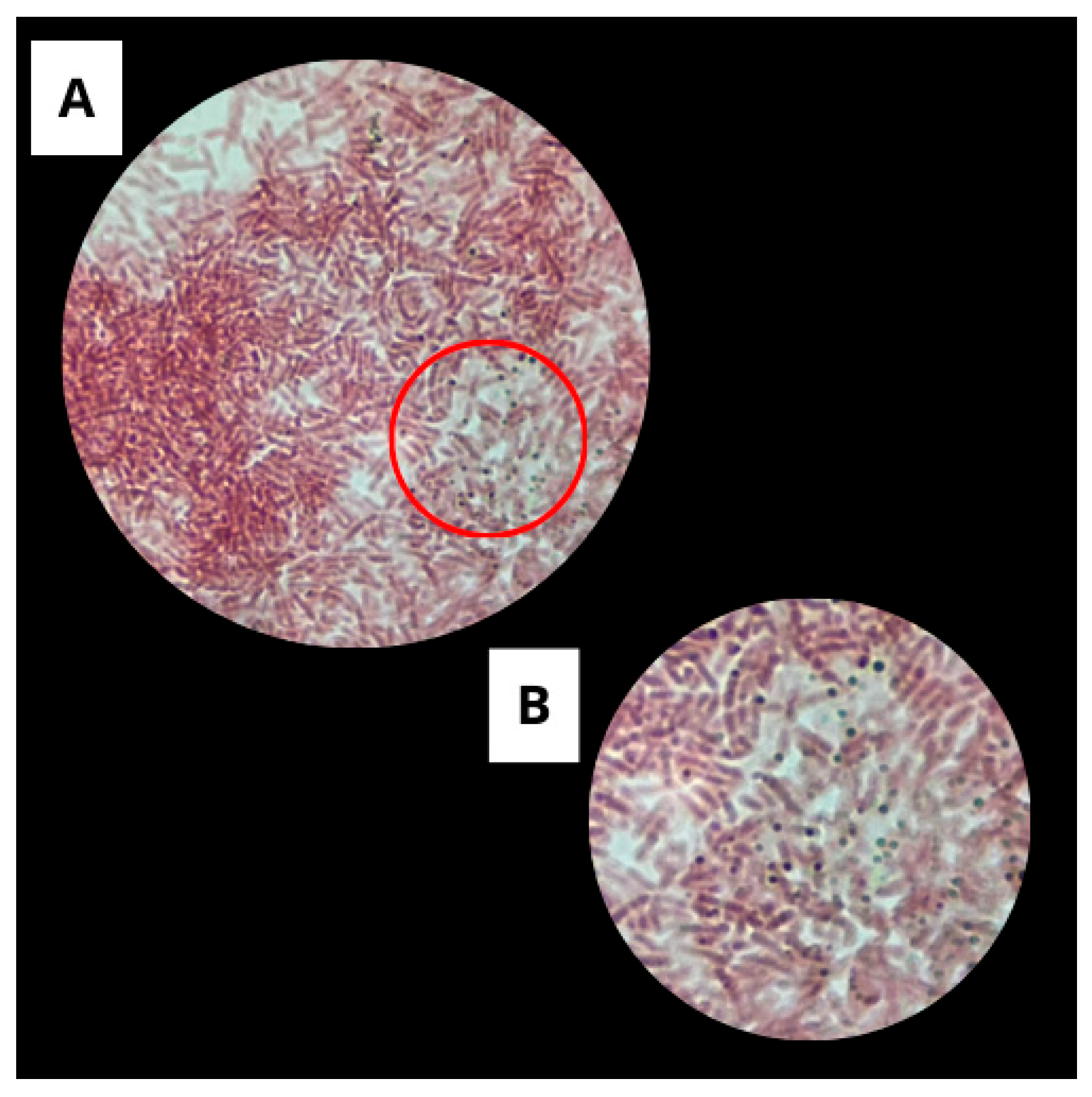
3.1.3. Catalase Test
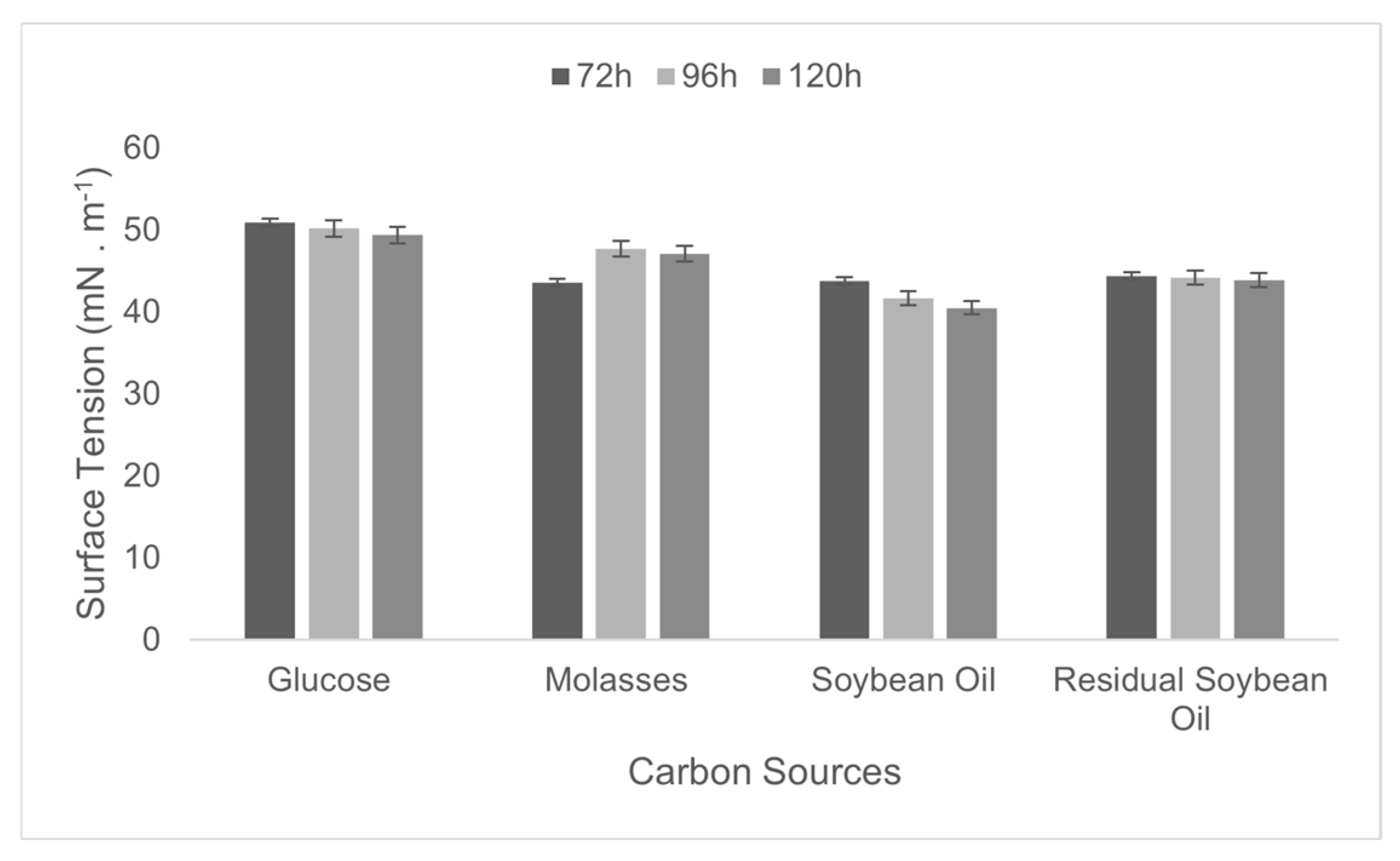
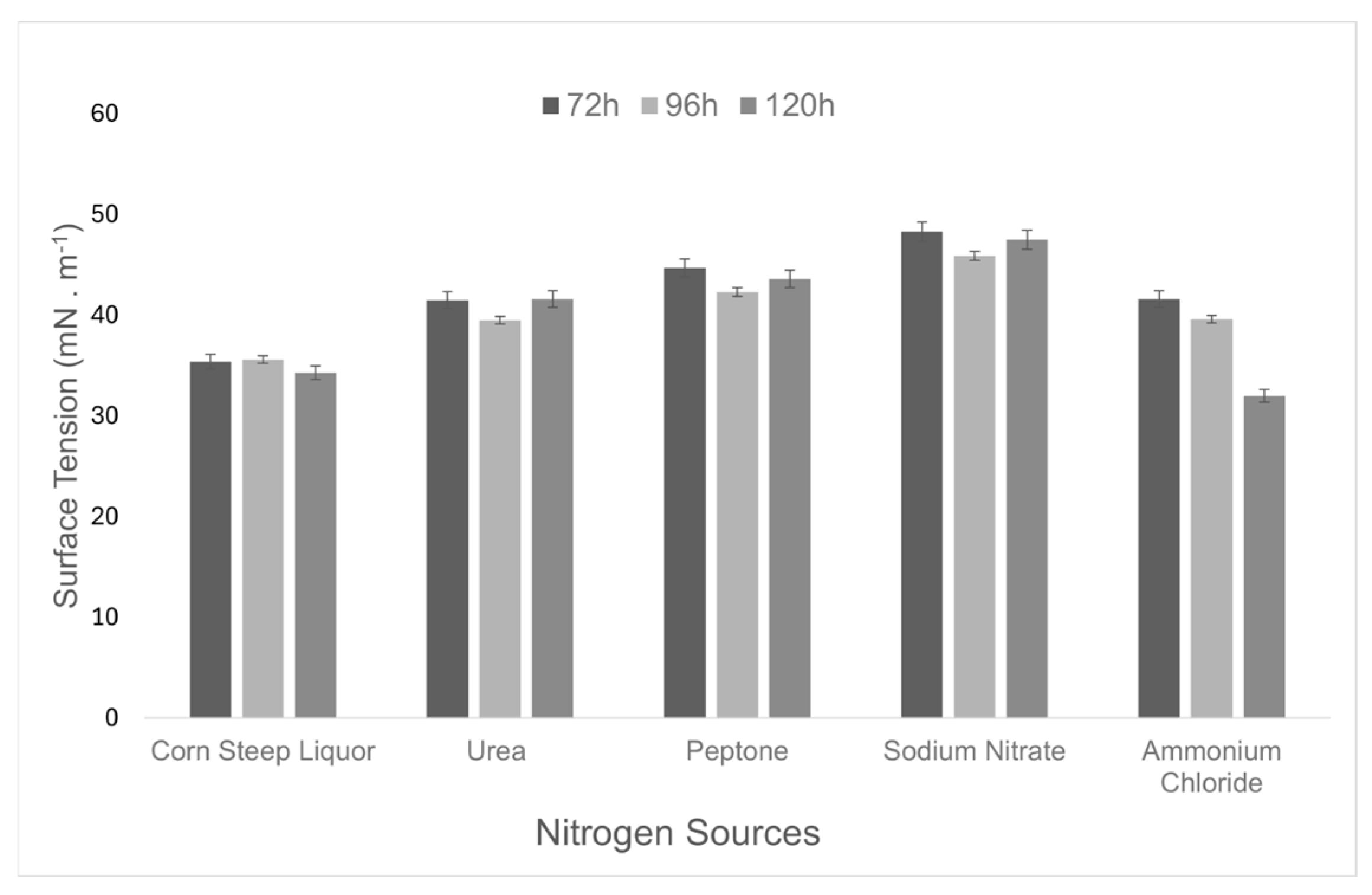
3.2. Selection of Cultivation Conditions for Biosurfactant Production
3.3. Biosurfactant Production and Yield
3.4. Physicochemical Properties of the Biosurfactant
3.5. Biosurfactant Stability
3.6. Determination of Ionic Charge
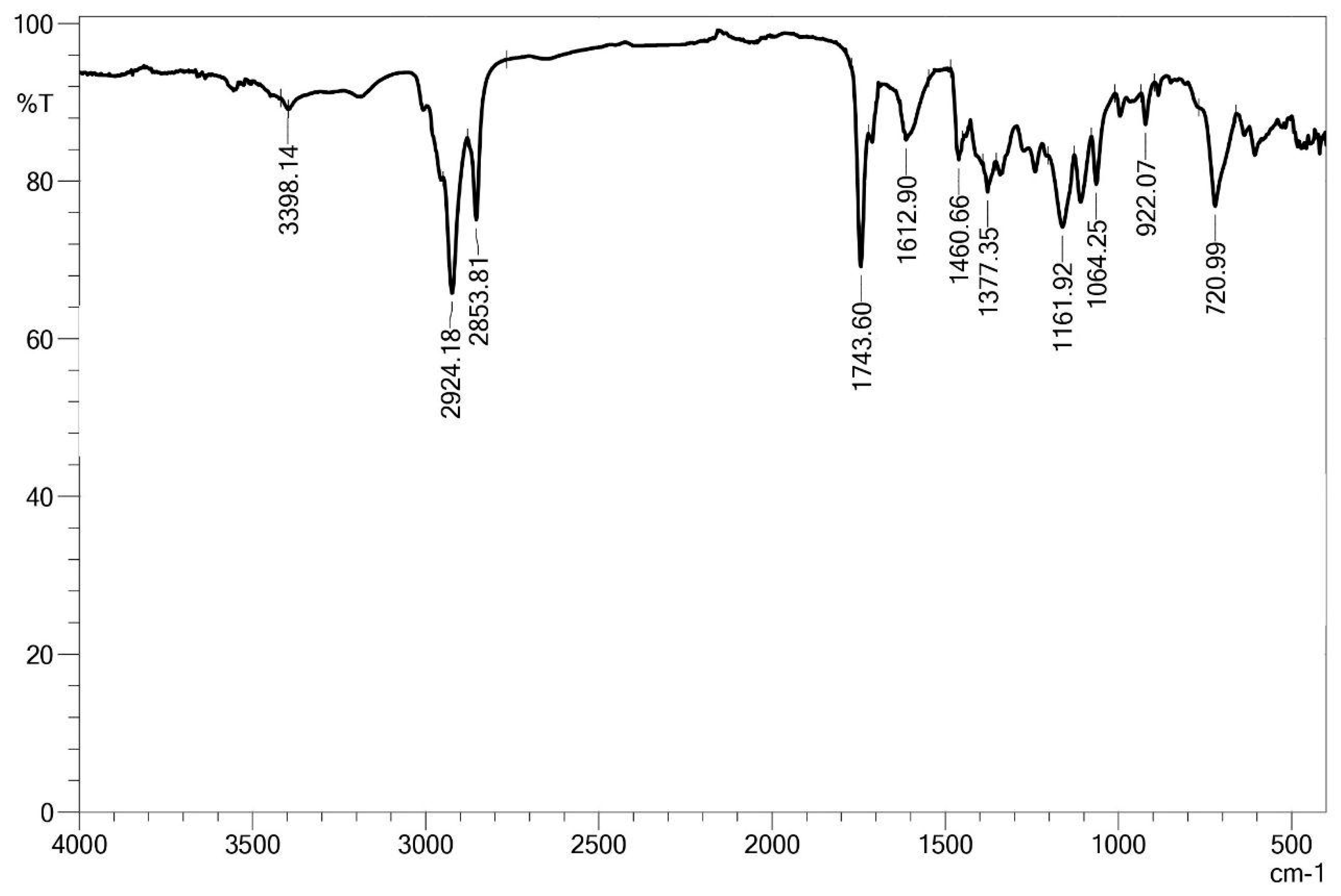
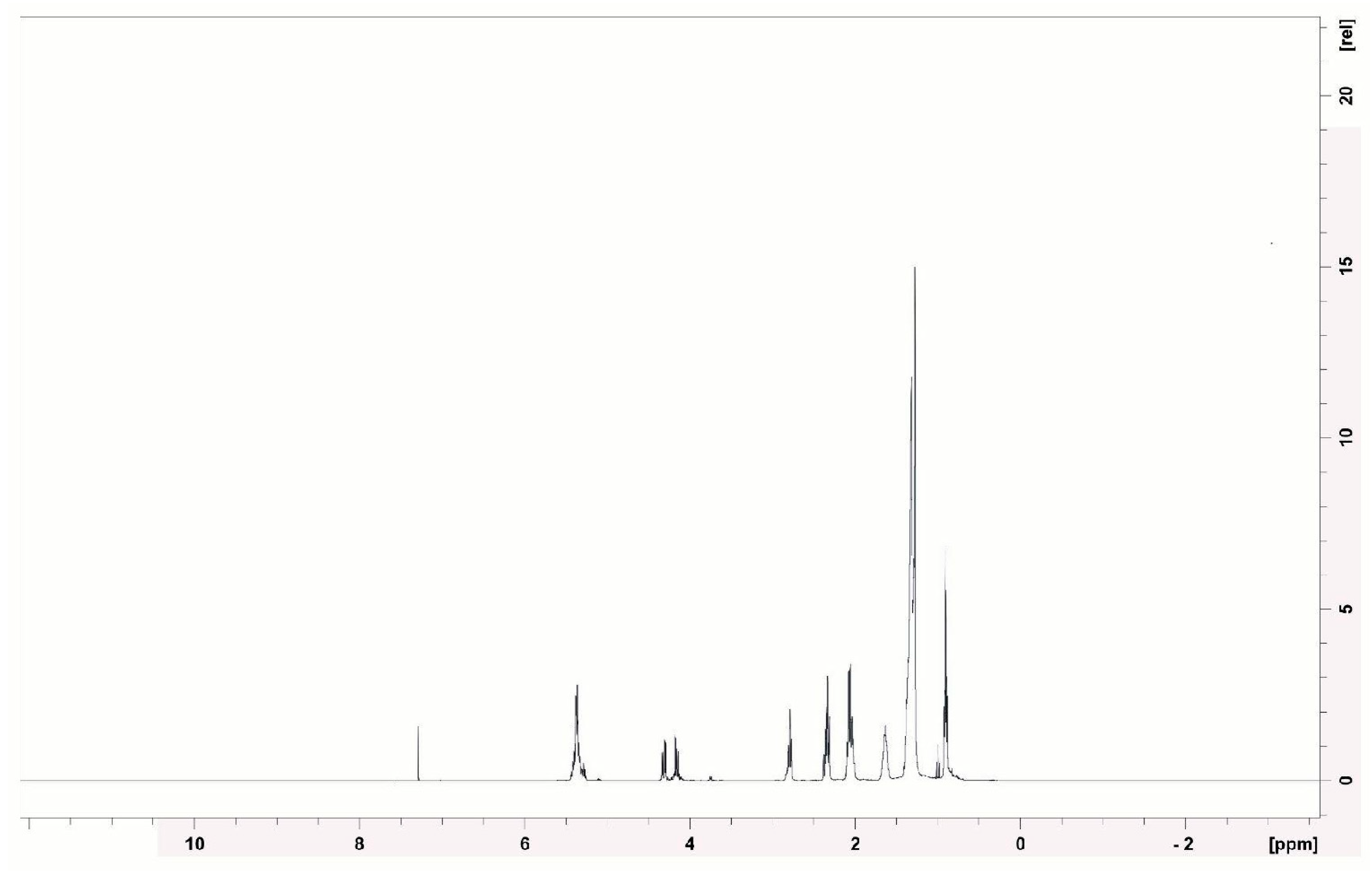
3.7. Structural Characterization of the Biosurfactant Produced by Bacillus subtilis UCP 1533
3.8. Evaluation of the Biosurfactant Toxicity
3.8.1. Phytotoxicity
3.8.2. Toxicity to Tenebrio molitor
3.9. Antimicrobial Activity of Biosurfactant
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Benali, T.; Lemhadri, A.; Harboul, K.; Chtibi, H.; Khabbach, A.; Jadouali, S.M.; Quesada-Romero, L.; Louahlia, S.; Hammani, K.; Ghaleb, A.; et al. Chemical profiling and biological properties of essential oils of Lavandula stoechas L. collected from three Moroccan sites: In vitro and in silico investigations. Plants 2023, 12, 1413. [Google Scholar] [CrossRef] [PubMed]
- Elghoul, M.; Bouassida, M.; Ghribi, D.; Mnif, I. A new bacterial-derived biosurfactant for biotechnological applications in the oil industry: Production, optimization, biosurfactant functional and physicochemical characterization. Water Pract. Technol. 2023, 18, 810–830. [Google Scholar] [CrossRef]
- Caioni, G.; Reyes, C.P.; Laurenti, D.; Chiaradia, C.; Dainese, E.; Mattioli, R.; Di Risola, D.; Santavicca, E.; Francioso, A. Biochemistry and future perspectives of antibiotic resistance: An eye on active natural products. Antibiotics 2024, 13, 1071. [Google Scholar] [CrossRef]
- Liu, H.Y.; Prentice, E.L.; Webber, M.A. Mechanisms of antimicrobial resistance in biofilms. npj Antimicrob. Resist. 2024, 2, 27. [Google Scholar] [CrossRef]
- GBD 2021 Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance 1990–2021: A systematic analysis with forecasts to 2050. Lancet 2024, 404, 1199–1226. [Google Scholar] [CrossRef]
- Rahman, S.; Hollis, A. The effect of antibiotic usage on resistance in humans and food-producing animals: A longitudinal, One Health analysis using European data. Front. Public Health 2023, 11, 1170426. [Google Scholar] [CrossRef]
- Igarashi, M. New natural products to meet the antibiotic crisis: A personal journey. J. Antibiot. 2019, 72, 890–898. [Google Scholar] [CrossRef]
- Lourenço, M.; Duarte, N.; Ribeiro, I.A.C. Exploring biosurfactants as antimicrobial approaches. Pharmaceuticals 2024, 17, 1239. [Google Scholar] [CrossRef]
- Shaji, A.; Thamarai, P.; Deivayanai, V.C.; Saravanan, A.; Yaashikaa, P.R. Progress in sustainable remediation: Utilizing biosurfactants for eco-friendly contaminant cleanup. Bioresour. Technol. Rep. 2024, 27, 101901. [Google Scholar] [CrossRef]
- Banat, I.M.; Carboue, Q.; Saucedo-Castaneda, G.; de Jesus Cazares-Marinero, J. Biosurfactants: The green generation of speciality chemicals and potential production using solid-state fermentation (SSF) technology. Bioresour. Technol. 2021, 320, 124222. [Google Scholar] [CrossRef] [PubMed]
- Datta, D.; Ghosh, S.; Kumar, S.; Gangola, S.; Majumdar, B.; Saha, R.; Mazumdar, S.P.; Singh, S.V.; Kar, G. Microbial biosurfactants: Multifarious applications in sustainable agriculture. Microbiol. Res. 2024, 279, 127551. [Google Scholar] [CrossRef] [PubMed]
- Marchut-Mikołajczyk, O.; Drożdżyński, P.; Polewczyk, A.; Smułek, W.; Antczak, T. Biosurfactant from endophytic Bacillus pumilus 2A: Physicochemical characterization, production and optimization and potential for plant growth promotion. Microb. Cell Fact. 2021, 20, 40. [Google Scholar] [CrossRef] [PubMed]
- Ambust, S.; Das, A.J.; Kumar, R. Bioremediation of petroleum contaminated soil through biosurfactant and Pseudomonas sp. SA3 amended design treatments. Curr. Res. Microb. Sci. 2021, 2, 100031. [Google Scholar] [CrossRef]
- Gudiña, E.J.; Fernandes, E.C.; Rodrigues, A.I.; Teixeira, J.A.; Rodrigues, L.R. Biosurfactants produced by Bacillus subtilis using corn steep liquor as culture medium. Front. Microbiol. 2015, 6, 59. [Google Scholar] [CrossRef]
- Paiva, M.M.; Santos, D.T. Antimicrobial and emulsifying properties of biosurfactants produced by marine bacteria: A review. Braz. J. Dev. 2023, 9, 13241–13256. [Google Scholar]
- Leal, E.; Teixeira, J.A.; Gudiña, E.J. Development of foam-free biosurfactant production processes using Bacillus licheniformis. Fermentation 2024, 10, 340. [Google Scholar] [CrossRef]
- Parvin, A.; Adhikary, R.; Guha, S.; Mitra, P.K.; Mandal, V. Antibiofilm and antimicrobial activity of biosurfactants from two Lactiplantibacillus pentosus strains against food and topical pathogens. J. Food Process. Preserv. 2022, 46, e16927. [Google Scholar] [CrossRef]
- Lima-Neto, R.; Santos, C.; Lima, N.; Sampaio, P.; Pais, C.; Neves, R.P. Application of MALDI-TOF MS for requalification of a Candida clinical isolates culture collection. Braz. J. Microbiol. 2014, 45, 515–522. [Google Scholar] [CrossRef]
- Cappuccino, J.G.; Sherman, N. Microbiologia: Uma Abordagem de Laboratório, 11th ed.; Pearson: London, UK, 2017. [Google Scholar]
- Mac Faddin, J.F. Biochemical Tests for Identification of Medical Bacteria, 3rd ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2000. [Google Scholar]
- Cooper, D.G.; Goldenberg, B.G. Surface active agents from two Bacillus species. Appl. Environ. Microbiol. 1987, 53, 224–229. [Google Scholar] [CrossRef]
- Meylheuc, T.; Van Oss, C.J.; Bellon-Fontaine, M.N. Adsorption of biosurfactant on solid surfaces and consequences regarding the bioadhesion of Listeria monocytogenes LO28. J. Appl. Microbiol. 2001, 91, 822–832. [Google Scholar] [CrossRef] [PubMed]
- Silva, T.F.S.; Cavalcanti Filho, J.R.N.; Fonsêca, M.M.L.B.; Santos, N.M.; Silva, A.C.B.; Zagmignan, A.; Abreu, A.G.; Silva, A.P.S.; Lima, V.L.M.; Silva, N.H.; et al. Products derived from Buchenavia tetraphylla leaves have in vitro antioxidant activity and protect Tenebrio molitor larvae against Escherichia coli-induced injury. Pharmaceuticals 2020, 13, 46. [Google Scholar] [CrossRef] [PubMed]
- Tiquia, S.M.; Tam, N.F.Y.; Hodgkiss, I.J. Effects of composting on phytotoxicity of spent pig-manure sawdust litter. Environ. Pollut. 1996, 93, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Leyton, A.; Araya, M.; Tala, F.; Flores, L.; Lienqueo, M.E.; Shene, C. Macrocystis pyrifera extract residual as nutrient source for the production of sophorolipids compounds by marine yeast Rhodotorula rubra. Molecules 2021, 26, 2355. [Google Scholar] [CrossRef]
- Dib, C.C. Utilização do MALDI-TOF MS para identificação de micobactérias em amostras clínicas de animais. Pubvet 2023, 17, e1414. [Google Scholar] [CrossRef]
- Bier, D.; Tutija, J.F.; Pasquatti, T.N.; Oliveira, T.L.; Araújo, F.R.; Verbisck, N.V. Identificação por espectrometria de massa MALDI-TOF de Salmonella spp. e Escherichia coli isolados de carcaças bovinas. Pesq. Vet. Bras. 2017, 37, 1373–1379. [Google Scholar] [CrossRef]
- Verbisck, N.; Suniga, P.; Mantovani, C.; Souza, I.; Dos Santos, M.G.; Araujo, F.; Dos Santos, L.R. Espectro de Massas de Referência para Identificação de Burkholderia mallei por Espectrometria de Massas MALDI-TOF, 7th ed.; Embrapa: Brasília, Brazil, 2022. [Google Scholar]
- Costa, R.G.; Esteves, W.T.C.; Da Rocha Nogueira, J.M. Principais métodos clássicos de coloração em bacteriologia: Aplicações, técnicas, fundamentos e limitações. RBAC 2024, 56, 183–196. [Google Scholar] [CrossRef]
- Cho, W.-I.; Chung, M.-S. Bacillus spores: A review of their properties and inactivation processing technologies. Food Sci. Biotechnol. 2020, 29, 1447–1461. [Google Scholar] [CrossRef]
- Zhang, X.; Al-Dossary, A.; Hussain, M.; Setlow, P.; Li, J. Applications of Bacillus subtilis spores in biotechnology and advanced materials. Appl. Environ. Microbiol. 2020, 86, e01096-20. [Google Scholar] [CrossRef]
- Biermann, R.; Beutel, S. Endospore production of Bacillus spp. for industrial use. Eng. Life Sci. 2023, 23, e2300013. [Google Scholar] [CrossRef]
- Riley, E.P.; Schwarz, C.; Derman, A.I.; Lopez-Garrido, J. Milestones in Bacillus subtilis sporulation research. Microb. Cell 2020, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liu, Z.; Zhao, J.; Zhao, Z.M. Mechanisms and strategies for regulating sporulation and germination of Bacillus subtilis to enhance probiotic effects. Ind. Crops Prod. 2024, 220, 119178. [Google Scholar] [CrossRef]
- Jiang, M.; Liu, Y.; Xue, H.; Wang, Y.; Wang, C.; Yang, F.; Li, X. Expression and biochemical characterization of a Bacillus subtilis catalase in Pichia pastoris X-33. Protein Expr. Purif. 2023, 208–209, 106277. [Google Scholar] [CrossRef] [PubMed]
- Bano, S.; Iqbal, S.; Abbasi, K.; Siddiqui, K.; Aman, A. Effect of oxidative stress on catalase induction and optimization of kinetic performance of enzyme from B. subtilis KIBGE HAS 1. Bact. Emp. 2022, 5, e433. [Google Scholar] [CrossRef]
- Qamar, S.A.; Pacifico, S. Cleaner production of biosurfactants via bio-waste valorization: A comprehensive review of characteristics, challenges, and opportunities in bio-sector applications. J. Environ. Chem. Eng. 2023, 11, 111555. [Google Scholar] [CrossRef]
- Begum, W.; Saha, B.; Mandal, U. A comprehensive review on production of bio-surfactants by bio-degradation of waste carbohydrate feedstocks: An approach towards sustainable development. RSC Adv. 2023, 13, 25599–25615. [Google Scholar] [CrossRef]
- Pinto, M.I.S.; Campos Guerra, J.M.; Meira, H.M.; Sarubbo, L.A.; de Luna, J.M. A biosurfactant from Candida bombicola: Its synthesis, characterization, and its application as a food emulsions. Foods 2022, 11, 561. [Google Scholar] [CrossRef]
- Machado, C.M.B.; Vandenberghe, L.P.S.; Mello, A.F.M.; Soccol, C.R. Corn or soybean oil as the sole carbon source for polyhydroxybutyrate production in a biofuel biorefinery concept. Polymers 2025, 17, 324. [Google Scholar] [CrossRef]
- Garcia-Sanchez, F.; Camara-Zapata, J.M.; Navarro-Morillo, I. Use of corn steep liquor as a biostimulant in agriculture. Horticulturae 2024, 10, 315. [Google Scholar] [CrossRef]
- Selva Filho, A.A.P.; Faccioli, Y.E.; Converti, A.; da Silva, R.D.C.F.S.; Sarubbo, L.A. Maximization of the production of a low-cost biosurfactant for application in the treatment of soils contaminated with hydrocarbons. Sustainability 2024, 16, 7970. [Google Scholar] [CrossRef]
- Ciurko, D.; Czyżnikowska, Ż.; Kancelista, A.; Łaba, W.; Janek, T. Sustainable production of biosurfactant from agro-industrial oil wastes by Bacillus subtilis and its potential application as antioxidant and ACE inhibitor. Int. J. Mol. Sci. 2022, 23, 10824. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.R.; Caldas, M.C.; Lima, C.V.; Meira, H.M.; Sarubbo, L.A.; Luna, J.M. Production, characterization and application of biosurfactant for cleaning cotton fabric and removing oil from contaminated sand. Processes 2024, 12, 2584. [Google Scholar] [CrossRef]
- Wu, B.; Xiu, J.; Yu, L.; Huang, L.; Yi, L.; Ma, Y. Biosurfactant production by Bacillus subtilis SL and its potential for enhanced oil recovery in low permeability reservoirs. Sci. Rep. 2022, 12, 7785. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Luo, J.; Zhu, Z.; Chen, B.; Ye, X.; Zhu, P.; Zhang, B. Multi-scale biosurfactant production by Bacillus subtilis using tuna fish waste as substrate. Catalysts 2021, 11, 456. [Google Scholar] [CrossRef]
- Liu, Q.; Niu, J.; Liu, Y.; Li, L.; Lv, J. Optimization of lipopeptide biosurfactant production by Bacillus licheniformis L20 and performance evaluation of biosurfactant mixed system for enhanced oil recovery. J. Pet. Sci. Eng. 2022, 208, 109678. [Google Scholar] [CrossRef]
- Barros, V.P.O.; Silva, J.R.M.; Melo, V.M.M.; Terceiro, P.S.; Oliveira, I.N.; Freitas, J.D.; Moura, O.F.S.; Araújo-Júnior, J.X.; Rodrigues, E.E.S.; Maraschin, M.; et al. Biosurfactants production by marine yeasts isolated from zoanthids and characterization of an emulsifier produced by Yarrowia lipolytica LMS 24B. Chemosphere 2024, 355, 141807. [Google Scholar] [CrossRef]
- Haidar, C.N.; Pereira, M.M.; Lima, Á.S.; Nerli, B.B.; Malpiedi, L.P. Biosurfactants produced by Pseudomonas syringae pv tabaci: A versatile mixture with interesting emulsifying properties. Process Biochem. 2020, 97, 121–129. [Google Scholar] [CrossRef]
- Singh, V.; Waris, Z.; Banat, I.M.; Saha, S.; Padmanabhan, P. Assessment of rheological behaviour of water-in-oil emulsions mediated by glycolipid biosurfactant produced by Bacillus megaterium SPSW1001. Appl. Biochem. Biotechnol. 2022, 194, 1310–1326. [Google Scholar] [CrossRef]
- Maia, P.C.V.S.; Santos, V.S.; Ferreira, A.S.; Luna, M.A.C.; Silvia, T.A.L.; Andrade, R.F.S.; Campos-Takaki, G.M. An Efficient Bioemulsifier-Producing Bacillus subtilis UCP 0146 Isolated from Mangrove Sediments. Colloids Interfaces 2018, 2, 58. [Google Scholar] [CrossRef]
- Gayathiri, E.; Prakash, P.; Karmegam, N.; Varjani, S.; Awasthi, M.K.; Ravindran, B. Biosurfactants: Potential and eco-friendly material for sustainable agriculture and environmental safety—A review. Agronomy 2022, 12, 662. [Google Scholar] [CrossRef]
- Ja’afaru, M.I.; Abbas, T.; Ajunwa, O.M.; Olaifa, K. Characterization and statistical optimization of biosurfactant production using Bacillus subtilis isolated from automotive oil-contaminated soil in Yola, Nigeria. Sci. Afr. 2022, 17, e01357. [Google Scholar] [CrossRef]
- Mnif, I.; Rajhi, H.; Bouallegue, A.; Trabelsi, N.; Ghribi, D. Characterization of lipopeptides biosurfactants produced by a newly isolated strain Bacillus subtilis ZNI5: Potential environmental application. J. Polym. Environ. 2022, 30, 2378–2391. [Google Scholar] [CrossRef]
- Mendes da Silva Santos, E.; Alvares da Silva Lira, I.R.; Moraes Meira, H.; dos Santos Aguiar, J.; Diniz Rufino, R.; Germano de Almeida, D.; Casazza, A.A.; Converti, A.; Asfora Sarubbo, L.; Moura de Luna, J. Enhanced Oil Removal by a Non-Toxic Biosurfactant Formulation. Energies 2021, 14, 467. [Google Scholar] [CrossRef]
- Ravera, F.; Dziza, K.; Santini, E.; Cristofolini, L.; Liggieri, L. Emulsification and emulsion stability: The role of the interfacial properties. Adv. Colloid Interface Sci. 2021, 288, 102344. [Google Scholar] [CrossRef]
- Jumpathong, W.; Intra, B.; Euanorasetr, J.; Wanapaisan, P. Biosurfactant-producing Bacillus velezensis PW192 as an anti-fungal biocontrol agent against Colletotrichum gloeosporioides and Colletotrichum musae. Microorganisms 2022, 10, 1017. [Google Scholar] [CrossRef]
- Maia, P.C.D.V.S.; Rodríguez, D.M.; Souza, A.F.; Andrade, R.F.S.; Campos-Takaki, G.M. Production of biosurfactant by Bacillus subtilis UCP 0999 using cassava wastewater (CWW) and waste frying oil (WFO) as renewable substrates. Res. Soc. Dev. 2022, 11, e17011628805. [Google Scholar] [CrossRef]
- Nurhasanah; Citra, S.; Bahri, S.; Hendri, J. Production of lipopetide biosurfactant by indigenous bacteria Bacillus sp. ALPD1 using diesel oil as the carbon source. AIP Conf. Proc. 2024, 2970, 060037. [Google Scholar] [CrossRef]
- Umar, A.; Zafar, A.; Wali, H.; Siddique, M.P.; Qazi, M.A.; Naeem, A.H.; Malik, Z.A.; Ahmed, S. Low-cost production and application of lipopeptide for bioremediation and plant growth by Bacillus subtilis SNW3. AMB Express 2021, 11, 165. [Google Scholar] [CrossRef]
- Durval, I.J.B.; Mendonça, A.H.R.; Rocha, I.V.; Luna, J.M.; Rufino, R.D.; Converti, A.; Sarubbo, L.A. Production, characterization, evaluation and toxicity assessment of a Bacillus cereus UCP 1615 biosurfactant for marine oil spills bioremediation. Mar. Pollut. Bull. 2020, 157, 111357. [Google Scholar] [CrossRef]
- Kumari, K.; Behera, H.T.; Nayak, P.P.; Sinha, A.; Nandi, A.; Ghosh, A.; Saha, U.; Suar, M.; Panda, P.K.; Verma, S.K.; et al. Amelioration of lipopeptide biosurfactants for enhanced antibacterial and biocompatibility through molecular antioxidant property by methoxy and carboxyl moieties. Biomed. Pharmacother. 2023, 161, 114493. [Google Scholar] [CrossRef]
- Abdelraof, M.; Nooman, M.U.; Hashem, A.H.; Al-Kashef, A.S. Production and optimization of surfactin produced from locally isolated Bacillus halotolerans grown on agro-industrial wastes and its antimicrobial efficiency. BMC Microbiol. 2024, 24, 193. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Amaki, Y.; Ishihara, A.; Nakajima, H. Synergistic effects of [Ile7] surfactin homologues with bacillomycin D in suppression of gray mold disease by Bacillus amyloliquefaciens biocontrol strain SD-32. J. Agric. Food Chem. 2015, 63, 5344–5353. [Google Scholar] [CrossRef] [PubMed]
- de Souza, E.M.C.; de Oliveira, M.V.D.; Siqueira, J.E.S.; Rocha, D.C.D.C.; Marinho, A.D.N.R.; Marinho, A.M.D.R.; Marinho, P.S.B.; Lima, A.H. Molecular characterization and in silico evaluation of surfactins produced by endophytic bacteria from Phanera splendens. Front. Chem. 2023, 11, 1240704. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Xie, W.; Liang, Y.; Luo, G.; Li, L.; Zheng, W.; Xu, Q.; Xu, H. Algicidal characteristics of novel algicidal compounds, cyclic lipopeptide surfactins from Bacillus tequilensis strain D8, in eliminating Heterosigma akashiwo blooms. Front. Microbiol. 2022, 13, 1066747. [Google Scholar] [CrossRef]
- de Faria, A.F.; Teodoro-Martinez, D.S.; de Oliveira Barbosa, G.N.; Vaz, B.G.; Silva, Í.S.; Garcia, J.S.; Tótola, M.R.; Eberlin, M.N.; Grossman, M.; Alves, O.L.; et al. Production and structural characterization of surfactin (C14/Leu7) produced by Bacillus subtilis isolate LSFM-05 grown on raw glycerol from the biodiesel industry. Process Biochem. 2011, 46, 1951–1957. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, S.; Zhang, S.; Wu, G.; Shao, Y.; Mi, Q.; Liang, J.; Sun, K.; Hu, J. Isolation and characterization of a new cyclic lipopeptide surfactin from a marine-derived Bacillus velezensis SH-B74. J. Antibiot. 2020, 73, 863–867. [Google Scholar] [CrossRef]
- Silva, I.G.S.; de Almeida, F.C.G.; da Rocha e Silva, N.M.P.; de Oliveira, J.T.R.; Converti, A.; Sarubbo, L.A. Application of green surfactants in the remediation of soils contaminated by hydrocarbons. Processes 2021, 9, 1666. [Google Scholar] [CrossRef]
- Brai, A.; Poggialini, F.; Vagaggini, C.; Pasqualini, C.; Simoni, S.; Francardi, V.; Dreassi, E. Tenebrio molitor as a simple and cheap preclinical pharmacokinetic and toxicity model. Int. J. Mol. Sci. 2023, 24, 2296. [Google Scholar] [CrossRef]
- Lima, B.G.A.; Santos, J.C.V.; Silva, R.R.; Caldas, M.C.F.; Meira, H.M.; Rufino, R.D.; Sarubbo, L.A.; Luna, J.M. Sustainable Production of Biosurfactant Grown in Medium with Industrial Waste and Use for Removal of Oil from Soil and Seawater. Surfaces 2024, 7, 537–549. [Google Scholar] [CrossRef]
- Almeida, H.F.; Silva, R.O.; Almeida, M.B.; Almeida, F.; Menezes, J.S.; Dias, T.A.P. Análise do perfil de sensibilidade aos antimicrobianos de um hospital público de Aracaju, Sergipe. Rev. Bras. Anal. Clin. 2021, 53, 277–284. [Google Scholar]
- BrCAST—Brazilian Committee for Sensitivity Testing. Tabelas de pontos de corte para interpretação de CIMs e diâmetros de halos. Available online: https://brcast.org.br/ (accessed on 23 May 2025).
- Yuliani, H.; Perdani, M.S.; Savitri, I.; Manurung, M.; Sahlan, M.; Wijanarko, A.; Hermansyah, H. Antimicrobial activity of biosurfactant derived from Bacillus subtilis C19. Energy Procedia 2018, 153, 274–278. [Google Scholar] [CrossRef]
- Das, R.P.; Sahoo, S.; Paidesetty, S.K.; Ahmad, I.; Sahoo, B.; Jayabaskaran, C.; Patel, H.; Arakha, M.; Pradhan, A.K. Isolation, characterization, and multimodal evaluation of novel glycolipid biosurfactant derived from Bacillus species: A promising Staphylococcus aureus tyrosyl-tRNA synthetase inhibitor through molecular docking and MD simulations. Int. J. Biol. Macromol. 2024, 261, 129848. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, V.; Dhiman, V.K.; Kanwar, S.S. Purification and characterization of a novel bacterial Lipopeptide(s) biosurfactant and determining its antimicrobial and cytotoxic properties. Process Biochem. 2022, 120, 114–125. [Google Scholar] [CrossRef]
- Gharaie, S.; Ohadi, M.; Hassanshahian, M.; Shakibaie, M.; Shahriary, P.; Forootanfar, H. Glycolipopeptide biosurfactant from Bacillus pumilus SG: Physicochemical characterization, optimization, antibiofilm and antimicrobial activity evaluation. 3 Biotech 2023, 13, 321. [Google Scholar] [CrossRef]
- Argentin, M.N.; Martins, L.F.; Sousa, M.P.; Bossolan, N.R.S. Biosurfactant from a thermo-halophilic strain of Bacillus alveayuensis isolated from a Brazilian oil reservoir: Production, chemical characterization, antimicrobial activity, and efficiency in wettability reversal and oil removal from oil-soaked sand. Geoenergy Sci. Eng. 2023, 231, 212324. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Evaluated Sources | Substrate | Concentration | Surface Tension (mN·m−1) of Best Growing Time (120 h) |
|---|---|---|---|
| Glucose | 49.4 ± 0.004 | ||
| Molasses | 2% | 47.1 ± 0.002 | |
| Carbon | Soybean oil | 40.5 ± 0.012 | |
| Residual soybean oil | 43.9 ± 0.009 | ||
| Corn steep liquor | 34.3 ± 0.008 | ||
| Urea | 41.6 ± 0.003 | ||
| Nitrogen | Peptone | 0.12% | 43.6 ± 0.002 |
| Sodium nitrate | 47.5 ± 0.009 | ||
| Ammonium chloride | 32.0 ± 0.004 |
| Temperature (°C) | Surface Tension mN·m−1 | pH | Surface Tension mN·m−1 | NaCl (%) | Surface Tension mN·m−1 |
|---|---|---|---|---|---|
| 5 | 31.9 ± 0.09 | 2 | 30.7 ± 0.08 | 2 | 27.9 ± 0.06 |
| 50 | 26.0 ± 0.05 | 4 | 27.7 ± 0.05 | 4 | 27.4 ± 0.07 |
| 70 | 27.7 ± 0.08 | 6 | 30.8 ± 0.02 | 6 | 27.4 ± 0.04 |
| 100 | 26.2 ± 0.05 | 8 | 26.8 ± 0.05 | 8 | 27.8 ± 0.08 |
| 10 | 29.5 ± 0.07 | 10 | 27.4 ± 0.08 | ||
| 12 | 28.4 ± 0.03 | 12 | 27.7 ± 0.04 |
| pH | Motor Oil (%) | Soybean Oil (%) | Sunflower Oil (%) |
|---|---|---|---|
| 2 | 100 ± 0.01 | 50 ± 0.03 | 35 ± 0.01 |
| 4 | 100 ± 0.02 | 55 ± 0.02 | 40 ± 0.02 |
| 6 | 100 ± 0.02 | 55 ± 0.02 | 40 ± 0.02 |
| 8 | 100 ± 0.01 | 55 ± 0.01 | 40 ± 0.02 |
| 10 | 100 ± 0.01 | 55 ± 0.02 | 40 ± 0.01 |
| 12 | 100 ± 0.02 | 55 ± 0.01 | 40 ± 0.02 |
| Temperature (°C) | Motor Oil (%) | Soybean Oil (%) | Sunflower Oil (%) |
|---|---|---|---|
| 5 | 100 ± 0.02 | 60 ± 0.03 | 35 ± 0.04 |
| 50 | 100 ± 0.01 | 55 ± 0.02 | 50 ± 0.03 |
| 70 | 100 ± 0.01 | 60 ± 0.01 | 55 ± 0.03 |
| 100 | 100 ± 0.02 | 60 ± 0.02 | 50 ± 0.02 |
| NaCl (%) | Motor Oil (%) | Soybean Oil (%) | Sunflower Oil (%) |
|---|---|---|---|
| 2 | 100 ± 0.03 | 40 ± 0.04 | 30 ± 0.01 |
| 4 | 100 ± 0.01 | 40 ± 0.05 | 30 ± 0.03 |
| 6 | 100 ± 0.02 | 40 ± 0.04 | 35 ± 0.02 |
| 8 | 100 ± 0.02 | 40 ± 0.04 | 30 ± 0.01 |
| 10 | 100 ± 0.01 | 45 ± 0.02 | 30 ± 0.04 |
| 12 | 100 ± 0.02 | 45 ± 0.04 | 30 ± 0.03 |
| Concentration of Biosurfactant | Tomato (Solanum lycopersicum) (%) | Lettuce (Lactuca sativa) (%) |
|---|---|---|
| Distilled water | 90 | 85 |
| ½ CMC | 80 | 83 |
| 1 CMC | 90 | 87 |
| 2 CMC | 80 | 90 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albuquerque, A.P.d.C.; Ferreira, H.d.S.; Silva, Y.A.d.; Silva, R.R.d.; Lima, C.V.A.d.; Sarubbo, L.A.; Luna, J.M. Biosurfactant Produced by Bacillus subtilis UCP 1533 Isolated from the Brazilian Semiarid Region: Characterization and Antimicrobial Potential. Microorganisms 2025, 13, 1548. https://doi.org/10.3390/microorganisms13071548
Albuquerque APdC, Ferreira HdS, Silva YAd, Silva RRd, Lima CVAd, Sarubbo LA, Luna JM. Biosurfactant Produced by Bacillus subtilis UCP 1533 Isolated from the Brazilian Semiarid Region: Characterization and Antimicrobial Potential. Microorganisms. 2025; 13(7):1548. https://doi.org/10.3390/microorganisms13071548
Chicago/Turabian StyleAlbuquerque, Antônio P. da C., Hozana de S. Ferreira, Yali A. da Silva, Renata R. da Silva, Carlos V. A. de Lima, Leonie A. Sarubbo, and Juliana M. Luna. 2025. "Biosurfactant Produced by Bacillus subtilis UCP 1533 Isolated from the Brazilian Semiarid Region: Characterization and Antimicrobial Potential" Microorganisms 13, no. 7: 1548. https://doi.org/10.3390/microorganisms13071548
APA StyleAlbuquerque, A. P. d. C., Ferreira, H. d. S., Silva, Y. A. d., Silva, R. R. d., Lima, C. V. A. d., Sarubbo, L. A., & Luna, J. M. (2025). Biosurfactant Produced by Bacillus subtilis UCP 1533 Isolated from the Brazilian Semiarid Region: Characterization and Antimicrobial Potential. Microorganisms, 13(7), 1548. https://doi.org/10.3390/microorganisms13071548