

Salt-Tolerant Bacteria Support Salinity Stress Mitigating Impact of Arbuscular Mycorrhizal Fungi in Maize (Zea mays L.)



Abstract
1. Introduction
2. Materials and Methods
2.1. Soil Sampling and Isolation of Salt-Tolerant Bacteria
2.2. Estimating Plant Growth Promotion Traits of Salt-Tolerant Bacteria
2.3. Identification of Bacteria
2.4. Experimental Setup
2.5. Bacterial Count Determination and Estimation of Mycorrhizal Colonization
2.6. Growth Measurements and Photosynthetic Pigments Determination
2.7. Determination of Nutrients, Proline, and Enzyme Activities
2.8. Statistical Analysis
3. Results
3.1. Isolation and Screening of Salt-Tolerant Bacteria
3.2. Plant Growth Promotion Traits of Salt-Tolerant Bacteria
3.3. Identification of the Most Effective Salt-Tolerant Bacterial Isolate
3.4. Bacterial Count and Mycorrhizal Colonization Under Salinity Stress
3.5. Morphological Traits of Maize Plants Under Salinity Stress
3.6. Photosynthetic-Related Pigments of Maize Plants Under Salinity Stress
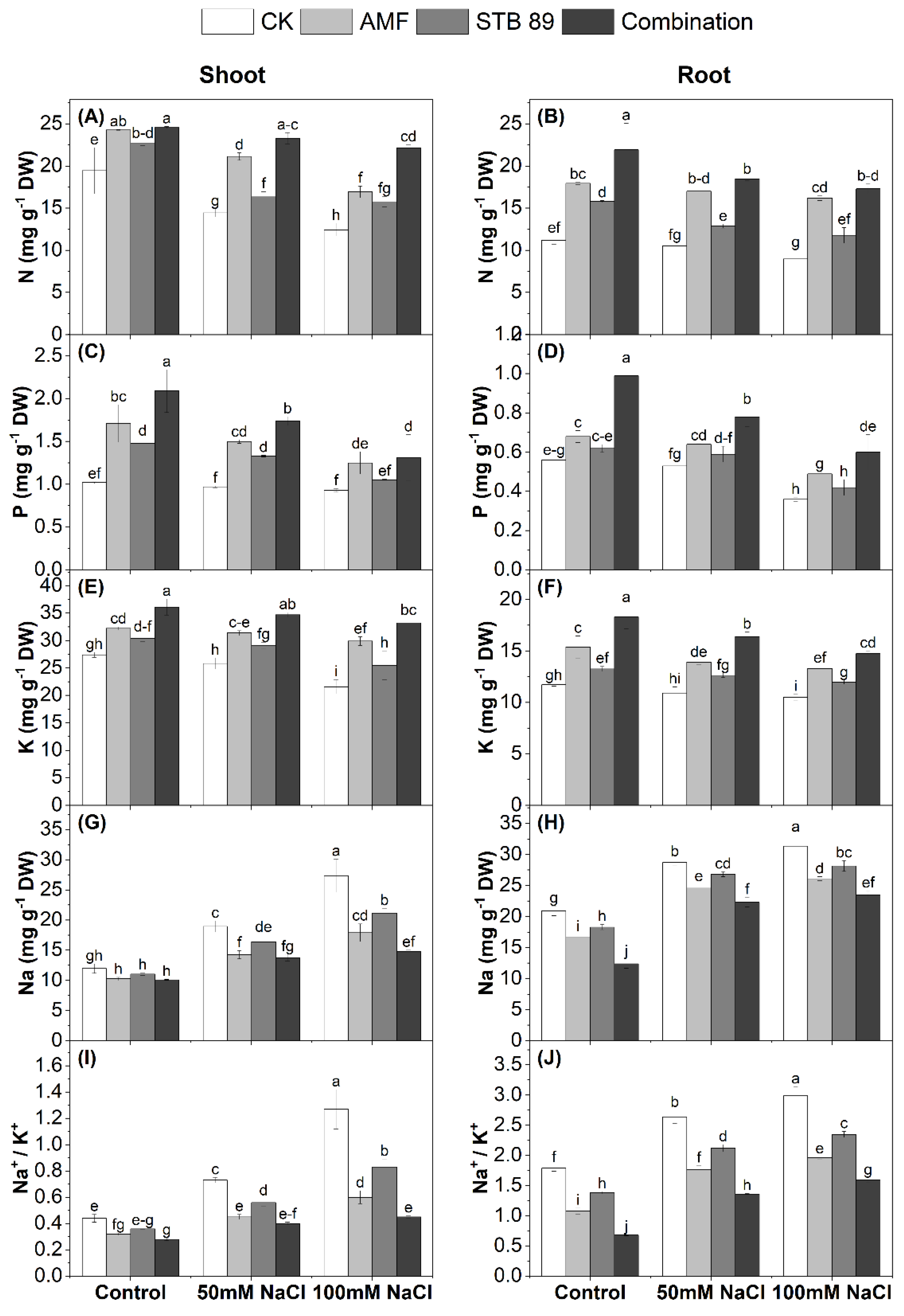
3.7. Nutrient Content and Determination of the Na+/K+ Ratio
3.8. Proline Content and Determination of Antioxidant Enzyme Activities
3.9. Evaluation of the Impact of AMF, STB, and Their Combination by Principal Component Analysis and Heatmap of Correlation Under Salinity Stress
4. Discussion
4.1. Isolated Salt-Tolerant Bacteria Produce Plant Growth Substances Actively
4.2. AMF and STB 89 Improve Bacterial Counts and Mycorrhizal Colonization
4.3. AMF and STB 89 Improve the Morphological Traits of Maize Plants
4.4. AMF and STB 89 Improve Photosynthetic-Related Pigments Content in Maize Plants
4.5. AMF and STB 89 Enhance Nutrient Content and Reduce the Na+/K+ Ratio in Maize Plants
4.6. AMF and STB 89 Reduce Proline Content and Activate Antioxidant Defense System in Maize Plants
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Negm, A.M.; Omran, E.-S.E.; Mahmoud, M.A.; Abdel-Fattah, S. Update, Conclusions, and Recommendations for Conventional Water Resources and Agriculture in Egypt. In Conventional Water Resources and Agriculture in Egypt; Negm, A.M., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 659–681. [Google Scholar]
- El-Sawah, A.M.; Abdel-Fattah, G.G.; Holford, P.; Korany, S.M.; Alsherif, E.A.; AbdElgawad, H.; Ulhassan, Z.; Jośko, I.; Ali, B.; Sheteiwy, M.S. Funneliformis constrictum modulates polyamine metabolism to enhance tolerance of Zea mays L. to salinity. Microbiol. Res. 2023, 266, 127254. [Google Scholar] [CrossRef]
- Sang, H.; Guo, W.; Gao, Y.; Jiao, X.; Pan, X. Effects of Alternating Fresh and Saline Water Irrigation on Soil Salinity and Chlorophyll Fluorescence of Summer Maize. Water 2020, 12, 3054. [Google Scholar] [CrossRef]
- Liu, C.; Jiang, X.; Yuan, Z. Plant Responses and Adaptations to Salt Stress: A Review. Horticulturae 2024, 10, 1221. [Google Scholar] [CrossRef]
- Boorboori, M.R.; Lackóová, L. Arbuscular mycorrhizal fungi and salinity stress mitigation in plants. Front. Plant Sci. 2025, 15, 1504970. [Google Scholar] [CrossRef]
- Muhammad, M.; Waheed, A.; Wahab, A.; Majeed, M.; Nazim, M.; Liu, Y.-H.; Li, L.; Li, W.-J. Soil salinity and drought tolerance: An evaluation of plant growth, productivity, microbial diversity, and amelioration strategies. Plant Stress 2024, 11, 100319. [Google Scholar] [CrossRef]
- Evelin, H.; Devi, T.S.; Gupta, S.; Kapoor, R. Mitigation of Salinity Stress in Plants by Arbuscular Mycorrhizal Symbiosis: Current Understanding and New Challenges. Front. Plant Sci. 2019, 10, 470. [Google Scholar] [CrossRef] [PubMed]
- Balasubramaniam, T.; Shen, G.; Esmaeili, N.; Zhang, H. Plants’ Response Mechanisms to Salinity Stress. Plants 2023, 12, 2253. [Google Scholar] [CrossRef] [PubMed]
- Atta, K.; Mondal, S.; Gorai, S.; Singh, A.P.; Kumari, A.; Ghosh, T.; Roy, A.; Hembram, S.; Gaikwad, D.J.; Mondal, S.; et al. Impacts of salinity stress on crop plants: Improving salt tolerance through genetic and molecular dissection. Front. Plant Sci. 2023, 14, 1241736. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, Z.; Feng, S.; Guo, P.; Wang, Y.; Hao, B.; Guo, W.; Li, F.Y. Synergistic effects of AMF and PGPR on improving saline-alkaline tolerance of Leymus chinensis by strengthening the link between rhizosphere metabolites and microbiomes. Environ. Technol. Innov. 2024, 36, 103900. [Google Scholar] [CrossRef]
- Wen, Y.; Wu, R.; Qi, D.; Xu, T.; Chang, W.; Li, K.; Fang, X.; Song, F. The effect of AMF combined with biochar on plant growth and soil quality under saline-alkali stress: Insights from microbial community analysis. Ecotoxicol. Environ. Saf. 2024, 281, 116592. [Google Scholar] [CrossRef]
- Khalloufi, M.; Martínez-Andújar, C.; Karray-Bouraouib, N.; Pérez-Alfocea, F.; Albacete, A. The crosstalk interaction of ethylene, gibberellins, and arbuscular mycorrhiza improves growth in salinized tomato plants by modulating the hormonal balance. J. Plant Physiol. 2024, 303, 154336. [Google Scholar] [CrossRef]
- Huang, P.; Huang, S.; Ma, Y.; Danish, S.; Hareem, M.; Syed, A.; Elgorban, A.M.; Eswaramoorthy, R.; Wong, L.S. Alleviation of salinity stress by EDTA chelated-biochar and arbuscular mycorrhizal fungi on maize via modulation of antioxidants activity and biochemical attributes. BMC Plant Biol. 2024, 24, 63. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Zhang, P.; Cao, F.; Liu, X.; Ji, M.; Xie, M. Effects of AMF on Maize Yield and Soil Microbial Community in Sandy and Saline Soils. Plants 2024, 13, 2056. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho Neta, S.J.; Araújo, V.L.V.P.; Fracetto, F.J.C.; da Silva, C.C.G.; de Souza, E.R.; Silva, W.R.; Lumini, E.; Fracetto, G.G.M. Growth-promoting bacteria and arbuscular mycorrhizal fungus enhance maize tolerance to saline stress. Microbiol. Res. 2024, 284, 127708. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; An, T.; Huang, D.; Liu, R.; Xu, B.; Zhang, S.; Deng, X.; Siddique, K.H.M.; Chen, Y. Arbuscular mycorrhizal symbioses alleviating salt stress in maize is associated with a decline in root-to-leaf gradient of Na+/K+ ratio. BMC Plant Biol. 2021, 21, 457. [Google Scholar] [CrossRef]
- Chandra, P.; Singh, A.; Prajapat, K.; Rai, A.K.; Yadav, R.K. Native arbuscular mycorrhizal fungi improve growth, biomass yield, and phosphorus nutrition of sorghum in saline and sodic soils of the semi–arid region. Environ. Exp. Bot. 2022, 201, 104982. [Google Scholar] [CrossRef]
- Gao, C.; El-Sawah, A.M.; Ali, D.F.I.; Alhaj Hamoud, Y.; Shaghaleh, H.; Sheteiwy, M.S. The Integration of Bio and Organic Fertilizers Improve Plant Growth, Grain Yield, Quality and Metabolism of Hybrid Maize (Zea mays L.). Agronomy 2020, 10, 319. [Google Scholar] [CrossRef]
- Xu, W.; Liu, Q.; Wang, B.; Zhang, N.; Qiu, R.; Yuan, Y.; Yang, M.; Wang, F.; Mei, L.; Cui, G. Arbuscular mycorrhizal fungi communities and promoting the growth of alfalfa in saline ecosystems of northern China. Front. Plant Sci. 2024, 15, 1438771. [Google Scholar] [CrossRef]
- Malik, J.A.; Alqarawi, A.A.; Alotaibi, F.; Habib, M.M.; Sorrori, S.N.; Almutairi, M.B.R.; Dar, B.A. Alleviation of NaCl Stress on Growth and Biochemical Traits of Cenchrus ciliaris L. via Arbuscular Mycorrhizal Fungi Symbiosis. Life 2024, 14, 1276. [Google Scholar] [CrossRef]
- Kakabouki, I.; Stavropoulos, P.; Roussis, I.; Mavroeidis, A.; Bilalis, D. Contribution of Arbuscular Mycorrhizal Fungi (AMF) in Improving the Growth and Yield Performances of Flax (Linum usitatissimum L.) to Salinity Stress. Agronomy 2023, 13, 2416. [Google Scholar] [CrossRef]
- Qin, W.; Yan, H.; Zou, B.; Guo, R.; Ci, D.; Tang, Z.; Zou, X.; Zhang, X.; Yu, X.; Wang, Y.; et al. Arbuscular mycorrhizal fungi alleviate salinity stress in peanut: Evidence from pot-grown and field experiments. Food Energy Secur. 2021, 10, e314. [Google Scholar] [CrossRef]
- Ghosh, S.; Bhowmik, S.; Dutta, S.S. Challenges in Application of Arbuscular Mycorrhizal Inocula in Conventional Agriculture. In Arbuscular Mycorrhizal Fungi in Sustainable Agriculture: Inoculum Production and Application; Parihar, M., Rakshit, A., Adholeya, A., Chen, Y., Eds.; Springer Nature: Singapore, 2024; pp. 229–252. [Google Scholar]
- Szymańska, S.; Lis, M.I.; Piernik, A.; Hrynkiewicz, K. Pseudomonas stutzeri and Kushneria marisflavi Alleviate Salinity Stress-Associated Damages in Barley, Lettuce, and Sunflower. Front. Microbiol. 2022, 13, 788893. [Google Scholar] [CrossRef]
- Lami, M.J.; Adler, C.; Caram-Di Santo, M.C.; Zenoff, A.M.; de Cristóbal, R.E.; Espinosa-Urgel, M.; Vincent, P.A. Pseudomonas stutzeri MJL19, a rhizosphere-colonizing bacterium that promotes plant growth under saline stress. J. Appl. Microbiol. 2020, 129, 1321–1336. [Google Scholar] [CrossRef]
- Szymańska, S.; Dąbrowska, G.B.; Tyburski, J.; Niedojadło, K.; Piernik, A.; Hrynkiewicz, K. Boosting the Brassica napus L. tolerance to salinity by the halotolerant strain Pseudomonas stutzeri ISE12. Environ. Exp. Bot. 2019, 163, 55–68. [Google Scholar] [CrossRef]
- Ke, X.; Feng, S.; Wang, J.; Lu, W.; Zhang, W.; Chen, M.; Lin, M. Effect of inoculation with nitrogen-fixing bacterium Pseudomonas stutzeri A1501 on maize plant growth and the microbiome indigenous to the rhizosphere. Syst. Appl. Microbiol. 2019, 42, 248–260. [Google Scholar] [CrossRef] [PubMed]
- Bacilio, M.; Moreno, M.; Bashan, Y. Mitigation of negative effects of progressive soil salinity gradients by application of humic acids and inoculation with Pseudomonas stutzeri in a salt-tolerant and a salt-susceptible pepper. Appl. Soil Ecol. 2016, 107, 394–404. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.; Bellingrath-Kimura, S.D.; Mishra, J.; Arora, N.K. Salt-Tolerant Plant Growth Promoting Rhizobacteria for Enhancing Crop Productivity of Saline Soils. Front. Microbiol. 2019, 10, 2791. [Google Scholar] [CrossRef]
- Patel, T.; Saraf, M. Biosynthesis of phytohormones from novel rhizobacterial isolates and their in vitro plant growth-promoting efficacy. J. Plant Interact. 2017, 12, 480–487. [Google Scholar] [CrossRef]
- Sadeghi, A.; Karimi, E.; Dahaji, P.A.; Javid, M.G.; Dalvand, Y.; Askari, H. Plant growth promoting activity of an auxin and siderophore producing isolate of Streptomyces under saline soil conditions. World J. Microbiol. Biotechnol. 2012, 28, 1503–1509. [Google Scholar] [CrossRef]
- Tiwari, G.; Duraivadivel, P.; Sharma, S.; Hariprasad, P. 1-Aminocyclopropane-1-carboxylic acid deaminase producing beneficial rhizobacteria ameliorate the biomass characters of Panicum maximum Jacq. by mitigating drought and salt stress. Sci. Rep. 2018, 8, 17513. [Google Scholar] [CrossRef]
- Qurashi, A.; Sabri, A. Bacterial exopolysaccharide and biofilm formation stimulate chickpea growth and soil aggregation under salt stress. Braz. J. Microbiol. 2012, 43, 11183–11191. [Google Scholar] [CrossRef]
- Saum, S.H.; Müller, V. Salinity-Dependent Switching of Osmolyte Strategies in a Moderately Halophilic Bacterium: Glutamate Induces Proline Biosynthesis in Halobacillus halophilus. J. Bacteriol. 2007, 189, 6968–6975. [Google Scholar] [CrossRef] [PubMed]
- Bharti, N.; Pandey, S.S.; Barnawal, D.; Patel, V.K.; Kalra, A. Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci. Rep. 2016, 6, 34768. [Google Scholar] [CrossRef] [PubMed]
- Kohler, J.; Hernández, J.A.; Caravaca, F.; Roldán, A. Induction of antioxidant enzymes is involved in the greater effectiveness of a PGPR versus AM fungi with respect to increasing the tolerance of lettuce to severe salt stress. Environ. Exp. Bot. 2009, 65, 245–252. [Google Scholar] [CrossRef]
- Patel, D.; Saraf, M. Influence of soil ameliorants and microflora on induction of antioxidant enzymes and growth promotion of Jatropha curcas L. under saline condition. Eur. J. Soil Biol. 2013, 55, 47–54. [Google Scholar] [CrossRef]
- Tu, Q.; Tang, S.; Huang, S. Mitigation of salinity stress via improving growth, chlorophyll contents and antioxidants defense in sunflower with Bacillus pumilis and biochar. Sci. Rep. 2025, 15, 9641. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, D.; Alheswairini, S.S.; Barasarathi, J.; Enshasy, H.A.E.; Lalitha, S.; Mir, S.H.; Nithyapriya, S.; Sayyed, R. Halophilic rhizobacteria promote growth, physiology and salinity tolerance in Sesamum indicum L. grown under salt stress. Front. Microbiol. 2025, 16, 1590854. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, P.; Wang, B.; Li, H.; Li, S.; Zhang, H.; Haider, F.U.; Li, X. Harnessing the role of rhizo-bacteria to mitigate salinity stress in rice (Orzya sativa); focus on antioxidant defense system, photosynthesis response, and rhizosphere microbial diversity. Rhizosphere 2025, 33, 101043. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H.M. Salt stress in maize: Effects, resistance mechanisms, and management. A review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef]
- Jackson, M.L. Soil Chemical Analysis: Advanced Course: A Manual of Methods Useful for Instruction and Research in Soil Chemistry, Physical Chemistry of Soils, Soil Fertility, and Soil Genesis; UW-Madison Libraries parallel press: Madison, WI, USA, 2005. [Google Scholar]
- Nader, A.A.; Hauka, F.I.A.; Afify, A.H.; El-Sawah, A.M. Drought-Tolerant Bacteria and Arbuscular Mycorrhizal Fungi Mitigate the Detrimental Effects of Drought Stress Induced by Withholding Irrigation at Critical Growth Stages of Soybean (Glycine max, L.). Microorganisms 2024, 12, 1123. [Google Scholar] [CrossRef]
- Abdelsattar, M.; El-Sawah, A.M.; El-Kady, S.; Hauka, F.I.A. Evaluation of Plant Growth Promoting of Salt-tolerant Rhizobacteria Isolated from Egyptian Saline Soils. J. Agric. Chem. Biotechnol. 2022, 13, 95–100. [Google Scholar] [CrossRef]
- Ahmad, F.; Ahmad, I.; KHAN, M.S. Indole acetic acid production by the indigenous isolates of Azotobacter and fluorescent Pseudomonas in the presence and absence of tryptophan. Turk. J. Biol. 2005, 29, 29–34. [Google Scholar]
- Abou-Aly, H.E.; Youssef, A.M.; El-Meihy, R.M.; Tawfik, T.A.; El-Akshar, E.A. Evaluation of heavy metals tolerant bacterial strains as antioxidant agents and plant growth promoters. Biocatal. Agric. Biotechnol. 2019, 19, 101110. [Google Scholar] [CrossRef]
- Pikovskaya, R. Mobilization of phosphorus in soil in connection with vital activity of some microbial species. Mikrobiologiya 1948, 17, 362–370. [Google Scholar]
- Boltz, D.F.; Mellon, M.G. Spectrophotometric Determination of Phosphorus as Molybdiphosphoric Acid. Anal. Chem. 1948, 20, 749–751. [Google Scholar] [CrossRef]
- Hemalatha, N.; Raja, N.; Jayachitra, A.; Rajalakshmi, A.; Valarmathi, N. Isolation and characterization of phosphate solubilizing bacteria and analyzing their effect on Capsicum annum L. Inter. J. Biol. Pharm. Res 2013, 4, 159–167. [Google Scholar]
- Sharath, S.; Triveni, S.; Nagaraju, Y.; Latha, P.C.; Vidyasagar, B. The Role of Phyllosphere Bacteria in Improving Cotton Growth and Yield Under Drought Conditions. Front. Agron. 2021, 3, 680466. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Skerman, V.B. A Guide to the Identification of the Genera of Bacteria, 2nd ed.; The Williams & Wilkins Co.: Baltimore, MD, USA, 1967. [Google Scholar]
- Setiawati, T.C.; Mutmainnah, L. Solubilization of potassium containing mineral by microorganisms from sugarcane rhizosphere. Agric. Agric. Sci. Procedia 2016, 9, 108–117. [Google Scholar] [CrossRef]
- Phillips, J.; Hayman, D. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158-IN118. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Measure du taux de mycorrhization d’un systeme radiculaire. Recherche de methods d’estimation ayant une signification fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae; INRA: Paris, France, 1986; pp. 217–221. [Google Scholar]
- Amanullah, H.; Marwat, K.; Shah, P.; Maula, N.; Arifullah, S. Nitrogen levels and its time of application influence leaf area, height and biomass of maize planted at low and high density. Pak. J. Bot 2009, 41, 761–768. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Snell, F.D.; Snell, C.T. Colorimetric Methods of Analysis; D. van Nostrand Company, Inc.: New York, NY, USA, 1961. [Google Scholar]
- Chapman, H.D.; Pratt, P.F. Methods of analysis for soils, plants and waters. Soil Sci. 1962, 93, 68. [Google Scholar] [CrossRef]
- Malik, C.P.; Singh, M.B. Plant Enzymology and Histo Enzymology; Kalyani Publishers: New Delhi, India, 1980; p. 286. [Google Scholar]
- Hammerschmidt, R.; Nuckles, E.M.; Kuć, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Aebi, H.E. Catalase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Verlag Chemie: Weinhem, Germany, 1983; pp. 273–286. [Google Scholar]
- Alonazi, M.A.; Alwathnani, H.A.; AL-Barakah, F.N.I.; Alotaibi, F. Native Plant Growth-Promoting Rhizobacteria Containing ACC Deaminase Promote Plant Growth and Alleviate Salinity and Heat Stress in Maize (Zea mays L.) Plants in Saudi Arabia. Plants 2025, 14, 1107. [Google Scholar] [CrossRef] [PubMed]
- Afify, A.; Ashour, A. Use of Cyanobacteria for Controlling Flax Seedling Blight. J. Agric. Chem. Biotechnol. 2018, 9, 259–261. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Reyad, A.M.; Khalaf, M.H.; Sheteiwy, M.S.; Dawood, M.F.A.; El-Sawah, A.M.; Shaban Ahmed, E.; Malik, A.; Al-Qahtani, W.H.; Abdel-Maksoud, M.A.; et al. Investigating the Endophyte Actinomycetota sp. JW0824 Strain as a Potential Bioinoculant to Enhance the Yield, Nutritive Value, and Chemical Composition of Different Cultivars of Anise (Pimpinella anisum L.) Seeds. Biology 2024, 13, 553. [Google Scholar] [CrossRef] [PubMed]
- El-Amriti, F.A.; Ouf, S.A.; Abu-Elghait, M.; Desouky, S.E.; Mohamed, M.S.M. Alleviation of salt stress on Zea mays L. plant by PGPR isolates as an effective sustainable strategy. Biocatal. Agric. Biotechnol. 2024, 61, 103346. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Sheteiwy, M.S.; El-Keblawy, A.; Ulhassan, Z.; Khalaf, M.H.; Mohamed, H.S.; Okla, M.K.; AlGarawi, A.M.; El-Sawah, A.M.; Ahmed, E.S.; et al. The potential biofortification role of Actinopolyspora sp. JTT-01 in enhancing the yield and tissue chemical composition of caraway plants. BMC Plant Biol. 2025, 25, 540. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Mukherjee, A.; Rastogi, R.P.; Verma, J.P. Salt-tolerant plant growth-promoting Bacillus pumilus strain JPVS11 to enhance plant growth attributes of rice and improve soil health under salinity stress. Microbiol. Res. 2021, 242, 126616. [Google Scholar] [CrossRef]
- Shahid, M.; Altaf, M.; Ali, S.; Tyagi, A. Isolation and assessment of the beneficial effect of exopolysaccharide-producing PGPR in Triticum aestivum (L.) plants grown under NaCl and Cd-stressed conditions. Plant Physiol. Biochem. 2024, 215, 108973. [Google Scholar] [CrossRef] [PubMed]
- Juniper, S.; Abbott, L.K. Soil salinity delays germination and limits growth of hyphae from propagules of arbuscular mycorrhizal fungi. Mycorrhiza 2006, 16, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Huang, C.; Peng, F.; Wang, T.; Liao, J.; Ma, S.; You, Q.; Xue, X. Synergistic combination of arbuscular mycorrhizal fungi and plant growth-promoting rhizobacteria modulates morpho-physiological characteristics and soil structure in Nitraria tangutorum bobr. Under saline soil conditions. Res. Cold Arid Reg. 2022, 14, 393–402. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Boopathi, T.; Manivannan, P. Comprehensive Assessment of Ameliorative Effects of AMF in Alleviating Abiotic Stress in Tomato Plants. J. Fungi 2021, 7, 303. [Google Scholar] [CrossRef] [PubMed]
- Echeverria, M.; Sannazzaro, A.I.; Ruiz, O.A.; Menéndez, A.B. Modulatory effects of Mesorhizobium tianshanense and Glomus intraradices on plant proline and polyamine levels during early plant response of Lotus tenuis to salinity. Plant Soil 2013, 364, 69–79. [Google Scholar] [CrossRef]
- Elhindi, K.M.; El-Din, A.S.; Elgorban, A.M. The impact of arbuscular mycorrhizal fungi in mitigating salt-induced adverse effects in sweet basil (Ocimum basilicum L.). Saudi J. Biol. Sci. 2017, 24, 170–179. [Google Scholar] [CrossRef]
- Borde, M.; Dudhane, M.; Jite, P. Growth photosynthetic activity and antioxidant responses of mycorrhizal and non-mycorrhizal bajra (Pennisetum glaucum) crop under salinity stress condition. Crop Prot. 2011, 30, 265–271. [Google Scholar] [CrossRef]
- Ouhaddou, R.; Meddich, A.; Ikan, C.; Lahlali, R.; Ait Barka, E.; Hajirezaei, M.-R.; Duponnois, R.; Baslam, M. Enhancing Maize Productivity and Soil Health under Salt Stress through Physiological Adaptation and Metabolic Regulation Using Indigenous Biostimulants. Plants 2023, 12, 3703. [Google Scholar] [CrossRef]
- Shabaan, M.; Asghar, H.N.; Zahir, Z.A.; Zhang, X.; Sardar, M.F.; Li, H. Salt-Tolerant PGPR Confer Salt Tolerance to Maize Through Enhanced Soil Biological Health, Enzymatic Activities, Nutrient Uptake and Antioxidant Defense. Front. Microbiol. 2022, 13, 901865. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Sample No. | Location | Soil Type | EC Values (dSm−1) | pH Values | Isolates Number | Isolates Key |
|---|---|---|---|---|---|---|
| S1 | Klabsho | Sandy | 10.25 | 8.59 | 12 | STB: 14,27,32,77,78,79,80, 81,82, 83, 84,85, 87 |
| S2 | Klabsho | Sandy | 2.63 | 8.55 | 15 | STB: 24,25,26,28,29,30,31,33, 34,35,36,37,38,39,40 |
| S3 | Klabsho | Sandy | 3.38 | 8.52 | 3 | STB: 1,2,4 |
| S4 | North Delta | Clay | 2.38 | 8.34 | 3 | STB: 74,75,76 |
| S5 | Bahariya Oasis | Sandy | 9.90 | 8.11 | 1 | STB: 87 |
| S6 | Bahariya Oasis | Sandy | 10.71 | 8.24 | 5 | STB: 86,88,89,90,95 |
| S7 | Bahariya Oasis | Sandy | 7.19 | 7.98 | 3 | STB: 91,92,96 |
| S8 | Bahariya Oasis | Sandy | 9.35 | 8.04 | 3 | STB: 93,94,97 |
| S9 | El-Senbellawein | Clay | 7.36 | 8.79 | 21 | STB: 17,41,42,43,44,58,59, 60,61,62,63,64,65,66, 67,68,69,70,71,72,73 |
| S10 | Gamasa | Sandy | 2.11 | 7.98 | 31 | STB: 3,5,6,7,8,9,10,11,12, 13,15,16,18,19,20,21, 22,23,45,46,47,48,49, 50,51,52,53,54,55,56,57 |
| Property | Value |
|---|---|
| Particle size distribution (%) | |
| Coarse Sand | 2.24 |
| Fine Sand | 23.51 |
| Silt | 42.00 |
| Clay | 32.26 |
| Soil texture | Clay loam |
| Physical and chemical analysis | |
| OC % | 0.42 |
| OM % | 0.73 |
| pH | 8.05 |
| EC | 1.64 |
| Cations (meq L−1) | |
| Ca++ | 6.11 |
| Mg++ | 2.97 |
| Na+ | 9.24 |
| K+ | 0.43 |
| Anions (meq L−1) | |
| CO3−2 | 0.00 |
| HCO3− | 1.30 |
| Cl− | 5.79 |
| SO4−2 | 11.65 |
| Available nutrients (mg kg−1) | |
| N | 67.56 |
| P | 19.66 |
| K | 343 |
| Bacterial count (Log (cfu g−1 dry soil)) | |
| TBC | 6.238 |
| PSBC | 5.113 |
| KRBC | 4.369 |
| Treatments | Count of Bacteria Log (cfu g−1 Dry Soil) | Levels of AMF Colonization (%) | |||||
|---|---|---|---|---|---|---|---|
| TCB | PSBC | KRBC | F | M | A | ||
| Control | CK | 7.876 ± 0.027e | 5.825 ± 0.016ef | 5.106 ± 0.005e | – | – | – |
| AMF | 7.972 ± 0.016d | 5.974 ± 0.002c | 5.154 ± 0.013d | 90.91 ± 0.36ab | 67.92 ± 1.87b | 57.73 ± 0.78b | |
| STB 89 | 8.128 ± 0.043c | 6.108 ± 0.048b | 5.218 ± 0.006c | – | – | – | |
| Combination | 8.267 ± 0.021a | 6.259 ± 0.012a | 5.288 ± 0.011a | 91.67 ± 1.64a | 72.27 ± 0.54a | 63.37 ± 1.40a | |
| 50 mM NaCl | CK | 7.535 ± 0.026h | 5.771 ± 0.048f | 5.037 ± 0.033g | – | – | – |
| AMF | 7.800 ± 0.038f | 5.876 ± 0.012de | 5.091 ± 0.004ef | 81.82 ± 1.19c | 61.00 ± 1.11d | 40.95 ± 0.85d | |
| STB 89 | 7.980 ± 0.009d | 5.936 ± 0.007cd | 5.200 ± 0.006c | – | – | – | |
| Combination | 8.196 ± 0.008b | 6.110 ± 0.051b | 5.247 ± 0.007b | 90.00 ± 1.06b | 62.50 ± 0.56c | 42.80 ± 0.53c | |
| 100 mM NaCl | CK | 7.314 ± 0.023i | 5.624 ± 0.056g | 5.027 ± 0.009g | – | – | – |
| AMF | 7.715 ± 0.021g | 5.795 ± 0.043f | 5.074 ± 0.008f | 60.00 ± 0.45e | 31.82 ± 1.33f | 24.55 ± 0.63f | |
| STB 89 | 7.895 ± 0.020e | 5.902 ± 0.016d | 5.164 ± 0.015d | – | – | – | |
| Combination | 8.098 ± 0.008c | 5.965 ± 0.014c | 5.221 ± 0.016c | 72.73 ± 2.18d | 35.50 ± 0.61e | 27.82 ± 0.88e | |
| Salinity | *** | *** | *** | *** | *** | *** | |
| Bio-inoculation | *** | *** | *** | *** | *** | *** | |
| Salinity × Bio-inoculation | *** | ** | ns | *** | *** | *** | |
| Treatments | Shoot Length (cm) | Root Length (cm) | SDW (g Plant −1) | RDW (g Plant −1) | Leaf Area (cm2) | |
|---|---|---|---|---|---|---|
| Control | CK | 98.00 ± 5.98bc | 16.97 ± 2.8ef | 11.81 ± 0.68e | 1.51 ± 0.46de | 237.99 ± 18.5fg |
| AMF | 120.33 ± 4.04a | 21.53 ± 0.4b | 19.02 ± 3.93b | 4.43 ± 0.17b | 320.95 ± 5.8bc | |
| STB 89 | 107.93 ± 3.27b | 20.87 ± 0.35b | 17.08 ± 0.96bc | 3.05 ± 0.61c | 297.57 ± 16.76c–e | |
| Combination | 125.7 ± 2.48a | 25.43 ± 1.1a | 24.61 ± 0.81a | 5.41 ± 0.07a | 366.57 ± 20.36a | |
| 50 mM NaCl | CK | 80.53 ± 1.82de | 14.27 ± 1.1g | 8.38 ± 1.32fg | 0.90 ± 0.08ef | 196.98 ± 10.47h |
| AMF | 99.6 ± 13.43bc | 18.93 ± 0.23cd | 14.69 ± 0.56d | 2.68 ± 0.64c | 307.94 ± 4.08b–d | |
| STB 89 | 89.83 ± 9.17cd | 17.07 ± 0.12ef | 10.85 ± 0.79e | 1.61 ± 0.31d | 267.99 ± 6.41ef | |
| Combination | 105.97 ± 3.41b | 21.63 ± 0.78b | 15.26 ± 1.03cd | 4.64 ± 0.48b | 334.88 ± 3.37b | |
| 100 mM NaCl | CK | 61.57 ± 4.67f | 12.13 ± 0.12h | 4.50 ± 0.36h | 0.52 ± 0.01f | 130.88 ± 9.93i |
| AMF | 73.00 ± 1.30e | 18.17 ± 0.47de | 6.49 ± 0.08gh | 0.94 ± 0.04ef | 211.26 ± 8.02gh | |
| STB 89 | 72.77 ± 0.65e | 15.97 ± 1.05fg | 4.85 ± 0.14h | 0.70 ± 0.05f | 182.27 ± 12.32h | |
| Combination | 81.43 ± 5.02de | 20.07 ± 0.6bc | 9.69 ± 0.09ef | 1.42 ± 0.41de | 286.57 ± 48.9de | |
| Salinity | *** | *** | *** | *** | *** | |
| Bio-inoculation | *** | *** | *** | *** | *** | |
| Salinity × Bio-inoculation | ns | ns | *** | *** | ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaki, R.M.; Afify, A.H.; Ashour, E.H.; El-Sawah, A.M. Salt-Tolerant Bacteria Support Salinity Stress Mitigating Impact of Arbuscular Mycorrhizal Fungi in Maize (Zea mays L.). Microorganisms 2025, 13, 1345. https://doi.org/10.3390/microorganisms13061345
Zaki RM, Afify AH, Ashour EH, El-Sawah AM. Salt-Tolerant Bacteria Support Salinity Stress Mitigating Impact of Arbuscular Mycorrhizal Fungi in Maize (Zea mays L.). Microorganisms. 2025; 13(6):1345. https://doi.org/10.3390/microorganisms13061345
Chicago/Turabian StyleZaki, Randa M., Aida H. Afify, Eman H. Ashour, and Ahmed M. El-Sawah. 2025. "Salt-Tolerant Bacteria Support Salinity Stress Mitigating Impact of Arbuscular Mycorrhizal Fungi in Maize (Zea mays L.)" Microorganisms 13, no. 6: 1345. https://doi.org/10.3390/microorganisms13061345
APA StyleZaki, R. M., Afify, A. H., Ashour, E. H., & El-Sawah, A. M. (2025). Salt-Tolerant Bacteria Support Salinity Stress Mitigating Impact of Arbuscular Mycorrhizal Fungi in Maize (Zea mays L.). Microorganisms, 13(6), 1345. https://doi.org/10.3390/microorganisms13061345