

Step-Wise Ethanol Adaptation Drives Cell-Wall Remodeling and ROM2/KNR4 Activation in Brettanomyces bruxellensis

 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Media and Growth Conditions
2.2. Cell Wall Susceptibility Analysis
2.3. Transmission Electron Microscopy (TEM)
2.4. Polysaccharide Composition of the Cell Wall
2.5. Evaluation of Some Genes Overexpressed in B. bruxellensis Under Different Ethanol Concentrations
2.6. RNA Extraction
2.7. Relative Expression Quantification
2.8. Statistical Analysis
3. Results
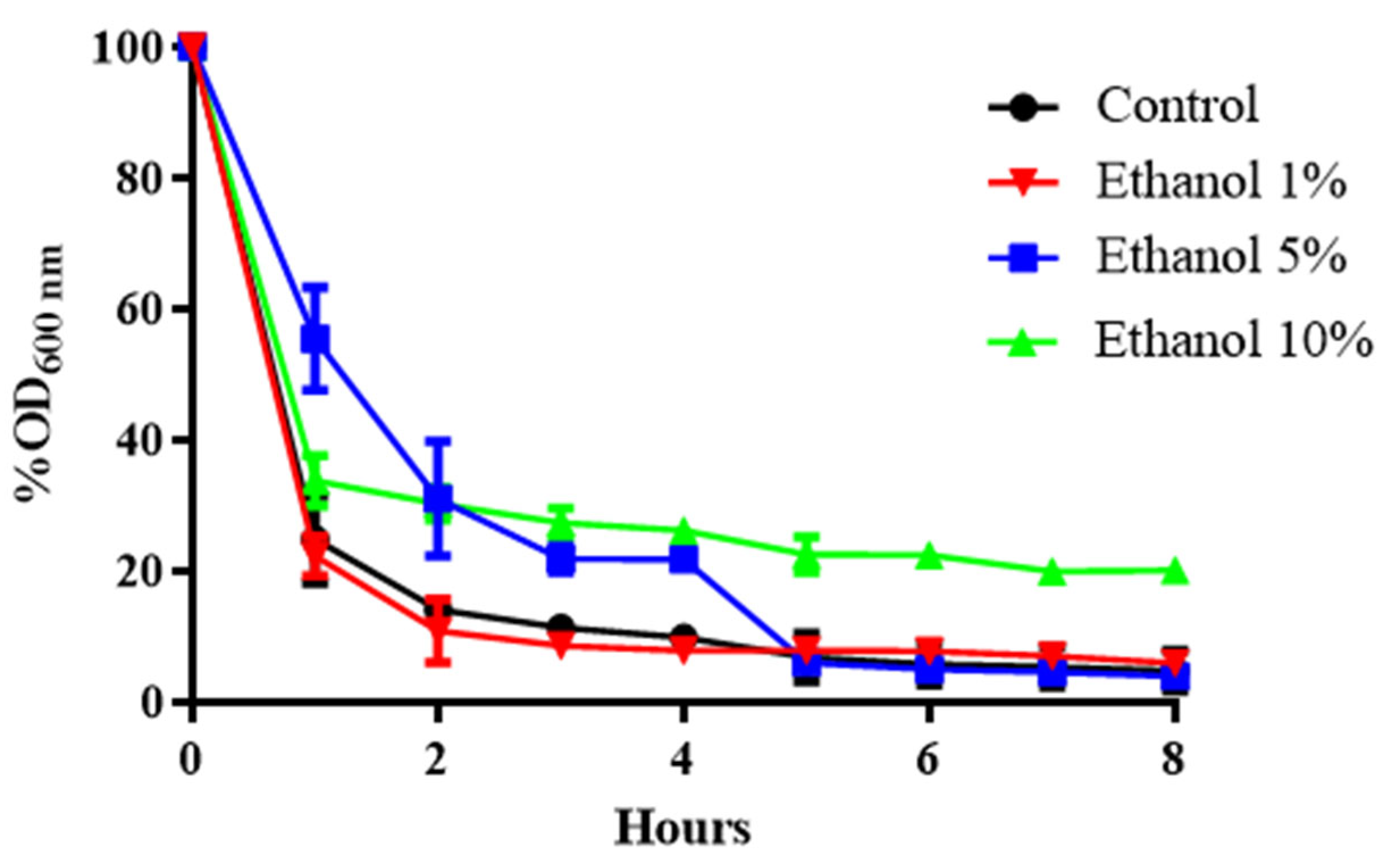
3.1. Effect of the Zymolyase Enzyme on the Cell Wall of B. bruxellensis Grown at Increasing Ethanol Concentrations
3.2. Analysis of the Cell Wall by TEM
3.3. Determination of Cell Wall Polysaccharide Concentration
3.4. Obtaining the orthologous genes KNR4/SMI1, ROM2 and SED1 from S. cerevisiae in B. bruxellensis
3.5. Gene Expression Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Claussen, N.H. On a Method for the Application of Hansen’s Pure Yeast System in the Manufacturing of Well-Conditioned English Stock Beers. J. Inst. Brew. 1904, 10, 308–331. [Google Scholar] [CrossRef]
- Harris, V.; Ford, C.M.; Jiranek, V.; Grbin, P.R. Dekkera and Brettanomyces Growth and Utilisation of Hydroxycinnamic Acids in Synthetic Media. Appl. Microbiol. Biotechnol. 2008, 78, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Da-Silva, E.; Surribas, A.; Gambari, C.; Granès, D.; Pic, L.; Barthelmebs, L. Development of a Molecular Diagnostic Test for the Specific Detection of Brettanomyces bruxellensis in Red Wine. Int. J. Food Microbiol. 2023, 407, 110394. [Google Scholar] [CrossRef] [PubMed]
- Schumaker, M.R.; Diako, C.; Castura, J.C.; Edwards, C.G.; Ross, C.F. Influence of Wine Composition on Consumer Perception and Acceptance of Brettanomyces Metabolites Using Temporal Check-All-That-Apply Methodology. Food Res. Int. 2019, 116, 963–972. [Google Scholar] [CrossRef]
- Miranda, J.; Miot-Sertier, C.; Olazabal, L.; Albertin, W.; Richard, T.; Da Costa, G.; Rouger, C.; Dols-Lafargue, M. Bordeaux Red Wines Display Variable Intrinsic Ability to Support Brettanomyces bruxellensis Growth. Food Control 2024, 155, 110067. [Google Scholar] [CrossRef]
- Chandra, M.; Branco, P.; Prista, C.; Malfeito-Ferreira, M. Role of P-Coumaric Acid and Micronutrients in Sulfur Dioxide Tolerance in Brettanomyces bruxellensis. Beverages 2023, 9, 69. [Google Scholar] [CrossRef]
- Louw, M.; du Toit, M.; Alexandre, H.; Divol, B. Comparative Morphological Characteristics of Three Brettanomyces bruxellensis Wine Strains in the Presence/absence of Sulfur Dioxide. Int. J. Food Microbiol. 2016, 238, 79–88. [Google Scholar] [CrossRef]
- Serpaggi, V.; Remize, F.; Recorbet, G.; Gaudot-Dumas, E.; Sequeira-Le Grand, A.; Alexandre, H. Characterization of the “Viable but Nonculturable” (VBNC) State in the Wine Spoilage Yeast Brettanomyces. Food Microbiol. 2012, 30, 438–447. [Google Scholar] [CrossRef]
- Oelofse, A.; Pretorius, I.S.; Du Toit, M. Significance of Brettanomyces and Dekkera during Winemaking: A Synoptic Review. S. Afr. J. Enol. Vitic. 2008, 29, 128–144. [Google Scholar] [CrossRef]
- Paschos, T.; Xiros, C.; Christakopoulos, P. Simultaneous Saccharification and Fermentation by Co-Cultures of Fusarium oxysporum and Saccharomyces cerevisiae Enhances Ethanol Production from Liquefied Wheat Straw at High Solid Content. Ind. Crops Prod. 2015, 76, 793–802. [Google Scholar] [CrossRef]
- Sahana, G.R.; Balasubramanian, B.; Joseph, K.S.; Pappuswamy, M.; Liu, W.-C.; Meyyazhagan, A.; Kamyab, H.; Chelliapan, S.; Joseph, B.V. A Review on Ethanol Tolerance Mechanisms in Yeast: Current Knowledge in Biotechnological Applications and Future Directions. Process Biochem. 2024, 138, 1–13. [Google Scholar] [CrossRef]
- Klis, F.M.; Boorsma, A.; De Groot, P.W.J. Cell Wall Construction in Saccharomyces cerevisiae. Yeast 2006, 23, 185–202. [Google Scholar] [CrossRef]
- Batista, M.; Donker, E.I.M.; Bon, C.; Guillien, M.; Caisso, A.; Mourey, L.; François, J.-M.; Maveyraud, L.; Zerbib, D. The Conserved Yeast Protein Knr4 Involved in Cell Wall Integrity Is a Multi-Domain Intrinsically Disordered Protein. J. Mol. Biol. 2023, 435, 168048. [Google Scholar] [CrossRef] [PubMed]
- Sanz, A.B.; García, R.; Rodríguez-Peña, J.M.; Arroyo, J. The CWI Pathway: Regulation of the Transcriptional Adaptive Response to Cell Wall Stress in Yeast. J. Fungi 2017, 4, 1. [Google Scholar] [CrossRef]
- Ibe, C.; Munro, C.A. Fungal Cell Wall Proteins and Signaling Pathways Form a Cytoprotective Network to Combat Stresses. J. Fungi 2021, 7, 739. [Google Scholar] [CrossRef]
- Li, X.; Ohmori, T.; Irie, K.; Kimura, Y.; Suda, Y.; Mizuno, T.; Irie, K. Different Regulations of and Expression by Ccr4, Pop2, and Dhh1 in the Cell Wall Integrity Pathway. mSphere 2016, 1, e00250-16. [Google Scholar] [CrossRef]
- Bakir, G.; Dahms, T.E.S.; Martin-Yken, H.; Bechtel, H.A.; Gough, K.M. CellWall Remodeling in the Absence of Knr4 and Kre6 Revealed by Nano-FourierTransform Infrared Spectroscopy. Appl. Spectrosc. 2024, 78, 355–364. [Google Scholar] [CrossRef]
- Schifferdecker, A.J.; Dashko, S.; Ishchuk, O.P.; Piškur, J. The Wine and Beer Yeast Dekkera bruxellensis. Yeast 2014, 31, 323–332. [Google Scholar] [CrossRef]
- Le Montagner, P.; Bakhtiar, Y.; Miot-Sertier, C.; Guilbaud, M.; Albertin, W.; Moine, V.; Dols-Lafargue, M.; Masneuf-Pomarède, I. Effect of Abiotic and Biotic Factors on Brettanomyces bruxellensis Bioadhesion Properties. Food Microbiol. 2024, 120, 104480. [Google Scholar] [CrossRef]
- Di Canito, A.; Foschino, R.; Vigentini, I. Flocculation Mechanisms in: Influence of Ethanol and Sulfur Dioxide on Gene Expression. Curr. Res. Microb. Sci. 2025, 8, 100372. [Google Scholar] [CrossRef]
- Coronado, P.; Aguilera, S.; Carmona, L.; Godoy, L.; Martínez, C.; Ganga, M.A. Comparison of the Behaviour of Brettanomyces bruxellensis Strain LAMAP L2480 Growing in Authentic and Synthetic Wines. Antonie Van. Leeuwenhoek 2015, 107, 1217–1223. [Google Scholar] [CrossRef] [PubMed]
- Shimoi, H.; Kitagaki, H.; Ohmori, H.; Iimura, Y.; Ito, K. Sed1p Is a Major Cell Wall Protein of Saccharomyces cerevisiae in the Stationary Phase and Is Involved in Lytic Enzyme Resistance. J. Bacteriol. 1998, 180, 3381–3387. [Google Scholar] [CrossRef] [PubMed]
- Mora-Montes, H.M.; Bates, S.; Netea, M.G.; Díaz-Jiménez, D.F.; López-Romero, E.; Zinker, S.; Ponce-Noyola, P.; Kullberg, B.J.; Brown, A.J.P.; Odds, F.C.; et al. Endoplasmic Reticulum Alpha-Glycosidases of Candida albicans Are Required for N Glycosylation, Cell Wall Integrity, and Normal Host-Fungus Interaction. Eukaryot. Cell 2007, 6, 2184–2193. [Google Scholar] [CrossRef]
- Fleet, G.H. Composition and Structure of Yeast Cell Walls. Curr. Top. Med. Mycol. 1985, 1, 24–56. [Google Scholar] [CrossRef]
- Godoy, L.; Vera-Wolf, P.; Martinez, C.; Ugalde, J.A.; Ganga, M.A. Comparative Transcriptome Assembly and Genome-Guided Profiling for Brettanomyces bruxellensis LAMAP2480 during P-Coumaric Acid Stress. Sci. Rep. 2016, 6, 34304. [Google Scholar] [CrossRef]
- Nardi, T.; Remize, F.; Alexandre, H. Adaptation of Yeasts Saccharomyces cerevisiae and Brettanomyces bruxellensis to Winemaking Conditions: A Comparative Study of Stress Genes Expression. Appl. Microbiol. Biotechnol. 2010, 88, 925–937. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Svec, D.; Tichopad, A.; Novosadova, V.; Pfaffl, M.W.; Kubista, M. How Good Is a PCR Efficiency Estimate: Recommendations for Precise and Robust qPCR Efficiency Assessments. Biomol. Detect. Quantif. 2015, 3, 9–16. [Google Scholar] [CrossRef]
- Arroyo, J.; Bermejo, C.; García, R.; Rodríguez-Peña, J.M. Genomics in the Detection of Damage in Microbial Systems: Cell Wall Stress in Yeast. Clin. Microbiol. Infect. 2009, 15 (Suppl. S1), 44–46. [Google Scholar] [CrossRef]
- Rodríguez-Peña, J.M.; Díez-Muñiz, S.; Bermejo, C.; Nombela, C.; Arroyo, J. Activation of the Yeast Cell Wall Integrity MAPK Pathway by Zymolyase Depends on Protease and Glucanase Activities and Requires the Mucin-like Protein Hkr1 but Not Msb2. FEBS Lett. 2013, 587, 3675–3680. [Google Scholar] [CrossRef]
- Schiavone, M.; François, J.M.; Zerbib, D.; Capp, J.-P. Emerging Relevance of Cell Wall Components from Non-Conventional Yeasts as Functional Ingredients for the Food and Feed Industry. Curr. Res. Food Sci. 2023, 7, 100603. [Google Scholar] [CrossRef] [PubMed]
- Birch, R.M.; Walker, G.M. Influence of Magnesium Ions on Heat Shock and Ethanol Stress Responses of Saccharomyces cerevisiae. Enzym. Microb. Technol. 2000, 26, 678–687. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.C.T.; Schneper, L.; Russo, M.; Fernandes, A.A.R.; Broach, J.R.; Fernandes, P.M.B. Comparative Transcriptome Analysis of Industrial and Laboratory Saccharomyces cerevisiae Strains after Sequential Stresses. Fermentation 2024, 10, 395. [Google Scholar] [CrossRef]
- Takahashi, T.; Shimoi, H.; Ito, K. Identification of Genes Required for Growth under Ethanol Stress Using Transposon Mutagenesis in Saccharomyces cerevisiae. Mol. Genet. Genom. 2001, 265, 1112–1119. [Google Scholar] [CrossRef]
- Woolfit, M.; Rozpedowska, E.; Piskur, J.; Wolfe, K.H. Genome Survey Sequencing of the Wine Spoilage Yeast Dekkera (Brettanomyces) Bruxellensis. Eukaryot. Cell 2007, 6, 721–733. [Google Scholar] [CrossRef]
- Schiavone, M.; Formosa-Dague, C.; Elsztein, C.; Teste, M.-A.; Martin-Yken, H.; De Morais, M.A., Jr.; Dague, E.; François, J.M. Evidence for a Role for the Plasma Membrane in the Nanomechanical Properties of the Cell Wall as Revealed by an Atomic Force Microscopy Study of the Response of Saccharomyces cerevisiae to Ethanol Stress. Appl. Environ. Microbiol. 2016, 82, 4789–4801. [Google Scholar] [CrossRef]
- Cansado, J.; Soto, T.; Franco, A.; Vicente-Soler, J.; Madrid, M. The Fission Yeast Cell Integrity Pathway: A Functional Hub for Cell Survival upon Stress and Beyond. J. Fungi 2021, 8, 32. [Google Scholar] [CrossRef]
- Teixeira, M.C.; Raposo, L.R.; Mira, N.P.; Lourenço, A.B.; Sá-Correia, I. Genome-Wide Identification of Saccharomyces cerevisiae Genes Required for Maximal Tolerance to Ethanol. Appl. Environ. Microbiol. 2009, 75, 5761–5772. [Google Scholar] [CrossRef]
- Aguilar-Uscanga, B.; François, J.M. A Study of the Yeast Cell Wall Composition and Structure in Response to Growth Conditions and Mode of Cultivation. Lett. Appl. Microbiol. 2003, 37, 268–274. [Google Scholar] [CrossRef]
- Schiavone, M.; Vax, A.; Formosa, C.; Martin-Yken, H.; Dague, E.; François, J.M. A Combined Chemical and Enzymatic Method to Determine Quantitatively the Polysaccharide Components in the Cell Wall of Yeasts. FEMS Yeast Res. 2014, 14, 933–947. [Google Scholar] [CrossRef]
- Orlean, P. Architecture and Biosynthesis of the Saccharomyces cerevisiae Cell Wall. Genetics 2012, 192, 775–818. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, R.A.; Bourbon-Melo, N.; Sá-Correia, I. The Cell Wall and the Response and Tolerance to Stresses of Biotechnological Relevance in Yeasts. Front. Microbiol. 2022, 13, 953479. [Google Scholar] [CrossRef] [PubMed]
- Bekirian, C.; Valsecchi, I.; Bachellier-Bassi, S.; Scandola, C.; Guijarro, J.I.; Chauvel, M.; Mourer, T.; Gow, N.A.R.; Aimanianda, V.K.; d’Enfert, C.; et al. β-1,6-Glucan Plays a Central Role in the Structure and Remodeling of the Bilaminate Fungal Cell Wall. Elife 2024, 13, 100569. [Google Scholar] [CrossRef]
- Valdivia, R.H.; Schekman, R. The Yeasts Rho1p and Pkc1p Regulate the Transport of Chitin Synthase III (Chs3p) from Internal Stores to the Plasma Membrane. Proc. Natl. Acad. Sci. USA 2003, 100, 10287–10292. [Google Scholar] [CrossRef]
- Piper, P.W. The Heat Shock and Ethanol Stress Responses of Yeast Exhibit Extensive Similarity and Functional Overlap. FEMS Microbiol. Lett. 1995, 134, 121–127. [Google Scholar] [CrossRef]
- Sekiya-Kawasaki, M.; Abe, M.; Saka, A.; Watanabe, D.; Kono, K.; Minemura-Asakawa, M.; Ishihara, S.; Watanabe, T.; Ohya, Y. Dissection of Upstream Regulatory Components of the Rho1p Effector, 1,3-Beta-Glucan Synthase, in Saccharomyces cerevisiae. Genetics 2002, 162, 663–676. [Google Scholar] [CrossRef]
- Gasch, A.P.; Spellman, P.T.; Kao, C.M.; Carmel-Harel, O.; Eisen, M.B.; Storz, G.; Botstein, D.; Brown, P.O. Genomic Expression Programs in the Response of Yeast Cells to Environmental Changes. Mol. Biol. Cell 2000, 11, 4241–4257. [Google Scholar] [CrossRef]
- Durand, F.; Dagkessamanskaia, A.; Martin-Yken, H.; Graille, M.; Van Tilbeurgh, H.; Uversky, V.N.; François, J.M. Structure-Function Analysis of Knr4/Smi1, a Newly Member of Intrinsically Disordered Proteins Family, Indispensable in the Absence of a Functional PKC1-SLT2 Pathway in Saccharomyces cerevisiae. Yeast 2008, 25, 563–576. [Google Scholar] [CrossRef]
- Lewis, J.A.; Broman, A.T.; Will, J.; Gasch, A.P. Genetic Architecture of Ethanol-Responsive Transcriptome Variation in Saccharomyces cerevisiae Strains. Genetics 2014, 198, 369–382. [Google Scholar] [CrossRef]
- Arnthong, J.; Ponjarat, J.; Bussadee, P.; Deenarn, P.; Prommana, P.; Phienluphon, A.; Charoensri, S.; Champreda, V.; Zhao, X.-Q.; Suwannarangsee, S. Enhanced Surface Display Efficiency of β-Glucosidase in Saccharomyces cerevisiae by Disruption of Cell Wall Protein-Encoding Genes YGP1 and CWP2. Biochem. Eng. J. 2022, 179, 108305. [Google Scholar] [CrossRef]
- Chandler, M.; Stanley, G.A.; Rogers, P.; Chambers, P. A Genomic Approach to Defining the Ethanol Stress Response in the Yeast Saccharomyces cerevisiae. 2004, 54, 427–454. 54.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′–3′) | Tm (°C) | Amplicon Length (bp) |
|---|---|---|---|
| KNR4/SMI1 | F: CCTTGCAGGAGCAGAAATAC | 55.0 | 103 |
| R: CTACCACTATCTTCGCCCTT | |||
| SED1 | F: CCACTGCTATCCCAACTAAC | 50.5 | 110 |
| R: GGTTGGAGCTTCAGTAGTAG | |||
| ROM2 | F ATGGAGCAGGTCAAGTTATGG | 54.7 | 99 |
| R: CAGCCCTGTTGGATGTATCTT |
| Gen | KNR4/SMI1 | ROM2 | SED1 |
|---|---|---|---|
| Description | Contig00356 | Contig00135 | Contig01043 |
| Max score | 177 | 786 | 39.3 |
| Total score | 177 | 786 | 78.9 |
| Query cover | 67% | 69% | 20% |
| E value | 1 × 10−47 | 00.0 | 0.001 |
| Percent identity | 30.62% | 45.08% | 82.61% |
| Accession length | 46050 | 15345 | 10582 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernandez-Cabello, L.; Rojas-Torres, N.; Godoy, L.; G-Poblete, C.; Concha, Y.; Plaza, V.; Castillo, L.; Mora-Montes, H.M.; Ganga, M.A. Step-Wise Ethanol Adaptation Drives Cell-Wall Remodeling and ROM2/KNR4 Activation in Brettanomyces bruxellensis. Microorganisms 2025, 13, 1489. https://doi.org/10.3390/microorganisms13071489
Hernandez-Cabello L, Rojas-Torres N, Godoy L, G-Poblete C, Concha Y, Plaza V, Castillo L, Mora-Montes HM, Ganga MA. Step-Wise Ethanol Adaptation Drives Cell-Wall Remodeling and ROM2/KNR4 Activation in Brettanomyces bruxellensis. Microorganisms. 2025; 13(7):1489. https://doi.org/10.3390/microorganisms13071489
Chicago/Turabian StyleHernandez-Cabello, Leslie, Nachla Rojas-Torres, Liliana Godoy, Camila G-Poblete, Yarabi Concha, Verónica Plaza, Luis Castillo, Héctor M. Mora-Montes, and María Angélica Ganga. 2025. "Step-Wise Ethanol Adaptation Drives Cell-Wall Remodeling and ROM2/KNR4 Activation in Brettanomyces bruxellensis" Microorganisms 13, no. 7: 1489. https://doi.org/10.3390/microorganisms13071489
APA StyleHernandez-Cabello, L., Rojas-Torres, N., Godoy, L., G-Poblete, C., Concha, Y., Plaza, V., Castillo, L., Mora-Montes, H. M., & Ganga, M. A. (2025). Step-Wise Ethanol Adaptation Drives Cell-Wall Remodeling and ROM2/KNR4 Activation in Brettanomyces bruxellensis. Microorganisms, 13(7), 1489. https://doi.org/10.3390/microorganisms13071489