Abstract
Root-knot nematodes (RKNs; Meloidogyne incognita) pose a significant threat to tomato crops, necessitating sustainable control methods. This study investigated the inoculation efficacy of co-cultured Burkholderia vietnamiensis B418 and Trichoderma harzianum T11W compared with single-strain treatments for RKNs suppression and their influence on the structure and function of the rhizosphere microbiome. Co-inoculation with B418 + T11W achieved a 71.42% reduction in the disease index, significantly outperforming single inoculations of B418 (54.46%) and T11W (58.93%). Co-inoculation also increased plant height by 38.51% and fresh weight by 76.02% compared to the RKNs infested plants control, promoting robust tomato growth. Metagenomic analysis reveals that co-inoculation enhanced bacterial diversity, with 378 unique bacterial species and a high Shannon index, while fungal diversity decreased with Trichoderma dominance (83.31% abundance). Actinomycetota (46.42%) and Ascomycota (97.92%) were enriched in the co-inoculated rhizosphere, showing negative correlations with RKNs severity. Functional analysis indicates enriched metabolic pathways, including streptomycin and unsaturated fatty acid biosynthesis, enhancing microbial antagonism. Single inoculations altered pathways like steroid degradation (B418) and terpenoid biosynthesis (T11W), but co-inoculation uniquely optimized the rhizosphere microenvironment. These findings highlight co-inoculation with B418 + T11W effectively suppressing RKNs and fostering plant health by reshaping microbial communities and functions, offering a promising approach for sustainable agriculture.
1. Introduction
Root-knot nematodes (RKNs; Meloidogyne incognita) are widespread plant parasites that severely impact economically important crops [1]. Their strong parasitism and broad distribution make them a major threat to agriculture [2]. Traditionally, chemical control, along with agricultural practices and resistant varieties, has been used, but it has limitations regarding environmental and human health issues. With advances in science and growing environmental concern, biological control has gained significant attention [3]. The use of antagonistic plant growth-promoting rhizobacteria (PGPR), such as Bacillus, Burkholderia, Pseudomonas, and Pasteuria, has been reported to effectively control RKNs while promoting plant growth and yield [4,5]. Kim et al. found that the cell-free filtrate of Burkholderia sp. JB-2 exhibited strong nematicidal activity, achieving an 87.5% mortality rate against second-instar larvae (J2) within 2 days [6]. Liu et al. reported that the individual application of B. vietnamiensis B418 presented a high control efficiency of 71.15% against RKNs on watermelon in a field experiment, and the control efficiencies remained at 62.71% and 67.87% when combined with fosthiazate and avermectin, exhibiting slight incompatibility, which was still notably higher than when the nematicides were applied separately [7].
Microbial co-culture systems exhibit high-efficiency potentials in suppressing RKNs and plant pathogens by activating complex interspecies interactions [8]. Studies show that the synergistic effects of multiple beneficial microorganisms significantly enhance nematicidal efficacy. For example, after co-culturing Bacillus amyloliquefaciens Sneb709 and Sinorhizobium fredii Sneb183, the inhibition rate of the fermentation broth against J2 of M. incognita significantly increased to 84.17%, far exceeding the effect of single strains [9]. Siddiqui et al. demonstrated that culture filtrate extracts of Trichoderma harzianum Th6 and Pseudomonas fluorescens CHA0 enhanced nematicidal activity and reduced RKNs population density in tomato plants [10]. The application of microbial agents or stimulants has been reported to directly inhibit RKNs, trigger plant resistance, and enhance the biodiversity of antagonistic microbial communities [11]. The effect of co-culture depends on the specific strains involved and their relative proportions. Compared with mono-cultures, co-culturing not only influences the microbial growth but also has a greater effect on metabolism. There was no previous report regarding Trichoderma and Burkholderia co-culture for RKNs inhibition.
Microbiomes play a crucial role in soil and agro-ecosystems [12]. Soil metagenomics enables the exploration of diverse microbial communities without the need for cultivation [13,14]. Whole-genome sequencing has been widely used to study soil samples, uncovering the functional diversity of microbial metagenomes [15]. Rhizosphere microbiota support tomato plant growth and development, enhancing its overall health [16,17]. Studies show that co-cultures significantly increased microbial diversity and biomass in the rhizosphere while also isolating rare and novel bacterial species [18,19]. Li et al. found that RKNs parasitism significantly influenced the diversity and assembly of plant-associated microbial communities [20]. Similarly, Lu et al. reported a strong correlation between rhizosphere bacterial distribution in tobacco and RKNs infection, identifying 15 genera positively correlated and 42 genera negatively correlated with RKNs density [21]. Notably, some negatively correlated genera, such as Pseudomonas and Bacillus, are known to help suppress soil-borne pathogens. The combined use of two biocontrol strains significantly altered the composition of the soil bacterial community and influenced the relative abundance of beneficial microorganisms in the rhizosphere [22]. Kamalanathan et al. found that treatment with Bacillus velezensis VB7 and T. koningiopsis TK modified the fungal community structure in the rhizosphere, promoted the accumulation of diverse nematode-resistant biomolecules, induced defense mechanisms that inhibited nematode infection, and enhanced plant growth [23].
Previous studies have demonstrated that both Burkholderia vietnamiensis B418 and Trichoderma harzianum T11W are effective against RKNs [7,24,25]. This study aims to evaluate the effectiveness of a co-cultured fermentation broth of these two strains in controlling tomato RKNs compared to single-train treatments and analyze differences in the rhizosphere microbiome structure and function under single and co-inoculation conditions.
2. Materials and Methods
2.1. Strains and Media
The biocontrol strains B. vietnamiensis B418 (China General Microbiological Culture Collection Center, CGMCC No. 1212) and T. harzianum T11W (CGMCC No. 7938) were isolated and preserved by the Environmental Microbiology Laboratory, Ecology Institute of Shandong Academy of Sciences [7,24,25]. Both strains were preserved at −80 °C prior to use. Meloidogyne incognita was obtained from the roots of diseased tomato plants collected from vegetable-growing greenhouses, as previously described [7,24]. The tomato variety used in this study was Sweet Treasure, a nematode susceptible cultivar harvested from greenhouses in Zhangqiu District, Jinan City, Shandong Province.
Various culture media were prepared for microbial growth and experimentation. A tryptone yeast extract (TY) medium was prepared with peptone (10 g), yeast powder (1 g), CaCl2 (0.2 g), and distilled water (1 L), adjusted to a pH of 7.2–7.4. The potato dextrose agar (PDA) consisted of peeled potato (200 g), dextrose (20 g), and agar (15 g) dissolved in 1 L of distilled water. The potato dextrose broth (PDB) was similarly composed of peeled potato (200 g) and dextrose (20 g) in 1 L of distilled water. Additionally, Modified King’s B broth (MKB) contained casein amino acids (20 g L−1), glycerol (10 mL), K2HPO4 (1.5 g L−1), MgSO4·7H2O (1.5 g L−1), and distilled water (1 L), adjusted to pH 7.2 ± 0.2.
2.2. Co-Culture Conditions
For the preparation of B. vietnamiensis B418, bacterial stocks preserved at −80 °C were retrieved and inoculated onto TY plates. After single colonies developed, an inoculating loop was used to transfer the colonies into a TY liquid medium. The culture was incubated at 28 °C with shaking at 180 rpm for 12 h. The B418 bacterial suspension was diluted 106 folds and spread onto TY plates, and then the individual colonies were counted to make the diluted suspension of 1 × 108 CFU/mL for further experiments.
For T. harzianum T11W, fungal plugs stored in glycerol at −80 °C were inoculated onto PDA plates and incubated at 28 °C for 5 days. After incubation, spores were washed from the plates, and their concentration was determined using a hemocytometer. The spore suspension was then diluted to 1 × 108 CFU/mL for use in co-culture experiments.
To establish the B. vietnamiensis B418 and T. harzianum T11W co-culture, 200 μL of B418 bacterial and T11W fungal spore suspensions were inoculated into 200 mL of MKB at a ratio of 1:1 in 1000 mL conical flasks. For the mono-culture, T11W and B418 were inoculated into 200 mL of MKB at the same inoculation ratio and cultivated in the same conditions, respectively. The cultures were incubated at 28 °C with shaking at 180 rpm for 14 days, resulting in the production of a mono- and co-culture fermentation broth.
2.3. Preparation of Root-Knot Nematodes (RKNs)
Tomato roots infected with RKNs were cut into small pieces and washed in the laboratory. The root fragments were soaked in a 0.5% NaClO solution for 5 min, then rinsed thoroughly by passing the suspension through 50 μm and 100 μm sieves under running water. The suspension was centrifuged at 3000 rpm for 3 min, after which the supernatant was discarded. An equal volume of 75% sucrose was added to the remaining suspension, followed by another centrifugation at 3000 rpm for 3 min to remove impurities.
The nematode suspension was further disinfected with a 0.1% NaClO solution for 3 min and then rinsed with sterile water under aseptic conditions. The nematode inoculum level was determined using an inverted microscope and adjusted to 200 eggs/mL, as previously described by Zuhair et al. [26].
2.4. Pot Experiment
Three-week-old tomato plants with uniform growth were transplanted into pots filled with RKNs-free soil, which was pre-tested by sucrose density centrifugation and microscopy examination to confirm the absence of nematodes. The RKNs-free soil has a pH value of 7.0, the organic matter content is 3%, and the soil texture is loam. After the tomato plants had been allowed to recover from transplantation for 7 days, select plants with uniform plant height for the experiment. The experimental design is summarized in Table 1, including four treatments (designated as RKNs, B418, T11W, and B418 + T11W). For each treatment, 20 mL of nematode suspension (200 eggs/mL) were infested into the pot, while 10 mL of 14-day fermentation broth of mono and co-culture were inoculated to tomato plants for B418, T11W, and B418 + T11W treatments, respectively. There were 16 replicates planted in 4 pots for each treatment, and they were randomly placed during the experiment. The mean temperature in the greenhouse was set to 28–35 °C during the day and 10–15 °C during the night, with 80–90% relative humidity. After 60 days of growth, plant height, fresh weight, and disease severity were measured at harvest. The disease index of the root knots (gall index) was assessed using the six-grade classification method [27] and calculated according to the standard formula [28]:

Table 1.
Experimental designs for pot experiment.
2.5. Metagenomic Sequencing and Bioinmatics
Three tomato plants out of four pots were randomly selected from each treatment. The bulk soil around the roots was shaken off, and any soil tightly attached to the root segment within the range of 0–4 mm was gently brushed off. The collected rhizosphere soil samples were stored at −80 °C for further analysis. DNA was extracted from soil samples using a Power Soil DNA Isolation Kit (Qiagen, Hilden, Germany). One microliter of RNase A was added to the tube to digest RNA, followed by incubation at 37 °C for 15 min. The concentration and purity of the extracted DNA were assessed using a Nanodrop spectrophotometer and agarose gel electrophoresis. Sequencing libraries were prepared using an NEBNext® UltraTM DNA Library Prep Kit for Illumina (NEB, Ipswich, MA, USA), according to the manufacturer’s instructions, targeting a fragment size of 350 bp. The libraries were then sequenced on an Illumina NovaSeq platform, generating paired-end reads.
The raw sequencing data was preprocessed to improve quality by removing sequencing adapters and filtering out reads containing low-quality bases or sequences shorter than 50 bp using Kneaddata (Version 0.7.4) software [29]. Additionally, Bowtie2 (Version 2.3.5.1) software [30] was used to eliminate host plant genomic DNA by filtering out reads of the host origin before further analysis. Finally, FastQC (Version 0.11.9) was employed to assess the effectiveness and reliability of the quality control process. Kraken2 (Version 2.0.7-beta) [31], along with a microbial database containing sequences from bacteria, fungi, archaea, and viruses (screened from the NT nucleic acid database and the RefSeq (https://www.ncbi.nlm.nih.gov/refseq/, accessed on 8 November 2023) whole genome database of NCBI (https://www.ncbi.nlm.nih.gov/); accessed on 8 November 2023, was used to identify the species present in the samples. Bracken (Version 2.0) [32] was then employed to estimate the relative abundance of these species. Kraken2, a k-mer-based classification tool, utilized a local database comprising 16,799 known bacterial genomes. Following quality control and host sequence removal, clean reads were aligned to the UniRef90 database (http://www.uniprot.org/uniref/; accessed on 8 November 2023) using HUMAnN3 (Version 3.6) software [33], which operates with DIAMOND (Version 0.8.22) for sequence alignment. Based on the correspondence between UniRef90 IDs and the KEGG database (https://www.kegg.jp/; accessed on 8 November 2023), functional annotation data and relative abundance tables were generated.
2.6. Statistical Analysis
The data were statistically analyzed using SPSS 26.0 (SPSS, Chicago, IL, USA) and PRISM 8.3.0.538 software (GraphPad Software, San Diego, CA, USA). Analysis of variance (ANOVA) was performed following Tukey’s honest significant difference (HSD) test to identify significant differences among the samples (p < 0.05). LEfSe (Linear Discriminant Analysis Effect Size) was used to identify rhizosphere microbiome functions that showed significant differences between treatments.
3. Results
Tomato plants subjected to the RKNs treatment without microbial inoculation exhibited dwarfing and wilting symptoms 60 days after the addition of the nematode suspension (Figure 1). In contrast, plants treated with B418 + T11W and T11W alone grew taller than those treated with B418 alone, with average heights of 23.31–24.28 cm compared to 22.14 cm (Table 2). The B418 + T11W treatment resulted in a 38.51% increase in plant height relative to the RKNs control, while T11W and B418 treatments led to increases of 32.97% and 26.30%, respectively. There was no significant difference in fresh weight between the B418 + T11W and T11W treatments, both of which yielded the highest fresh weights among the treatments. The B418 + T11W treatment increased fresh weight by 76.02% compared to the RKNs control, while T11W and B418 increased it by 65.78% and 39.34%, respectively. Specifically, the combined treatment of B418 + T11W significantly reduced the disease index of tomato plants to 26.67 ± 1.56 compared with the individual treatments of B418 and T11W, which were 38.33 ± 1.56 and 42.50 ± 1.02, respectively (p < 0.05) (Table 2). The co-inoculation with B418 + T11W achieved a 71.42% reduction in the disease index compared to the RKNs control, significantly outperforming single inoculations of B418 (54.46%) and T11W (58.93%).

Figure 1.
Results of tomato pot experiment in greenhouse conditions. (A) Potted plant images of different treatments; (B) root development of different treatments. Control: RKNs, B418: B. vietnamiensis B418 + RKNs, T11W: T. harzianum T11W + RKNs, B418 + T11W: B. vietnamiensis B418 + T. harzianum T11W + RKNs, RKNs: root-knot nematodes.
Table 2.
The plant height, fresh weight, disease index, and control effect of tomato in the different treatments 60 days after planting.
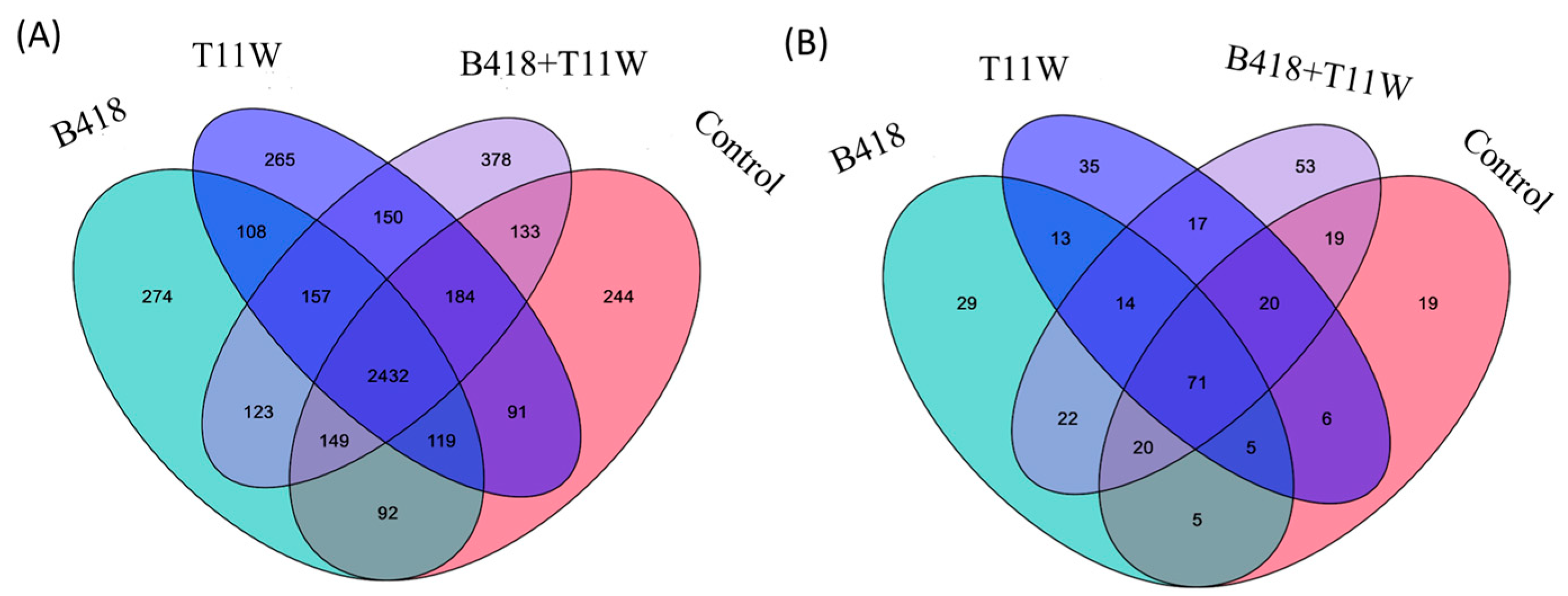
A total of 2432 bacterial species were detected in the rhizosphere soil across all four treatments (Figure 2A). The majority of these species were common to all treatments. Notably, 378 species were uniquely found in the rhizosphere of plants inoculated with the B418 + T11W combination, which was higher than the number of unique species in the other treatments. A similar pattern was observed for fungi, where most species were shared across treatments, but the co-culture group also harbored the highest number of unique fungal species (Figure 2B).

Figure 2.
(A) Comparison of OTUs at bacterial species level in different treatments. (B) Comparison of OTUs at fungal species level in different treatments. Control = RKNs, B418 = B. vietnamiensis B418 + RKNs, T11W = T. harzianum T11W + RKNs, B418 + T11W = B. vietnamiensis B418 + T. harzianum T11W + RKNs.
The rhizosphere bacterial community in the co-culture B418 + T11W treatment exhibited the highest Shannon diversity index, followed by the B418 treatment (Figure 3A). In contrast, the non-inoculated control and T11W treatments showed lower Shannon indices compared to the co-culture treatment. Conversely, for the fungal community, the Shannon index was lowest in the B418 + T11W treatment, while the control and B418 treatments displayed the highest fungal diversity (Figure 3B).
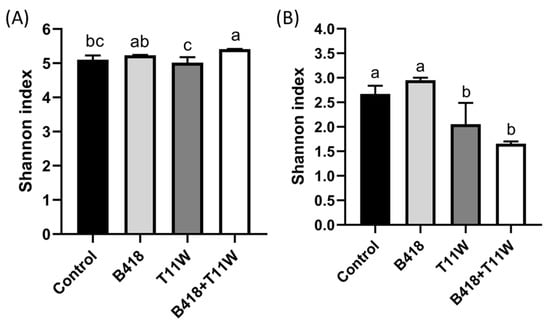
Figure 3.
Shannon diversity index of (A) bacterial and (B) fungal communities in the tomato rhizosphere under different inoculation treatments. Control: RKNs; B418: B. vietnamiensis B418 + RKNs; T11W: T. harzianum T11W + RKNs; B418 + T11W: B. vietnamiensis B418 + T. harzianum T11W + RKNs. RKNs: root-knot nematodes. Error bar indicates standard error. The same letter indicates no significant difference based on Duncan’s test.
The microbial community composition in the rhizosphere soil varied among the four treatments (Figure 4). The influence of different inoculation treatments on the bacterial community resembled their effect on the fungal community. Specifically, the microbial community structure in the rhizosphere of plants inoculated with the B418 + T11W combination differed from that observed in plants treated with either strain individually.
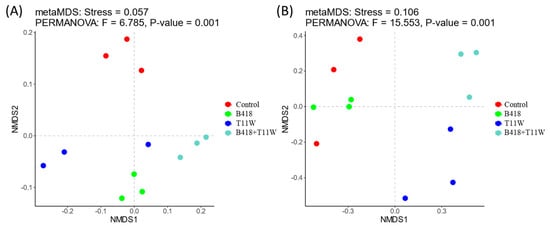
Figure 4.
Non-metric multidimensional scaling based on the Bray–Curtis distance for the composition of the (A) bacterial species and (B) fungal species in the tomato rhizosphere under different inoculation treatments. Control: RKNs; B418: B. vietnamiensis B418 + RKNs; T11W: T. harzianum T11W + RKNs; B418 + T11W: B. vietnamiensis B418 + T. harzianum T11W + RKNs. RKNs: root-knot nematodes. (A) PERMANOVA’s pseudo-F = 6.785, p-value = 0.001; (B) PERMANOVA’s pseudo-F = 15.553, p-value = 0.001.
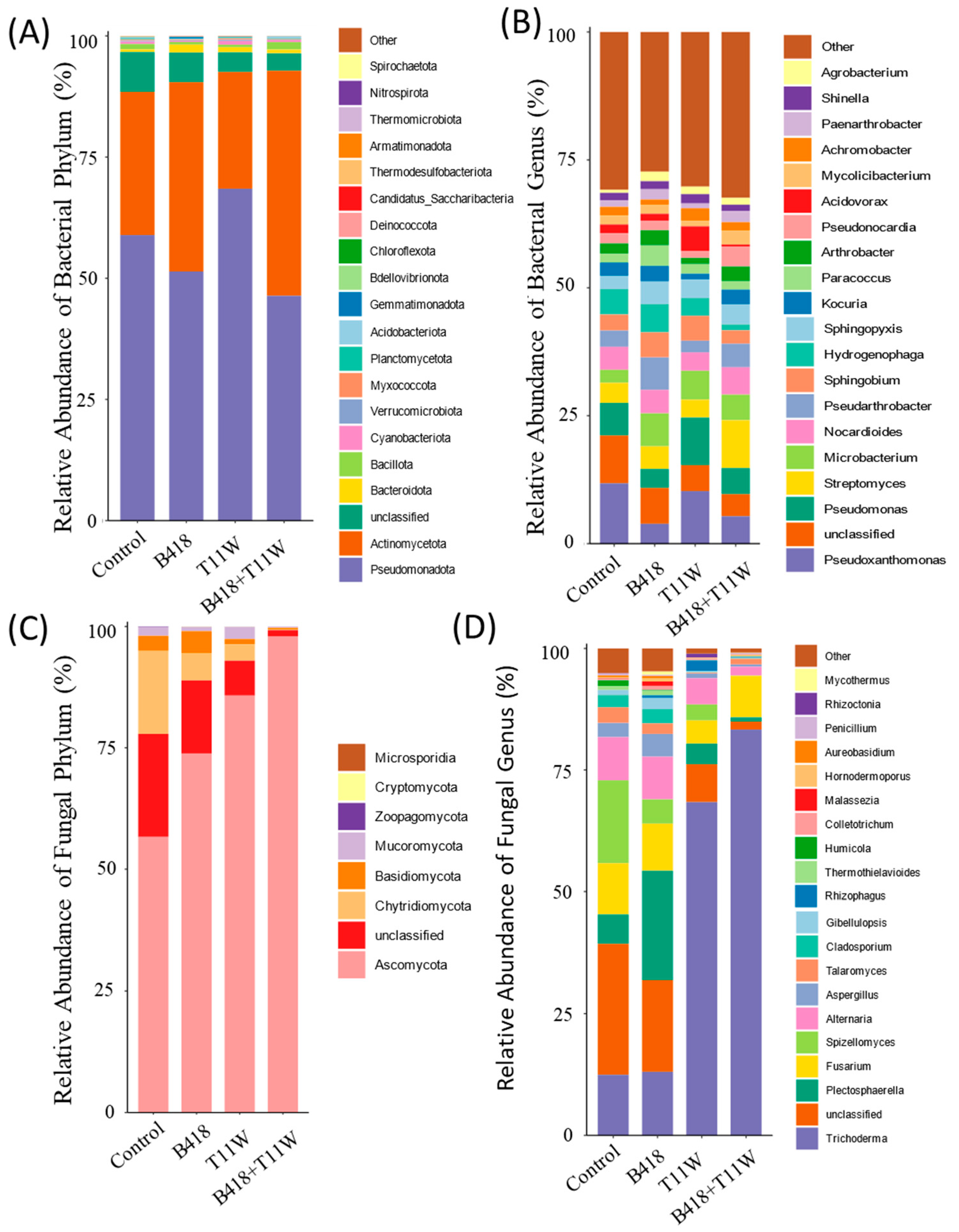
At the phylum level, Pseudomonadota and Actinomycetota were the dominant bacterial groups across all four treatments. In the control treatment, their relative abundances were 58.91% and 29.53%, respectively. In the B418-treated rhizosphere soil, Pseudomonadota accounted for 51.4% and Actinomycetota for 39.05%. For the T11W treatment, Pseudomonadota made up 68.48%, while Actinomycetota represented 24.09%. In the rhizosphere of plants treated with the B418 + T11W co-culture, Actinomycetota showed the highest relative abundance at 46.42%, closely followed by Pseudomonadota at 46.38% (Figure 5A). The predominance of Actinomycetota and Pseudomonadota in the rhizosphere under B418 + T11W, B418, and T11W treatments may be a key factor contributing to the suppression of RKNs.

Figure 5.
Relative abundances of microbial communities under different inoculation treatments. (A) Bacterial phylum; (B) bacterial genus; (C) fungal phylum; (D) fungal genus. Control: RKNs; B418: B. vietnamiensis B418 + RKNs; T11W: T. harzianum T11W + RKNs; B418 + T11W: B. vietnamiensis B418 + T. harzianum T11W + RKNs. RKNs: root-knot nematodes.
The bacterial genera Pseudoxanthomonas and Pseudomonas were dominant across all rhizosphere soil samples. Pseudoxanthomonas had the highest relative abundance in the control group, followed by the T11W treatment, while its abundance decreased in the B418 + T11W co-culture treatment. In contrast, Streptomyces reached its highest relative abundance (9.35%) in the B418 + T11W treatment compared to 4.42% in the B418 group, 3.52% in T11W, and 3.94% in the control group (Figure 5B).
In terms of the fungal community, Ascomycota was the most dominant phylum across all treatments. Its relative abundance was 56.63% in the non-inoculated control, increasing to 73.83% in the B418 group and 85.78% in the T11W group. The highest abundance was observed in the B418 + T11W co-culture treatment, reaching 97.92%, indicating that Ascomycota may play a key role in suppressing RKNs and enhancing plant growth (Figure 5C).
At the genus level, Trichoderma was dominant in all treatments. The co-culture B418 + T11W treatment showed a substantial increase in Trichoderma abundance compared to the control, rising from 12.44% to 83.31%. In the B418 and T11W treatments, Trichoderma accounted for 13.05% and 68.38%, respectively. The B418 + T11W treatment also markedly reduced the abundance of unclassified fungi from 26.89% in the control to just 1.65%. Plectosphaerella was most abundant in the B418 group (22.38%), followed by the control (6.03%) and T11W (4.24%), with the lowest abundance in the B418 + T11W group (0.86%) (Figure 5D).
The marked increase in Trichoderma levels in both the T11W and B418 + T11W treatments indicates that the inoculation significantly boosted its presence (Figure 6). The relative abundance of T. harzianum mirrored that of the Trichoderma genus, with the highest level found in the B418 + T11W group (48.89%), followed by the T11W group (38.63%). No significant difference in T. harzianum abundance was observed between the T11W and B418 + T11W treatments or between the control and B418 treatments.

Figure 6.
The proportions of Trichoderma harzianum under different inoculation treatments. Control: RKNs; B418: B. vietnamiensis B418 + RKNs; T11W: T. harzianum T11W + RKNs; B418 + T11W: B. vietnamiensis B418 + T. harzianum T11W + RKNs. RKNs: root-knot nematodes. Error bar indicates standard error. The same letter indicates no significant difference based on Duncan’s test.
Functional pathways among different treatments were presented in Figure 7 using linear discriminant analysis effect size (LEfSe), with linear discriminant analysis (LDA) scores greater than 2.5 and a significance threshold of p < 0.05. In the B418 treatment group, four metabolic pathways showed significantly different abundances compared to the other treatments. These pathways included steroid degradation, peptidoglycan biosynthesis, central carbon metabolism in cancer, and protein export, corresponding to the KEGG functional categories of metabolism, human diseases, genetic information processing, and cellular processes, respectively. Among these, the key genes involved in the central carbon metabolism in the cancer pathway were G6PD and zwf, both encoding glucose-6-phosphate 1-dehydrogenase.

Figure 7.
Functional pathways that showed significant differences from other treatments were identified using linear discriminant analysis effect size (LEfSe), with linear discriminant analysis (LDA) scores greater than 2.5 and a significance threshold of p < 0.05. Control: RKNs; B418: B. vietnamiensis B418 + RKNs; T11W: T. harzianum T11W + RKNs; B418 + T11W: B. vietnamiensis B418 + T. harzianum T11W + RKNs. RKNs: root-knot nematodes.
In the T11W treatment group, significantly enriched metabolic pathways were categorized into four major functional classes. The first was cellular processes, represented by quorum sensing and bacterial chemotaxis. Key genes identified in bacterial chemotaxis included mcp and fliM, encoding the methyl-accepting chemotaxis protein and the flagellar motor switch protein FliM, respectively. The second category, environmental information processing, included the ABC transporter system and bacterial secretion system. The key genes within the ABC transporters were phnE, livK, and urtE, which encode proteins associated with phosphonate transport, branched-chain amino acid transport, and urea transport, respectively. The third category, human diseases, involved pathways related to chemical carcinogenesis−DNA adducts and platinum drug resistance− both associated with the genes GST and gst, which encode glutathione S-transferase. The fourth category, metabolism, comprised pathways including glutathione metabolism, drug metabolism (cytochrome P450), xenobiotic metabolism (cytochrome P450), lipopolysaccharide biosynthesis, and biosynthesis of terpenoids and steroids. In these pathways, GST and gst again appeared as key genes involved in glutathione S-transferase activity.
In the B418 + T11W co-inoculation group, all significantly enriched pathways were related to metabolic functions. These included amino sugar and nucleotide sugar metabolism, ethylbenzene degradation, starch and sucrose metabolism, lipoarabinomannan (LAM) biosynthesis, galactose metabolism, styrene degradation, biosynthesis of unsaturated fatty acids, and streptomycin biosynthesis. The gene glk, encoding glucokinase, was identified as a key gene in both amino sugar and nucleotide sugar metabolism, as well as in starch and sucrose metabolism. Additionally, bglB (encoding beta-glucosidase) was involved in starch and sucrose metabolism. For ethylbenzene degradation, the key genes fadA and fadI encoding acetyl-CoA acyltransferase were identified. In galactose metabolism, the genes glk, pfkA, and PFK (encoding glucokinase and 6-phosphofructokinase) were prominent. Finally, for streptomycin biosynthesis, the key genes GCK and INO1, encoding glucokinase and myo-inositol-1-phosphate synthase, respectively, were significantly enriched.
4. Discussion
These findings demonstrate that co-inoculation with B418 and T11W provided the most effective control of root-knot nematodes (RKNs) in tomato plants, achieving a control efficacy of 71.43%. Individually, T. harzianum T11W and B. vietnamiensis B418 also showed moderate suppression of RKNs, with control rates ranging from 54% to 58%. However, the combined inoculation of both strains resulted in a significantly enhanced control effect. This co-inoculation strategy not only suppressed nematode infestation more effectively but also promoted tomato plant growth. Similarly, Sharma et al. reported that co-inoculation with Rhizophagus irregularis and Pseudomonas enhanced tomato growth and reduced nematode disease compared to single-strain treatments [34], although the inoculated beneficial microbes species differs from the present study. Notably, the present study further elucidates the mechanisms behind the enhanced performance of co-inoculation, revealing that it modulates both the composition and function of the rhizosphere microbiome.
Significant shifts in both bacterial and fungal community compositions were observed in the rhizosphere soil of tomato plants subjected to different inoculation treatments, with potential associations identified between these microbial changes and the incidence of RKNs disease. At the phylum level in the bacterial community, the relative abundance of Actinomycetota notably increased following microbial inoculation, with the most pronounced rise observed in the B418 + T11W co-inoculation treatment. This increase may play a crucial role in enhancing the suppression of tomato RKNs, as previous research has highlighted the nematode-inhibitory properties of Actinomycetota [35,36]. Moreover, actinomycetes are commonly utilized in agriculture as biocontrol agents and growth-promoting microbes for vegetable crops [37,38]. In the fungal community, the abundance of Ascomycota showed a negative correlation with RKNs disease severity in tomato. This observation aligns with the findings by Kamalanathan et al., who reported the highest levels of Ascomycota in the rhizosphere of tomato plants treated with biocontrol agents [23].
A significant increase in the proportion of Trichoderma spp. was observed in the rhizosphere microbial community of tomato plants under the T11W and B418 + T11W treatments compared to the control and B418 treatments. This increase is likely due to the introduction of the T11W strain in both groups. The dominance and persistence of Trichoderma spp. in the rhizosphere following inoculation align with the findings from previous studies. For instance, Wu et al. reported that Trichoderma spp. had a significantly higher relative abundance in treatments with a compound Trichoderma agent (CTA) than in water controls or compound fertilizer treatments, and it represented the most dominant genus in the CTA group [39].
Zhang et al. [40] demonstrated that treatment with Trichoderma biofertilizer significantly altered the fungal community structure in grassland rhizosphere soil, notably increasing the abundance of Archaeorhizomyces and Trichoderma while reducing the presence of Ophiosphaerella. Similarly, Halifu et al. [41] reported that Trichoderma spp. was the dominant fungal genus in the rhizosphere of Pinus sylvestris var. mongolica seedlings following inoculation with T. harzianum E15 and T. virens ZT05. The persistent dominance of Trichoderma spp. in rhizosphere soils has been widely attributed to their rapid growth rate, strong vitality, ability to quickly colonize available space, and efficient nutrient uptake. In contrast, Burkholderia spp. was not identified as a dominant genus in the bacterial communities of the B418 and B418 + T11W treatments, likely due to the higher overall bacterial diversity present in the rhizosphere soil under these conditions.
In this study, the application of microbial inoculants led to an increase in soil bacterial diversity, with the co-inoculation of B418 + T11W resulting in the most pronounced enhancement. Similar findings were reported by Zheng et al., where the introduction of Pantoea jilinensis D25 significantly elevated bacterial diversity in the tomato rhizosphere, particularly among beneficial taxa [42]. Likewise, Jie et al. observed that inoculation with Rhizophagus intraradices enhanced both bacterial and fungal diversity in the rhizosphere soil of soybean [43]. Soil microbial diversity plays a crucial role in determining soil health and is essential for sustainable soil development [44,45]. A decline in microbial diversity can impair soil ecosystem functions and productivity, as the richness and composition of microbial communities are fundamental to ecological processes [46]. High microbial diversity is also associated with improved plant protection against soil-borne pathogens [47]. The observed shift in microbial community composition following biological inoculation may, therefore, contribute to the suppression of root-knot nematode disease. On the other hand, the diversity of fungal communities in the rhizosphere decreased following inoculation with T11W or the B418 + T11W co-culture. This reduction is likely due to the dominance of Trichoderma introduced by these treatments, which may have outcompeted and limited the presence of other fungal species. There were usually plant pathogens combined with RKNs infestation due to the host vulnerability. The dominance of Trichoderma spp. in rhizosphere soils could, therefore, not only inhibit RKNs infestation but also prevent potential soil-borne diseases caused by secondary pathogens.
The LDA of functional pathway differences across treatment groups reveals that each inoculant exerted distinct influences on the functional dynamics of the tomato rhizosphere microbiome. In the B418 treatment group, rhizosphere microbes and enzymes associated with steroid degradation were particularly active, potentially contributing to the breakdown and transformation of steroid compounds in the soil environment [48]. Additionally, the upregulation of peptidoglycan biosynthesis in this group is likely linked to key bacterial physiological processes, particularly the formation of cell walls [49].
For the T11W treatment, notable enrichment was observed in pathways related to terpenoid and steroid biosynthesis, as well as bacterial chemotaxis. This suggests that the inoculation may stimulate the microbial production of terpenoids and steroids, compounds known to play roles in microbial ecology and physiological regulation [50]. Furthermore, the enhanced bacterial chemotaxis could improve microbial mobility, aiding in nutrient acquisition and the evasion of environmental stressors [51].
In the co-inoculated B418 + T11W treatment, there was a marked increase in metabolic activity related to the biosynthesis of streptomycin and unsaturated fatty acids. Streptomycin production can influence microbial antagonism and contribute to beneficial plant–microbe interactions, which are key for promoting plant health and improving soil fertility [52]. Meanwhile, the elevated synthesis of unsaturated fatty acids may impact microbial cell membrane fluidity and integrity, thereby influencing microbial metabolism and viability [53].
5. Conclusions
In summary, the use of biocontrol agents B. vietnamiensis B418, T. harzianum T11W, and their combination (B418 + T11W) effectively suppressed tomato RKNs, with the co-inoculation treatment showing the most pronounced control effect, reaching 71.42% reduction in the disease index. Additionally, these treatments significantly altered the composition and function of the rhizosphere microbial community. Application of the biocontrol agents notably enhanced soil bacterial diversity, with the most substantial increase observed in the B418 + T11W treatment. At the bacterial phylum level, Actinomycetota abundance increased markedly following biocontrol application and showed a strong negative correlation with RKNs incidence. For fungi, Ascomycota abundance also rose significantly under biocontrol treatments and was similarly negatively correlated with RKNs disease severity. Overall, the introduction of microbial inoculants reshaped the rhizosphere microbial structure, boosting the presence of beneficial microorganisms. The underlying mechanism involves the modulation of microbial metabolic pathways, which not only suppressed RKNs but also improved the soil microenvironment, thereby supporting healthier and more vigorous tomato plant growth.
Author Contributions
Y.J. and Y.W. (Yuanzheng Wu) conceived and designed the structure of this study. Y.J. and W.L. performed the experiments, collected data, provided interpretation, and prepared the original draft. J.L. and J.H. studied the data and obtained resources. Y.W. (Yanli Wei), Y.W. (Yilian Wang), and H.Y. performed data analysis with software. Y.Z. and S.Z. revised this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Shandong Provincial Natural Science Foundation (Grant number: ZR2021MC085), the Shandong Provincial Innovation Capability Promotion Project for Sci-tech Small and Medium-sized Enterprises (Grant number: 2022TSGC2370), and the Innovation Pilot Project of Integration of Science, Education, and Industry of Shandong Academy of Sciences (International Scientific and Technology Cooperation) (Grant number: 2024GH15).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding authors upon reasonable request.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Ralmi, N.H.A.A.; Khandaker, M.M.; Mat, N. Occurrence and control of root knot nematode in crops: A review. Aust. J. Crop Sci. 2016, 11, 1649–1654. [Google Scholar] [CrossRef]
- Sang, Y.H.; Ren, K.; Chen, Y.; Wang, B.; Meng, Y.F.; Zhou, W.B.; Jiang, Y.L.; Xu, J.J. Integration of soil microbiology and metabolomics to elucidate the mechanism of the accelerated infestation of tobacco by the root-knot nematode. Front. Microbiol. 2024, 15, 1455880. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.; Vicente, C.S.L.; Menéndez, E.; Faria, J.M.S.; Rusinque, L.; Camacho, M.J.; Inácio, M.L. The fight against plant-parasitic nematodes: Current status of bacterial and fungal biocontrol agents. Pathogens 2022, 11, 1178. [Google Scholar] [CrossRef] [PubMed]
- Aioub, A.A.A.; Elesawy, A.E.; Ammar, E.E. Plant growth promoting rhizobacteria (PGPR) and their role in plant-parasitic nematodes control: A fresh look at an old issue. J. Plant Dis. Prot. 2022, 129, 1305–1321. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, Y.; Chen, K.; Wu, Y.; Hu, J.; Wei, Y.; Li, J.; Yang, H.; Ryder, M.; Denton, M.D. Near-complete genomes of two Trichoderma species: A resource for biological control of plant pathogens. Mol. Plant-Microbe Interact. 2020, 33, 1036–1039. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, B.M.; Kang, M.K.; Park, D.J.; Choi, I.S.; Park, H.Y.; Lim, C.H.; Son, K.H. Assessment of nematicidal and plant growth-promoting effects of Burkholderia sp. JB-2 in root-knot nematode-infested soil. Front. Plant Sci. 2023, 14, 1216031. [Google Scholar] [CrossRef]
- Liu, M.M.; Philp, J.; Wang, Y.L.; Hu, J.D.; Wei, Y.L.; Li, J.S.; Ryder, M.; Toh, R.; Zhou, Y.; Denton, M.D.; et al. Plant growth-promoting rhizobacteria Burkholderia vietnamiensis B418 inhibits root-knot nematode on watermelon by modifying the rhizosphere microbial community. Sci. Rep. 2022, 12, 8381. [Google Scholar] [CrossRef]
- Xu, C.; Yu, H. Insights into constructing a stable and efficient microbial consortium. Chin. J. Chem. Eng. 2021, 30, 112–120. [Google Scholar] [CrossRef]
- Li, J.D.; Zhao, J.J.; Fan, H.Y.; Zhu, X.F.; Wang, Y.Y.; Liu, X.Y.; Duan, Y.X.; Chen, L.J. Optimization of fermentation conditions for co-culture nematicidal Bacillus amyloliquefaciens Sneb709 and Sinorhizobium fredii Sneb183. Chin. J. Biol. Control 2021, 37, 771–784. (In Chinese) [Google Scholar]
- Siddiqui, I.A.; Shaukat, S.S. Trichoderma harzianum enhances the production of nematicidal compounds in vitro and improves biocontrol of Meloidogyne javanica by Pseudomonas fluorescens in tomato. Lett. Appl. Microbiol. 2004, 38, 169–175. [Google Scholar] [CrossRef]
- Creamer, R.E.; Barel, J.M.; Bongiorno, G.; Zwetsloot, M. The life of soils: Integrating the who and how of multifunctionality. Soil Biol. Biochem. 2022, 166, 108561. [Google Scholar] [CrossRef]
- Jagadesh, M.; Dash, M.; Kumari, A.; Singh, S.K.; Verma, K.K.; Kumar, P.; Bhatt, R.; Sharma, S.K. Revealing the hidden world of soil microbes: Metagenomic insights into plant, bacteria, and fungi interactions for sustainable agriculture and ecosystem restoration. Microbiol. Res. 2024, 285, 127764. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Chowdhury, D.; Zhang, Z.M.; Cheung, W.K.; Lu, A.P.; Bian, Z.X.; Zhang, L. A review of computational tools for generating metagenome-assembled genomes from metagenomic sequencing data. Comput. Struct. Biotechnol. J. 2021, 19, 6301–6314. [Google Scholar] [CrossRef]
- Yue, H.; Yue, W.J.; Jiao, S.; Kim, H.; Lee, Y.; Wei, G.H.; Song, W.N.; Shu, D.T. Plant domestication shapes rhizosphere microbiome assembly and metabolic functions. Microbiome 2023, 11, 70. [Google Scholar] [CrossRef]
- Wani, A.K.; Rahayu, F.; Alkahtani, A.M.; Alreshidi, M.A.; Yadav, K.K.; Parnidi; Fauziah, L.; Murianingrum, M.; Akhtar, N.; Mufidah, E.; et al. Metagenomic profiling of rhizosphere microbiota: Unraveling the plant-soil dynamics. Physiol. Mol. Plant Pathol. 2024, 133, 102381. [Google Scholar] [CrossRef]
- Adedayo, A.A.; Fadiji, A.E.; Babalola, O.O. Plant health status affects the functional diversity of the rhizosphere microbiome associated with Solanum lycopersicum. Front. Sustain. Food Syst. 2022, 6, 894312. [Google Scholar] [CrossRef]
- Arora, D.; Gupta, P.; Jaglan, S.; Roullier, C.; Grovel, O.; Bertrand, S. Expanding the chemical diversity through microorganisms co-culture: Current status and outlook. Biotechnol. Adv. 2020, 40, 107521. [Google Scholar] [CrossRef]
- Hamid, B.; Zaman, M.; Farooq, S.; Fatima, S.; Sayyed, R.Z.; Baba, Z.A.; Sheikh, T.A.; Reddy, M.S.; Enshasy, H.E.; Gafur, A.; et al. Bacterial plant biostimulants: A sustainable way towards improving growth, productivity, and health of crops. Sustainability 2021, 13, 2856. [Google Scholar] [CrossRef]
- Berg, G.; Köberl, M.; Rybakova, D.; Müller, H.; Grosch, R.; Smalla, K. Plant microbial diversity is suggested as the key to future biocontrol and health trends. FEMS Microbiol. Ecol. 2017, 93, fix050. [Google Scholar] [CrossRef]
- Li, Y.; Lei, S.N.; Cheng, Z.Q.; Jin, L.Y.; Zhang, T.; Liang, L.M.; Cheng, L.J.; Zhang, Q.Y.; Xu, X.H.; Lan, C.H.; et al. Microbiota and functional analyses of nitrogen-fixing bacteria in root-knot nematode parasitism of plants. Microbiome 2023, 11, 48. [Google Scholar] [CrossRef]
- Lu, P.; Shi, H.L.; Tao, J.M.; Jin, J.J.; Wang, S.J.; Zheng, Q.X.; Liu, P.P.; Xiang, B.K.; Chen, Q.S.; Xu, Y.L.; et al. Metagenomic insights into the changes in the rhizosphere microbial community caused by the root-knot nematode Meloidogyne incognita in tobacco. Environ. Res. 2023, 216, 114848. [Google Scholar] [CrossRef]
- Yu, C.; Lv, J.; Xu, H.Y. Plant growth-promoting fungi and rhizobacteria control Fusarium damping-off in Mason pine seedlings by impacting rhizosphere microbes and altering plant physiological pathways. Plant Soil 2024, 499, 503–519. [Google Scholar] [CrossRef]
- Kamalanathan, V.; Sevugapperumal, N.; Nallusamy, S.; Ashraf, S.; Kailasam, K.; Afzal, M. Metagenomic approach deciphers the role of community composition of mycobiome structured by Bacillus velezensis VB7 and Trichoderma koningiopsis TK in tomato rhizosphere to suppress root-knot nematode infecting tomato. Microorganisms 2023, 11, 2467. [Google Scholar] [CrossRef]
- Wang, Y.L.; Li, J.S.; Yang, H.T. Identification of Trichoderma strain T11-W and its parasitism to southern root-knot nematode eggs. Shandong Sci. 2014, 27, 38–42. (In Chinese) [Google Scholar]
- Hu, J.D.; Zhou, Y.; Chen, K.; Li, J.S.; Wei, Y.L.; Wang, Y.L.; Wu, Y.Z.; Ryder, M.H.; Yang, H.T.; Denton, M.D. Large-scale Trichoderma diversity was associated with ecosystem, climate and geographic location. Environ. Microbiol. 2020, 22, 1011–1024. [Google Scholar] [CrossRef] [PubMed]
- Zuhair, R.; Moustafa, Y.T.A.; Mustafa, N.S.A.; El-Dahshouri, M.F.; Zhang, L.X.; Ageba, M.F. Efficacy of amended vermicompost for bio-control of root knot nematode (RKN) Meloidogyne incognita infesting tomato in Egypt. Environ. Technol. Innov. 2022, 27, 102397. [Google Scholar] [CrossRef]
- Fan, H.Y.; Yao, M.L.; Wang, H.M.; Zhao, D.; Zhu, X.F.; Wang, Y.Y.; Liu, X.Y.; Duan, Y.X.; Chen, L.J. Isolation and effect of Trichoderma citrinoviride Snef1910 for the biological control of root-knot nematode, Meloidogyne incognita. BMC Microbiol. 2020, 20, 299. [Google Scholar] [CrossRef]
- Ye, S.; Yan, R.; Li, X.W.; Lin, Y.F.; Yang, Z.H.; Ma, Y.H.; Ding, Z. Biocontrol potential of Pseudomonas rhodesiae GC-7 against the root-knot nematode Meloidogyne graminicola through both antagonistic effects and induced plant resistance. Front. Microbiol. 2022, 13, 1025727. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Breitwieser, F.P.; Thielen, P.; Salzberg, S.L. Bracken: Estimating species abundance in metagenomics data. PeerJ Comput. Sci. 2016, 3, e104. [Google Scholar] [CrossRef]
- Franzosa, E.A.; McIver, L.J.; Rahnavard, G.; Thompson, L.R.; Schirmer, M.; Weingart, G.; Lipson, K.S.; Knight, R.; Caporaso, J.G.; Segata, N.; et al. Species-level functional profiling of metagenomes and metatranscriptomes. Nat. Methods 2018, 15, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Sharma, I.P.; Sharma, A.K. Physiological and biochemical changes in tomato cultivar PT-3 with dual inoculation of mycorrhiza and PGPR against root-knot nematode. Symbiosis 2017, 71, 175–183. [Google Scholar] [CrossRef]
- Sharma, N.; Khanna, K.; Manhas, R.K.; Bhardwaj, R.; Ohri, P.; Alkahtani, J.; Alwahibi, M.S.; Ahmad, P. Insights into the role of Streptomyces hydrogenans as the plant growth promoter, photosynthetic pigment enhancer and biocontrol agent against Meloidogyne incognita in Solanum lycopersicum seedlings. Plants 2020, 9, 1109. [Google Scholar] [CrossRef]
- Kaur, T.; Jasrotia, S.; Ohri, P.; Manhas, R.K. Evaluation of in vitro and in vivo nematicidal potential of a multifunctional streptomycete, Streptomyces hydrogenans strain DH16 against Meloidogyne incognita. Microbiol. Res. 2016, 192, 247–252. [Google Scholar] [CrossRef]
- Chaurasia, A.; Meena, B.R.; Tripathi, A.N.; Pandey, K.K.; Rai, A.B.; Singh, B. Actinomycetes: An unexplored microorganisms for plant growth promotion and biocontrol in vegetable crops. World J. Microbiol. Biotechnol. 2018, 34, 132. [Google Scholar] [CrossRef]
- Liu, C.; Yu, J.; Ying, J.; Zhang, K.; Hu, Z.; Liu, Z.; Chen, S. Integrated metagenomics and metabolomics analysis reveals changes in the microbiome and metabolites in the rhizosphere soil of Fritillaria unibracteata. Front. Plant Sci. 2023, 14, 1223720. [Google Scholar] [CrossRef]
- Wu, L.; Wang, Y.; Lyu, H.; Chen, X. Effects of a compound Trichoderma agent on Coptis chinensis growth, nutrients, enzyme activity, and microbial community of rhizosphere soil. PeerJ 2023, 11, e15652. [Google Scholar] [CrossRef]
- Zhang, F.; Huo, Y.; Cobb, A.B.; Luo, G.; Zhou, J.; Yang, G.; Wilson, G.W.T.; Zhang, Y. Trichoderma biofertilizer links to altered soil chemistry, altered microbial communities, and improved grassland biomass. Front. Microbiol. 2018, 9, 848. [Google Scholar] [CrossRef]
- Halifu, S.; Deng, X.; Song, X.; Song, R. Effects of two Trichoderma strains on plant growth, rhizosphere soil nutrients, and fungal community of Pinus sylvestris var. mongolica annual seedlings. Forests 2019, 10, 758. [Google Scholar] [CrossRef]
- Zheng, L.; Wang, S.; Gu, X.; Gao, A.; Liu, L.; Wu, X.; Pan, H.; Zhang, H. Pantoea jilinensis D25 enhances tomato salt tolerance via altering antioxidant responses and soil microbial community structure. Environ. Res. 2024, 243, 117846. [Google Scholar] [CrossRef] [PubMed]
- Jie, W.; Yao, Y.; Guo, N.; Zhang, Y.; Qiao, W. Effects of Rhizophagus intraradices on plant growth and the composition of microbial communities in the roots of continuous cropping soybean at maturity. Sustainability 2021, 13, 6623. [Google Scholar] [CrossRef]
- Kennedy, A.C.; Smith, K.L. Soil microbial diversity and the sustainability ofagricultural soils. Plant Soil 1995, 170, 75–86. [Google Scholar] [CrossRef]
- Sui, L.; Li, J.; Philp, J.; Yang, K.; Wei, Y.; Li, H.; Li, J.; Li, L.; Ryder, M.; Toh, R.; et al. Trichoderma atroviride seed dressing influenced the fungal community and pathogenic fungi in the wheat rhizosphere. Sci. Rep. 2022, 12, 9677. [Google Scholar] [CrossRef]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Huston, M.A.; Raffaelli, D.; Schmid, B.; et al. Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 2001, 294, 804–808. [Google Scholar] [CrossRef]
- She, S.; Niu, J.; Zhang, C.; Xiao, Y.; Chen, W.; Dai, L.; Liu, X.; Yin, H. Significant relationship between soil bacterial community structure and incidence of bacterial wilt disease under continuous cropping system. Arch. Microbiol. 2017, 199, 267–275. [Google Scholar] [CrossRef]
- Bergstrand, L.H.; Cardenas, E.; Holert, J.; VanHamme, J.D.; Mohn, W.W. Delineation of steroid-degrading microorganisms through comparative genomic analysis. mBio 2016, 7, e00166. [Google Scholar] [CrossRef]
- Bouhss, A.; Trunkfield, A.E.; Bugg, T.D.H.; Mengin-Lecreulx, D. The biosynthesis of peptidoglycan lipid-linked intermediates. FEMS Microbiol Rev. 2008, 32, 208–233. [Google Scholar] [CrossRef]
- Pang, Z.; Chen, J.; Wang, T.; Gao, C.; Li, Z.; Guo, L.; Xu, J.; Cheng, Y. Linking plant secondary metabolites and plant microbiomes: A review. Front Plant Sci. 2021, 12, 621276. [Google Scholar] [CrossRef]
- Wong-Ng, J.; Celani, A.; Vergassola, M. Exploring the function of bacterial chemotaxis. Curr. Opin. Microbiol. 2018, 45, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Núñez, M.A.; Orozco-Ramírez, Q. Characterizing bacterial communities in agroecosystems of the UNESCO global geopark Mixteca Alta, Oaxaca. Agriculture 2024, 14, 2180. [Google Scholar] [CrossRef]
- Wittlinger, J.P.; Castejón, N.; Hausmann, B.; Berry, D.; Schnorr, S.L. Shewanella is a putative producer of polyunsaturated fatty acids in the gut soil of the composting earthworm Eisenia fetida. Appl. Environ. Microbiol. 2025, 91, e0206924. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).