
Bioaugmentation with Plant Growth-Promoting Rhizobacteria Alleviates Chromium and Salt Stress in Rice Through the Improvement of Physiology, Ion Homeostasis, and Antioxidant Defense

 , , ,
, , ,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Preparation of Bacterial Inocula, Treatments, and Design
2.3. Measurements of Crop Growth Attributes
2.4. Calculation of NaCl and Cr Stress Tolerance Index
2.5. Measurements of Physiological and Biochemical Attributes
2.5.1. Determination of Relative Water Content
2.5.2. Determination of Proline
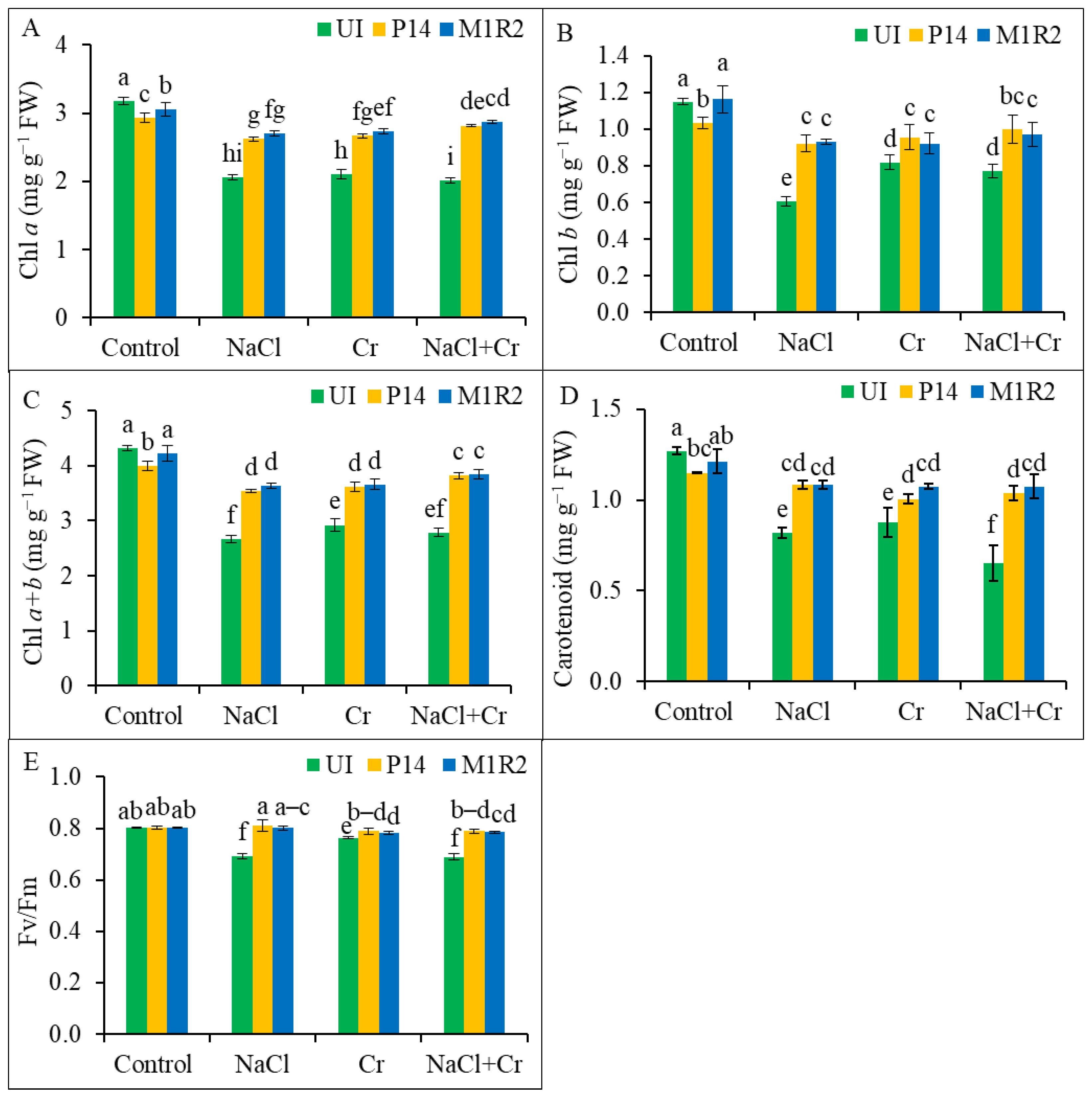
2.5.3. Measurement of Chlorophyll Fluorescence
2.5.4. Determination of Photosynthetic Pigments
2.5.5. Determination of Total Na+ and K+
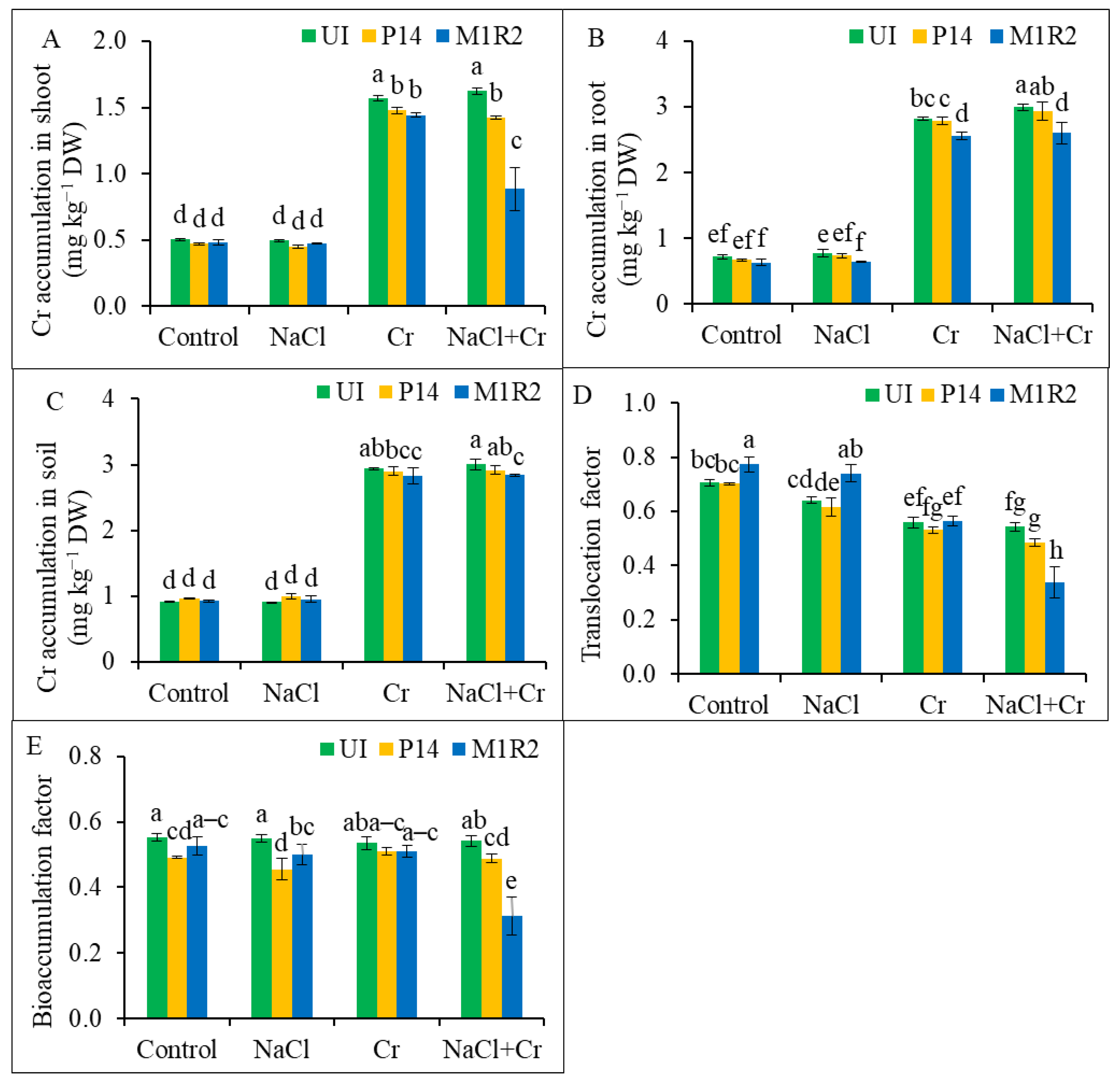
2.5.6. Cr Analysis in Shoot, Root, and Soil
2.6. Determination of Oxidative Stress Indicators
2.6.1. Determination of Lipid Peroxidation
2.6.2. Determination of Hydrogen Peroxide
2.6.3. Determination of Electrolyte Leakage
2.7. Enzyme Extraction and Estimation of Free Protein
2.8. Estimation of Antioxidant Enzyme Activity
2.9. Determination of Glyoxalase Enzyme Activity
2.10. Statistical Analysis
3. Results
3.1. Effect of HMT-PGPR Inoculation on Plant Growth and Biomass Accumulation
3.2. Phenotypic Appearances
3.3. Effect of HMT-PGPR Inoculation on Photosynthetic Pigments and Chlorophyll Fluorescence
3.4. Effect of HMT-PGPR Inoculation on Relative Water Content and Osmoregulation
3.5. Effect of HMT-PGPR Inoculation on Oxidative Stress Markers
3.6. Effect of HMT-PGPR Inoculation on Ion Homeostasis
3.7. Evaluation of Inoculation of HMT-PGPR on Antioxidant Enzyme Activities
3.7.1. Antioxidant Enzyme Activities
3.7.2. Glyoxalase System
3.8. Effects of HMT-PGPR Inoculation on Cr Accumulation, Cr Translocation Factor, and Bioaccumulation Factor
3.9. Principal Component Analysis
4. Discussion
4.1. Inoculation of HMT-PGPR Improves Plant Growth and Biomass
4.2. Inoculation of HMT-PGPR Improves Photosynthetic Attributes
4.3. Inoculation of HMT-PGPR Improves Relative Water Content and Osmotic Balance
4.4. Inoculation of HMT-PGPR Reduced Oxidative Damage
4.5. Inoculation of HMT-PGPR Improves Ion Homeostasis
4.6. Inoculation of HMT-PGPR Promotes the Modulation of Antioxidant Defense System
4.7. Inoculation of HMT-PGPR Reduces Cr Concentrations and Its Bioaccumulation
4.8. Response to Epiphytic and Endophytic Strains
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ullah, H.; Santiago-Arenas, R.; Ferdous, Z.; Attia, A.; Datta, A. Improving water use efficiency, nitrogen use efficiency, and radiation use efficiency in field crops under drought stress: A review. Adv. Agron. 2019, 156, 109–157. [Google Scholar]
- Chung, Y.S.; Kim, K.S.; Hamayun, M.; Kim, Y. Silicon confers soybean resistance to salinity stress through regulation of reactive oxygen and reactive nitrogen species. Front. Plant Sci. 2019, 10, 1725. [Google Scholar] [CrossRef]
- Phour, M.; Sindhu, S.S. Soil Salinity and Climate Change: Microbiome-Based Strategies for Mitigation of Salt Stress to Sustainable Agriculture BT. In Climate Change and Microbiome Dynamics: Carbon Cycle Feedbacks; Parray, J.A., Ed.; Springer International Publishing: Cham, Switzerland, 2023; pp. 191–243. [Google Scholar]
- Jamil, A.; Riaz, S.; Ashraf, M.; Foolad, M.R. Gene expression profiling of plants under salt stress. Crit. Rev. Plant Sci. 2011, 30, 435–458. [Google Scholar] [CrossRef]
- Xiaoqin, S.; Dongli, S.; Yuanhang, F.; Hongde, W.; Lei, G. Three-dimensional fractal characteristics of soil pore structure and their relationships with hydraulic parameters in biochar-amended saline soil. Soil Till. Res. 2021, 205, 104809. [Google Scholar] [CrossRef]
- Hashem, A.; Abd_Allah, E.F.; Alqarawi, A.A.; Al-Huqail, A.A.; Shah, M.A. Induction of osmoregulation and modulation of salt stress in Acacia gerrardii Benth. by arbuscular mycorrhizal fungi and Bacillus subtilis (BERA 71). Biomed. Res. Int. 2016, 2016, 6294098. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Bhuyan, M.B.; Oku, H.; Fujita, M. Exogenous nitric oxide pretreatment protects Brassica napus L. seedlings from paraquat toxicity through the modulation of antioxidant defense and glyoxalase systems. Plant Physiol. Biochem. 2018, 126, 173–186. [Google Scholar] [CrossRef]
- Pan, J.; Peng, F.; Xue, X.; You, Q.; Zhang, W.; Wang, T. The growth promotion of two salt-tolerant plant groups with PGPR inoculation: A meta-analysis. Sustainability 2019, 11, 378. [Google Scholar] [CrossRef]
- Krivokapic, M. Study on the evaluation of (heavy) metals in water and sediment of skadar lake (Montenegro), with BCF assessment and translocation ability (TA) by Trapa natans and a review of SDGs. Water 2021, 13, 876. [Google Scholar] [CrossRef]
- Alam, R.; Rasheed, R.; Ashraf, M.A.; Hussain, I.; Ali, S. Allantoin alleviates chromium phytotoxic effects on wheat by regulating osmolyte accumulation, secondary metabolism, ROS homeostasis and nutrient acquisition. J. Hazard. Mater. 2023, 458, 131920. [Google Scholar] [CrossRef]
- Suresh, G.; Ravichandran, N.; Ramesh, B.; Suresh, A.; Siva, G.V. Isolation and characterization of chromium-tolerant bacteria from chromium containing waste water. Bioremed. Biodiver. Bioavail. 2011, 5, 22–27. [Google Scholar]
- Kaszycki, P.; Gabryś, H.; Appenroth, K.-J.; Jaglarz, A.; Sedziwy, S.; Walczak, T.; Kolochek, H. Exogenously applied sulphate as a tool to investigate transport and reduction of chromate in the duckweed Spirodela polyrhiza. Plant Cell Environ. 2005, 28, 260–268. [Google Scholar] [CrossRef]
- Su, C.Q.; Li, L.Q.; Yang, Z.H.; Chai, L.Y.; Liao, Q.; Shi, Y.; Li, J.W. Cr (VI) reduction in chromium-contaminated soil by indigenous microorganisms under aerobic condition. Trans. Nonferrous Met. Soc. China 2019, 29, 1304–1311. [Google Scholar] [CrossRef]
- Smrithi, A.; Usha, K. Isolation and characterization of chromium removing bacteria from tannery effluent disposal site. Intl. J. Adv. Biotec. Res. 2012, 3, 644–652. [Google Scholar]
- Singh, H.P.; Mahajan, P.; Kaur, S.; Batish, D.R.; Kohli, R.K. Chromium toxicity and tolerance in plants. Environ. Chem. Lett. 2013, 11, 229–254. [Google Scholar] [CrossRef]
- Mizan, A.; Mamun, M.A.H.; Islam, M.S. Metal contamination in soil and vegetables around Savar tannery area, Dhaka, Bangladesh: A preliminary study for risk assessment. Heliyon 2023, 9, e13856. [Google Scholar] [CrossRef]
- Vardharajula, S.; Zulfikar, A.S.; Grover, M.; Reddy, G.; Bandi, V. Drought-tolerant plant growth promoting Bacillus spp., effect on growth, osmolytes, and antioxidant status of maize under drought stress. J. Plant Inter. 2011, 6, 1–14. [Google Scholar]
- Alqarawi, A.A.; Abd_Allah, E.F.; Hashem, A. Alleviation of salt-induced adverse impact via mycorrhizal fungi in Ephedra aphylla Forssk. J. Plant Interact. 2014, 9, 802–810. [Google Scholar] [CrossRef]
- Kumar, V.; Suryakant, P.; Kumar, S.; Kumar, N. Effect of chromium toxicity on plants: A review. Agriways 2016, 4, 107–120. [Google Scholar]
- Ma, N.L.; Lah, W.A.C.; Kadir, N.A.; Mustaqim, M.; Rahmat, Z.; Ahmad, A.; Lam, S.D.; Ismail, M.R. Susceptibility and tolerance of rice crop to salt threat: Physiological and metabolic inspections. PLoS ONE 2018, 13, e0192732. [Google Scholar] [CrossRef]
- Basit, F.; Bhat, J.A.; Han, J.; Guan, Y.; Jan, B.L.; Shakoor, A.; Alansi, S. Screening of rice cultivars for Cr-stress response by using the parameters of seed germination, morpho-physiological and antioxidant analysis. Saudi J. Biol. Sci. 2022, 29, 3918–3928. [Google Scholar] [CrossRef]
- Paul, D.; Lade, H. Plant-growth-promoting rhizobacteria to improve crop growth in saline soils: A review. Agron. Sustain. Dev. 2014, 34, 737–752. [Google Scholar] [CrossRef]
- Agbodjato, N.A.; Babalola, O.O. Promoting sustainable agriculture by exploiting plant growth-promoting rhizobacteria (PGPR) to improve maize and cowpea crops. PeerJ 2024, 12, e16836. [Google Scholar] [CrossRef] [PubMed]
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef] [PubMed]
- Dame, Z.T.; Rahman, M.; Islam, T. Bacilli as sources of agrobiotechnology: Recent advances and future directions. Green Chem. Lett. Rev. 2021, 14, 246–271. [Google Scholar] [CrossRef]
- Efe, D. Potential plant growth-promoting bacteria with heavy metal resistance. Curr. Microbiol. 2020, 77, 3861–3868. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, G.; Huang, Y.; Guo, M.; Song, J.; Zhang, T.; Long, Y.; Wang, B.; Liu, H. A Potential biofertilizer—Siderophilic bacteria isolated from the rhizosphere of Paris polyphylla var. yunnanensis. Front. Microbiol. 2022, 13, 870413. [Google Scholar] [CrossRef] [PubMed]
- Saikia, J.; Sarma, R.K.; Dhandia, R.; Yadav, A.; Bharali, R.; Gupta, V.K.; Saikia, R. Alleviation of drought stress in pulse crops with ACC deaminase producing rhizobacteria isolated from acidic soil of Northeast India. Sci. Rep. 2018, 8, 3560. [Google Scholar] [CrossRef]
- Khan, M.A.; Asaf, S.; Khan, A.L.; Adhikari, A.; Jan, R.; Ali, S.; Imran, M.; Kim, K.-M.; Lee, I.-J. Halotolerant rhizobacterial strains mitigate the adverse effects of NaCl stress in soybean seedlings. BioMed Res. Int. 2019, 2019, 9530963. [Google Scholar] [CrossRef]
- Daraz, U.; Ahmad, I.; Li, Q.S.; Zhu, B.; Saeed, M.F.; Li, Y.; Ma, J.; Wang, X.B. Plant growth promoting rhizobacteria induced metal and salt stress tolerance in Brassica juncea through ion homeostasis. Ecotoxicol. Environ. Saf. 2023, 267, 115657. [Google Scholar] [CrossRef]
- BRRI (Bangladesh Rice Research Institute). Adhunik Dhaner Chash; Bangladesh Rice Research Institute: Gazipur, Bangladesh, 2016; p. 26. [Google Scholar]
- Shetty, G.; Hetrick, D.; Schwat, P. Effects of mycorrhizal fertilizers amendments on zinc tolerance of plants. Environ. Pollut. 1995, 88, 308–314. [Google Scholar] [CrossRef]
- Sairam, R.K.; Rao, K.V.; Srivastava, G.C. Differential response of wheat genotypes to long term salinity stress relation to oxidative stress, antioxidant activity and osmolyte concentration. Plant Sci. 2002, 163, 1037–1046. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teari, D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Yoshida, S.; Forno, D.A.; Cock, J.H.; Gomez, K.A. Laboratory Manual for Physiological Studies of Rice, 3rd ed.; International Rice Research Institute: Manila, Philippines, 1976; pp. 61–66. [Google Scholar]
- Campbell, C.R.; Plank, C.O. Sample preparation. In Plant Analysis Reference Procedures for the Southern Region of the United States; Plank, C.O., Ed.; Southern Cooper, Series Bull; Georgia Cooperative Extension Service: Athens, GA, USA, 1992; pp. 1–12. [Google Scholar]
- Shahabivand, S.; Parvaneh, A.; Aliloo, A.A. Different response of Alyssum montanum and Helianthus annuus to cadmium bioaccumulation mediated by the endophyte fungus Serendipita indica. Acta Ecol. Sin. 2020, 40, 315–322. [Google Scholar] [CrossRef]
- Madhava Rao, K.V.; Sresty, T.V.S. Antioxidative parameters in the seedlings of pigeon pea (Cajanus cajan L. Millapaugh) to Zn and Ni stresses. Plant Sci. 2000, 157, 113–128. [Google Scholar] [CrossRef]
- Yu, C.W.; Murphy, T.M.; Lin, C.H. Hydrogen peroxide induced chilling tolerance in mung beans mediated through ABA independent glutathione accumulation. Funct. Plant Biol. 2003, 30, 955–963. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Ahmed, N.; Saha, T.; Rahman, M.; Rahman, K.; Alam, M.M.; Rohman, M.M.; Nahar, K. Exogenous salicylic acid and kinetin modulate reactive oxygen species metabolism and glyoxalase system to confer waterlogging stress tolerance in soybean (Glycine max L.). Plant Stress 2022, 3, 100057. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hemeda, H.M.; Klein, B.P. Effects of naturally occurring antioxidants on peroxidase activity of vegetable extracts. J. Food Sci. 1990, 55, 184–185. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Elia, A.C.; Galarini, R.; Taticchi, M.I.; Dorr, A.J.M.; Mantilacci, L. Antioxidant responses and bioaccumulation in Ictalurus melas under mercury exposure. Ecotoxicol. Environ. Saf. 2003, 55, 162–167. [Google Scholar] [CrossRef]
- Yadav, S.K.; Singla-Pareek, S.L.; Ray, M.; Reddy, M.K.; Sopory, S.K. Methylglyoxal levels in plants under salinity stress are dependent on glyoxalase I and glutathione. Biochem. Biophys. Res. Commun. 2005, 337, 61–67. [Google Scholar] [CrossRef]
- Principato, G.B.; Rosi, G.; Talesa, V.; Giovanni, E.; Uotila, L. Purification and characterization of two forms of glyoxalase II from the liver and brain of Wistar rats. Biochem. Biophys. Acta 1987, 911, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Vishnupradeep, R.; Bruno, L.B.; Taj, Z.; Karthik, C.; Challabathula, D.; Tripti; Kumar, A.; Freitas, H.; Rajkumar, M. Plant growth promoting bacteria improve growth and phytostabilization potential of Zea mays under chromium and drought stress by altering photosynthetic and antioxidant responses. Environ. Technol. Innov. 2022, 25, 102154. [Google Scholar] [CrossRef]
- Ji, J.; Zhang, J.; Wang, X.; Song, W.; Ma, B.; Wang, R.; Li, T.; Wang, G.; Guan, C.; Gao, X. The alleviation of salt stress on rice through increasing photosynthetic capacity, maintaining redox homeostasis and regulating soil enzyme activities by Enterobacter sp. JIV1 assisted with putrescine. Microb. Res. 2024, 280, 127590. [Google Scholar] [CrossRef] [PubMed]
- Sultana, S.; Paul, S.C.; Parveen, S.; Alam, S.; Rahman, N.; Jannat, B.; Hoque, S.; Rahman, M.T.; Karim, M.M. Isolation and identification of salt-tolerant plant growth-promoting rhizobacteria and its application for rice cultivation under salt stress. Can. J. Microbiol. 2020, 66, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Mallick, I.; Bhattacharyya, C.; Mukherji, S.; Dey, D.; Sarkar, S.C.; Mukhopadhyay, U.K.; Ghosh, A. Effective rhizoinoculation and biofilm formation by arsenic immobilizing halophilic plant growth promoting bacteria (PGPB) isolated from mangrove rhizosphere: A step towards arsenic rhizoremediation. Sci. Total Environ. 2018, 610–611, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, R.K.; Rani, V.; Tuteja, N. Azotobacter vinelandii helps to combat chromium stress in rice by maintaining antioxidant machinery. 3 Biotech 2021, 11, 275. [Google Scholar] [CrossRef]
- Sultana, S.; Alam, S.; Karim, M.M. Screening of siderophore-producing salt-tolerant rhizobacteria suitable for supporting plant growth in saline soils with iron limitation. J. Agric. Food. Res. 2021, 4, 100150. [Google Scholar] [CrossRef]
- Kapadia, C.; Patel, N.; Rana, A.; Vaidya, H.; Alfarraj, S.; Ansari, M.J.; Gafur, A.; Poczai, P.; Sayyed, R.Z. Evaluation of plant growth-promoting and salinity ameliorating potential of halophilic bacteria isolated from saline soil. Front. Plant Sci. 2022, 13, 946217. [Google Scholar] [CrossRef]
- Ning, Z.; Lin, K.; Gao, M.; Han, X.; Guan, Q.; Ji, X.; Yu, S.; Lu, L. Mitigation of salt stress in rice by the halotolerant plant growth-promoting bacterium Enterobacter asburiae D2. J. Xenobiot. 2024, 14, 333–349. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, H.; Nosheen, A.; Naz, R.; Bano, A.; Keyani, R. L-tryptophan-assisted PGPR-mediated induction of drought tolerance in maize (Zea mays L.). J. Plant Interact. 2017, 12, 567–578. [Google Scholar] [CrossRef]
- Jirakkakul, J.; Khoiri, A.N.; Duangfoo, T.; Dulsawat, S.; Sutheeworapong, S.; Petsong, K.; Wattanachaisaereekul, S.; Paenkaew, P.; Tachaleat, A.; Cheevadhanarak, S.; et al. Insights into the genome of Methylobacterium sp. NMS14P, a novel bacterium for growth promotion of maize, chili, and sugarcane. PLoS ONE 2023, 18, e0281505. [Google Scholar] [CrossRef]
- Jhuma, T.A.; Rafeya, J.; Sultana, S.; Rahman, M.T.; Karim, M.M. Isolation of endophytic salt-tolerant plant growth-promoting rhizobacteria from Oryza sativa and evaluation of their plant growth-promoting traits under salinity stress condition. Front. Sustain. Food Syst. 2021, 5, 250. [Google Scholar] [CrossRef]
- Karthik, C.; Oves, M.; Thangabalu, R.; Sharma, R.; Santhosh, S.B.; Indra, A.P. Cellulosimicrobium funkei-like enhances the growth of Phaseolus vulgaris by modulating oxidative damage under chromium (VI) toxicity. J. Adv. Res. 2016, 7, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Syed, A.; Elgorban, A.M.; Bahkali, A.H.; Eswaramoorthy, R.; Iqbal, R.K.; Danish, S. Metal-tolerant and siderophore producing Pseudomonas fluorescence and Trichoderma spp. improved the growth, biochemical features and yield attributes of chickpea by lowering Cd uptake. Sci. Rep. 2023, 13, 4471. [Google Scholar] [CrossRef]
- Alharby, H.F.; Ali, S. Combined role of Fe nanoparticles (Fe NPs) and Staphylococcus aureus L. in the alleviation of chromium stress in rice plants. Life 2022, 12, 338. [Google Scholar] [CrossRef]
- Cappellari, L.D.R.; Banchio, E. Volatile organic compounds produced by Bacillus amyloliquefaciens GB03 ameliorate the effects of salt stress in Mentha piperita principally through acetoin emission. J. Plant Growth Regul. 2020, 39, 764–775. [Google Scholar] [CrossRef]
- Zafar-ul-Hye, M.; Naeem, M.; Danish, S.; Khan, M.J.; Fahad, S.; Datta, R.; Brtnicky, M.; Kintl, A.; Hussain, G.S.; El-Esawi, M.A. Effect of cadmium-tolerant rhizobacteria on growth attributes and chlorophyll contents of bitter gourd under cadmium toxicity. Plants 2020, 9, 1386. [Google Scholar] [CrossRef]
- Siddika, A.; Rashid, A.A.; Khan, S.N.; Khatun, A.; Karim, M.M.; Prasad, P.V.V.; Hasanuzzaman, M. Harnessing plant growth-promoting rhizobacteria, Bacillus subtilis and B. aryabhattai to combat salt stress in rice: A study on the regulation of antioxidant defense, ion homeostasis, and photosynthetic parameters. Front. Plant Sci. 2024, 15, 1419764. [Google Scholar] [CrossRef]
- Tátrai, Z.A.; Sanoubar, R.; Pluhár, Z.; Mancarella, S.; Orsini, F.; Gianquinto, G. Morphological and physiological plant responses to drought stress in Thymus citriodorus. Int. J. Agron. 2016, 2016, 4165750. [Google Scholar] [CrossRef]
- Wang, G.; Li, B.; Peng, D.; Zhao, H.; Lu, M.; Zhang, L.; Li, J.; Zhang, S.; Guan, C.; Ji, J. Combined application of H2S and a plant growth promoting strain JIL321 regulates photosynthetic efficacy, soil enzyme activity and growth-promotion in rice under salt stress. Microbiol. Res. 2022, 256, 126943. [Google Scholar] [CrossRef]
- Tirry, N.; Kouchou, A.; El Omari, B.; Ferioun, M.; Ghachtouli, E.N. Improved chromium tolerance of Medicago sativa by plant growth-promoting rhizobacteria (PGPR). J. Genet. Eng. Biotechnol. 2021, 19, 149. [Google Scholar] [CrossRef] [PubMed]
- Nader, A.A.; Hauka, F.I.A.; Afify, A.H.; El-Sawah, A.M. Drought-tolerant bacteria and arbuscular mycorrhizal fungi mitigate the detrimental effects of drought stress induced by withholding irrigation at critical growth stages of soybean (Glycine max, L.). Microorganisms 2024, 12, 1123. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, L.; Zhang, S.; Li, B.; Li, J.; Wang, X.; Zhang, J.; Guan, C.; Ji, J. The combined use of a plant growth promoting Bacillus sp. strain and GABA promotes the growth of rice under salt stress by regulating antioxidant enzyme system, enhancing photosynthesis and improving soil enzyme activities. Microbiol. Res. 2023, 266, 127225. [Google Scholar] [CrossRef]
- Zhao, Z.J.; Zhang, H.L.; Wang, M.J.; Zhang, X.M.; Li, L.X. Salt stress-related regulation mechanism of intracellular pH and ion homeostasis in plants. Plant Physiol. J. 2020, 56, 337–344. [Google Scholar]
- Arora, N.K.; Fatima, T.; Mishra, J.; Mishra, I.; Verma, S.; Verma, R.; Verma, M.; Bhattacharya, A.; Verma, P.; Mishra, P.; et al. Halo-tolerant plant growth promoting rhizobacteria for improving productivity and remediation of saline soils. J. Adv. Res. 2020, 26, 69–82. [Google Scholar] [CrossRef]
- Asif, S.; Jan, R.; Kim, N.; Asaf, S.; Lubna; Khan, M.A.; Kim, E.G.; Jang, Y.H.; Bhatta, D.; Lee, I.J.; et al. Halotolerant endophytic bacteria alleviate salinity stress in rice (Oryza sativa L.) by modulating ion content, endogenous hormones, the antioxidant system and gene expression. BMC Plant Biol. 2023, 23, 494. [Google Scholar] [CrossRef]
- Akhtar, S.S.; Andersen, M.N.; Naveed, M.; Zahir, Z.A.; Liu, F. Interactive effect of biochar and plant growth-promoting bacterial endophytes on ameliorating salinity stress in maize. Funct. Plant Biol. 2015, 42, 770–781. [Google Scholar] [CrossRef]
- Dobrowolski, R.; Szcze, A.; Czemierska, M.; Jarosz-Wiko, A.A. Studies of cadmium(II), lead(II), nickel(II), cobalt(II) and chromium(VI) sorption on extracellular polymeric substances produced by Rhodococcus opacus and Rhodococcus rhodochrous. Bioresour. Technol. 2017, 225, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Wang, Z.; Sha, W.; Song, S.; Qin, F.; Zhang, W. Role of plant-growth-promoting rhizobacteria in plant machinery for soil heavy metal detoxification. Microorganisms 2024, 12, 700. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, R.; Maqsood, M.F.; Shahbaz, M.; Naz, N.; Zulfiqar, U.; Ali, M.F.; Jamil, M.; Khalid, F.; Ali, Q.; Sabir, M.A.; et al. Exogenous ascorbic acid as a potent regulator of antioxidants, osmo-protectants, and lipid peroxidation in pea under salt stress. BMC Plant Biol. 2024, 24, 247. [Google Scholar] [CrossRef]
- Ahmad, S.; Mfarrej, M.F.B.; El-Esawi, M.A.; Waseem, M.; Alatawi, A.; Nafees, M.; Saleem, M.H.; Rizwan, M.; Yasmeen, T.; Anayat, A.; et al. Chromium-resistant Staphylococcus aureus alleviates chromium toxicity by developing synergistic relationships with zinc oxide nanoparticles in wheat. Ecotoxicol. Environ. Saf. 2024, 230, 113142. [Google Scholar] [CrossRef]
- Ali, S.; Waseem, M.; Hussain, A.; Rizwan, M.; Ahmad, A.; Khan, N. Combined application of citric acid and Cr resistant microbes improved castor bean growth and photosynthesis while it alleviated Cr toxicity by reducing Cr+6 to Cr3+. Microorganisms 2021, 9, 2499. [Google Scholar] [CrossRef]
- Nemat, H.; Shah, A.A.; Akram, W.; Ramzan, M.; Yasin, N.A. Ameliorative effect of co-application of Bradyrhizobium japonicum EI09 and Se to mitigate chromium stress in Capsicum annum L. Int. J. Phytoremediat. 2020, 22, 1396–1407. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Chouhan, R.; Bakshi, P.; Gandhi, S.G.; Kaur, R.; Sharma, A.; Bhardwaj, R. Amelioration of chromium-Induced oxidative stress by combined treatment of selected plant-growth-promoting rhizobacteria and earthworms via modulating the expression of genes related to reactive oxygen species metabolism in Brassica juncea. Front. Microbiol. 2022, 13, 802512. [Google Scholar] [CrossRef]
- Alabdallah, N.M.; Al-Shammari, A.S.; Saleem, K.; Alzahrani, S.S.; Raza, A.; Asghar, M.A.; Ullah, A.; Hussain, M.I.; Yong, J.W.H. Unveiling the mechanisms of silicon-induced salinity stress tolerance in Panicum turgidum: Insights from antioxidant defense system and comprehensive metabolic and nutritional profiling. S. Afr. J. Bot. 2024, 168, 328–339. [Google Scholar] [CrossRef]
- Rizvi, A.; Khan, M.S. Heavy metal-induced oxidative damage and root morphology alterations of maize (Zea mays L.) plants and stress mitigation by metal tolerant nitrogen-fixing Azotobacter chroococcum. Ecotoxicol. Environ. Saf. 2018, 157, 9–20. [Google Scholar] [CrossRef]
- Tirry, N.; Joutey, N.T.; Sayel, H.; Kouchou, A.; Bahafid, W.; Asri, M.; El Ghachtouli, N. Screening of plant growth promoting traits in heavy metals resistant bacteria: Prospects in phytoremediation. J. Genet. Eng. Biotechnol. 2018, 16, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Mishra, J.; Arora, N.K. Plant growth promoting bacteria for combating salinity stress in plants—Recent developments and prospects: A review. Microbiol. Res. 2021, 252, 126861. [Google Scholar] [CrossRef] [PubMed]
- Sarwar, M.J.; Shabaan, M.; Asghar, H.N.; Ayyub, M.; Ali, Q.; Zulfiqar, U.; Nazim, M.; Alarjani, K.M.; Elshikh, M.S. Interaction of chromium (Cr) resistant plant growth promoting rhizobacteria with compost to phytostabilize Cr in spinach rhizosphere. Plant Stress 2023, 10, 100261. [Google Scholar] [CrossRef]
- Islam, M.K.; Alam, I.; Khanam, M.S.; Lee, S.Y.; Waghmode, T.R.; Huh, M.R. Accumulation and tolerance characteristics of chromium in nine jute varieties (“Corchorus spp.” and “Hibiscus spp.”). Plant Omics 2014, 7, 392–402. [Google Scholar]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobahan, M.A.; Akter, N.; Karim, M.M.; Badhon, M.M.I.; Khan, S.N.; Alam, S.; Prasad, P.V.V.; Hasanuzzaman, M. Bioaugmentation with Plant Growth-Promoting Rhizobacteria Alleviates Chromium and Salt Stress in Rice Through the Improvement of Physiology, Ion Homeostasis, and Antioxidant Defense. Microorganisms 2025, 13, 1462. https://doi.org/10.3390/microorganisms13071462
Sobahan MA, Akter N, Karim MM, Badhon MMI, Khan SN, Alam S, Prasad PVV, Hasanuzzaman M. Bioaugmentation with Plant Growth-Promoting Rhizobacteria Alleviates Chromium and Salt Stress in Rice Through the Improvement of Physiology, Ion Homeostasis, and Antioxidant Defense. Microorganisms. 2025; 13(7):1462. https://doi.org/10.3390/microorganisms13071462
Chicago/Turabian StyleSobahan, Muhammad Abdus, Nasima Akter, Muhammad Manjurul Karim, Md. Muzahidul Islam Badhon, Shakila Nargis Khan, Samiul Alam, P.V. Vara Prasad, and Mirza Hasanuzzaman. 2025. "Bioaugmentation with Plant Growth-Promoting Rhizobacteria Alleviates Chromium and Salt Stress in Rice Through the Improvement of Physiology, Ion Homeostasis, and Antioxidant Defense" Microorganisms 13, no. 7: 1462. https://doi.org/10.3390/microorganisms13071462
APA StyleSobahan, M. A., Akter, N., Karim, M. M., Badhon, M. M. I., Khan, S. N., Alam, S., Prasad, P. V. V., & Hasanuzzaman, M. (2025). Bioaugmentation with Plant Growth-Promoting Rhizobacteria Alleviates Chromium and Salt Stress in Rice Through the Improvement of Physiology, Ion Homeostasis, and Antioxidant Defense. Microorganisms, 13(7), 1462. https://doi.org/10.3390/microorganisms13071462