

Genome-Driven Functional Validation of Bacillus amyloliquefaciens Strain MEPW12: A Multifunctional Endophyte for Sustainable Sweet Potato Cultivation

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Isolation of Bacterial Endophytes
2.2. Functional Identification of Endophytic Bacteria
2.2.1. Detection of Phosphorus Solubilization
2.2.2. Detection of Nitrogen Fixation
2.2.3. Detection of Siderophore Production
2.2.4. Detection of 1-Aminocyclopropane-1-Carboxylic Acid (ACC) Deaminase Production
2.2.5. Detection of Cellulase Production
2.2.6. Detection of Indole-3-Acetic Acid (IAA) Production by Strains
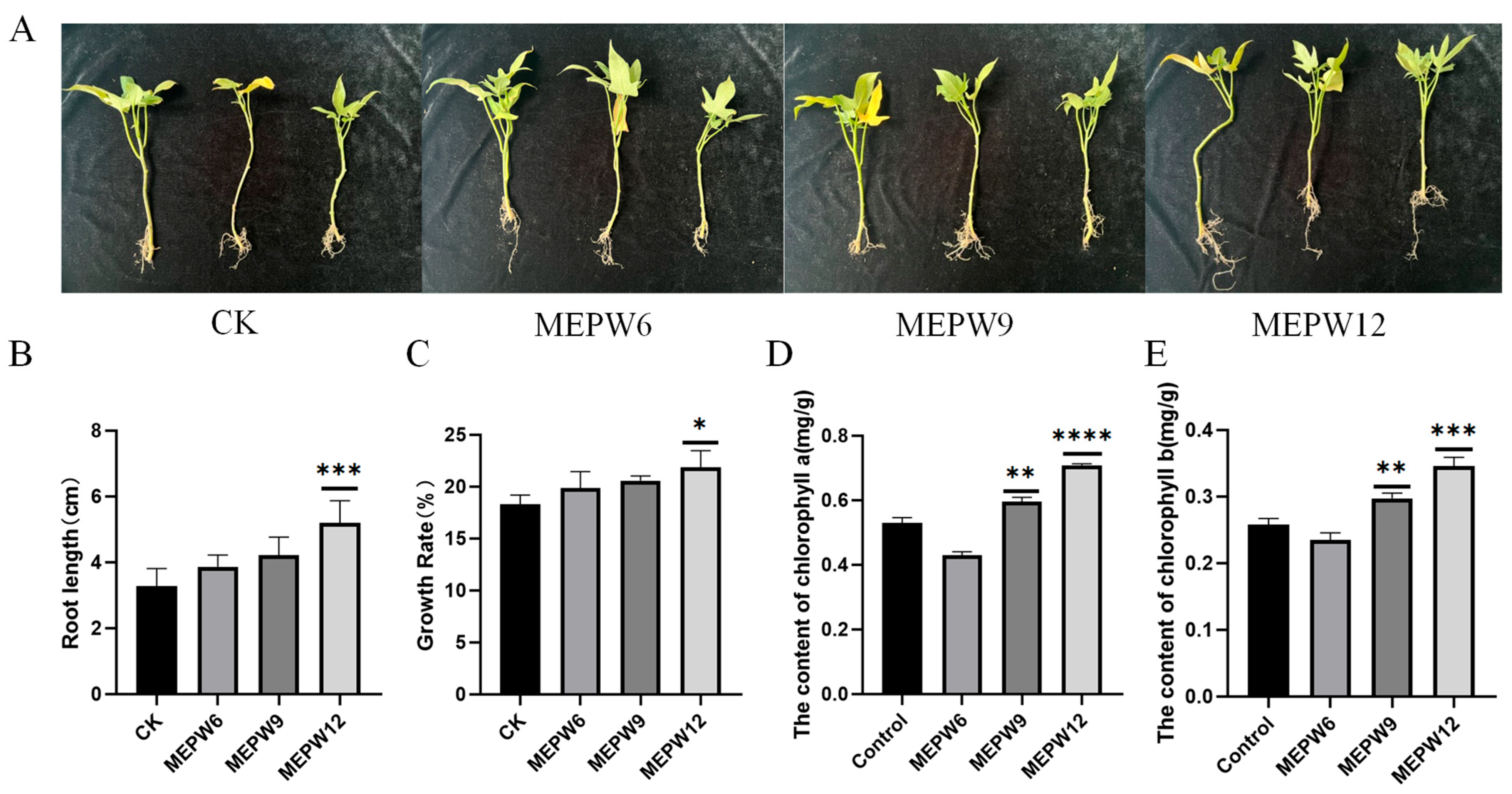
2.3. Plant Growth Promotion Experiment
2.4. Determination of the Chlorophyll Content
2.5. Identification of Growth Promoting Bacteria
2.6. Physiological and Biochemical Identification of MEPW12
2.7. Antagonistic Activity of MEPW12 Against Sweet Potato Pathogens
2.8. Detection of MEPW12 Colonization
2.9. Whole Genome Analysis of Strain MEPW12
2.10. Comparative Genomics and Pan-Genomic Analysis
2.11. RNA Extraction and Quantitative Reverse Transcription PCR (qRT-PCR) Validation
2.12. Data Analysis
3. Results
3.1. Isolation and Functional Identification of Endophytes from Sweet Potatoes
3.2. Molecular Identification of MEPW12 and Identification of Other Physiological and Biochemical Functions
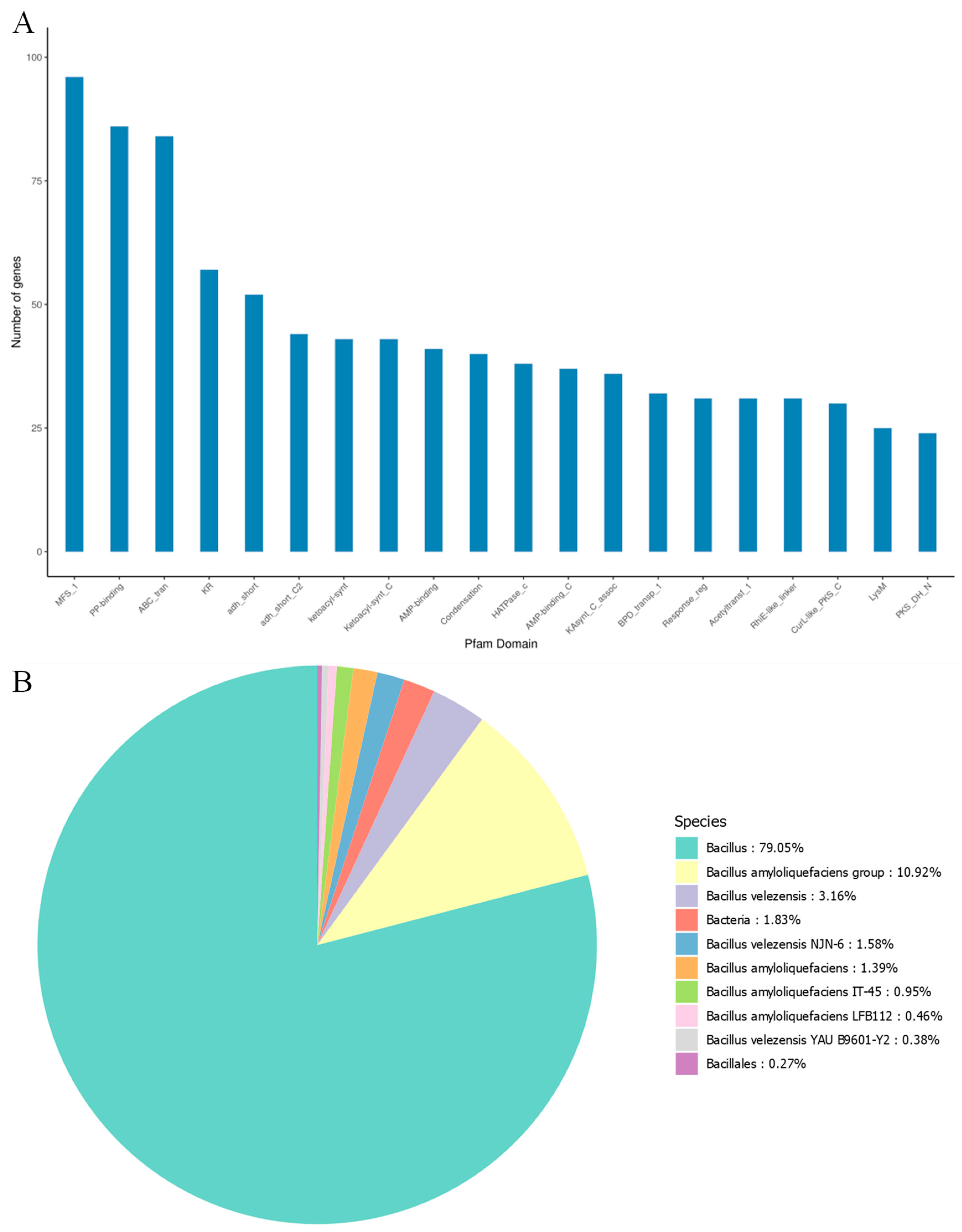
3.3. Genomic, Comparative Genomic, and Pan-Genomic Analyses of MEPW12
3.4. Effect of MEPW12 on the Expression of Growth-Related Genes in Sweet Potatoes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACC | 1-aminocyclopropane-1-carboxylic |
| IAA | Indole-3-acetic acid |
| CMC | Carboxymethyl cellulose |
| CAS | Chrome Azurol S |
| NF | Nitrogen fixation |
| SP | Solubilization of phosphate |
| CK | Control Check |
| qPCR | Quantitative Real-time Polymerase Chain Reaction |
| qRT-PCR | Quantitative Reverse Transcription Polymerase Chain Reaction |
| MYC2 | Myelocytomatosis proteins 2 |
| IAA1 | Indole acetic acid 1 |
| ERF1 | Ethylene responsive factor 1 |
| GH3.10 | Gretchen Hage 3.10 |
| JAZ6 | Jasmonate ZIM-domain 6 |
| PYL-2 | Pyrabactin resistance 1-like 2 |
| AUX1 | Auxin 1 |
| PR-1 | Plant pathogenesis-related protein 1 |
| EIN3 | Ethylene-insensitive 3 |
References
- Chaudhary, P.; Singh, S.; Chaudhary, A.; Sharma, A.; Kumar, G. Overview of biofertilizers in crop production and stress management for sustainable agriculture. Front. Plant Sci. 2022, 13, 930340. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Yang, Y.; Zhang, L.; Zhang, J.; Zhou, Y.; Huang, H.; Luo, S.; Luo, L. Silicon-based additive on heavy metal remediation in soils: Toxicological effects, remediation techniques, and perspectives. Environ. Res. 2022, 205, 112244. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Cong, H.; Li, C.; Wu, J.; Li, L.; Jiang, J.; Cao, X. Cultivable Endophyte Resources in Medicinal Plants and Effects on Hosts. Life 2023, 13, 1695. [Google Scholar] [CrossRef]
- Dwibedi, V.; Rath, S.K.; Joshi, M.; Kaur, R.; Kaur, G.; Singh, D.; Kaur, G.; Kaur, S. Microbial endophytes: Application towards sustainable agriculture and food security. Appl. Microbiol. Biotechnol. 2022, 106, 5359–5384. [Google Scholar] [CrossRef]
- Kashyap, N.; Singh, S.K.; Yadav, N.; Singh, V.K.; Kumari, M.; Kumar, D.; Shukla, L.; Kaushalendra; Bhardwaj, N.; Kumar, A. Biocontrol Screening of Endophytes: Applications and Limitations. Plants 2023, 12, 2480. [Google Scholar] [CrossRef]
- Muhammad, M.; Wahab, A.; Waheed, A.; Mohamed, H.I.; Hakeem, K.R.; Li, L.; Li, W.J. Harnessing bacterial endophytes for environmental resilience and agricultural sustainability. J. Environ. Manag. 2024, 368, 122201. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.; Mandal, S. Unveiling growth-promoting attributes of peanut root endophyte Micromonospora sp. Arch. Microbiol. 2024, 206, 182. [Google Scholar] [CrossRef]
- Morales-Cedeno, L.R.; Orozco-Mosqueda, M.D.C.; Loeza-Lara, P.D.; Parra-Cota, F.I.; de Los Santos-Villalobos, S.; Santoyo, G. Plant growth-promoting bacterial endophytes as biocontrol agents of pre- and post-harvest diseases: Fundamentals, methods of application and future perspectives. Microbiol. Res. 2021, 242, 126612. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Shi, H.; Heng, J.; Wang, D.; Bian, K. Antimicrobial, plant growth-promoting and genomic properties of the peanut endophyte Bacillus velezensis LDO2. Microbiol. Res. 2019, 218, 41–48. [Google Scholar] [CrossRef]
- Liu, A.; Zhang, P.; Bai, B.; Bai, F.; Jin, T.; Ren, J. Volatile Organic Compounds of Endophytic Burkholderia pyrrocinia Strain JK-SH007 Promote Disease Resistance in Poplar. Plant Dis. 2020, 104, 1610–1620. [Google Scholar] [CrossRef]
- Montes-Osuna, N.; Cernava, T.; Gomez-Lama Cabanas, C.; Berg, G.; Mercado-Blanco, J. Identification of Volatile Organic Compounds Emitted by Two Beneficial Endophytic Pseudomonas Strains from Olive Roots. Plants 2022, 11, 318. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.Y.; Sun, K.; Chen, F.; Yuan, J.; Li, X.; Dai, C.C. Endophytic Pseudomonas induces metabolic flux changes that enhance medicinal sesquiterpenoid accumulation in Atractylodes lancea. Plant Physiol. Biochem. 2018, 130, 473–481. [Google Scholar] [CrossRef]
- Liu, Z.; Ma, Y.; Lv, X.; Li, N.; Li, X.; Xing, J.; Li, C.; Hu, B. Abiotic factors and endophytes co-regulate flavone and terpenoid glycoside metabolism in Glycyrrhiza uralensis. Appl. Microbiol. Biotechnol. 2023, 107, 2671–2688. [Google Scholar] [CrossRef] [PubMed]
- Tsipinana, S.; Husseiny, S.; Alayande, K.A.; Raslan, M.; Amoo, S.; Adeleke, R. Contribution of endophytes towards improving plant bioactive metabolites: A rescue option against red-taping of medicinal plants. Front. Plant Sci. 2023, 14, 1248319. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cai, S.; Tao, Z.; Peng, J.; Li, D.; Li, L.; Cao, X.; Jiang, J. Isolation of Endophytic Fungi and Effects on Secondary Metabolites in Hairy Roots of Salvia miltiorrhiza. J. Microbiol. Biotechnol. 2025, 35, e2411051. [Google Scholar] [CrossRef]
- Liu, T.; Xie, Q.; Zhang, M.; Gu, J.; Huang, D.; Cao, Q. Reclaiming Agriceuticals from Sweetpotato (Ipomoea batatas [L.] Lam.) By-Products. Foods 2024, 13, 1180. [Google Scholar] [CrossRef]
- Mattoo, A.K.; Dwivedi, S.L.; Dutt, S.; Singh, B.; Garg, M.; Ortiz, R. Anthocyanin-Rich Vegetables for Human Consumption-Focus on Potato, Sweetpotato and Tomato. Int. J. Mol. Sci. 2022, 23, 2634. [Google Scholar] [CrossRef]
- Mollinari, M.; Olukolu, B.A.; Pereira, G.D.S.; Khan, A.; Gemenet, D.; Yencho, G.C.; Zeng, Z.B. Unraveling the Hexaploid Sweetpotato Inheritance Using Ultra-Dense Multilocus Mapping. G3 2020, 10, 281–292. [Google Scholar] [CrossRef]
- Pountain, A.W.; Jiang, P.; Yao, T.; Homaee, E.; Guan, Y.; McDonald, K.J.C.; Podkowik, M.; Shopsin, B.; Torres, V.J.; Golding, I.; et al. Transcription-replication interactions reveal bacterial genome regulation. Nature 2024, 626, 661–669. [Google Scholar] [CrossRef]
- Hiralal, A.; Geelhoed, J.S.; Hidalgo-Martinez, S.; Smets, B.; van Dijk, J.R.; Meysman, F.J.R. Closing the genome of unculturable cable bacteria using a combined metagenomic assembly of long and short sequencing reads. Microb. Genom. 2024, 10, 001197. [Google Scholar] [CrossRef]
- Srinivasan, S. Radiation-Tolerant Fibrivirga spp. from Rhizosphere Soil: Genome Insights and Potential in Agriculture. Genes. 2024, 15, 1048. [Google Scholar] [CrossRef]
- Sarawad, A.; Hosagoudar, S.; Parvatikar, P. Pan-genomics: Insight into the Functional Genome, Applications, Advancements, and Challenges. Curr. Genom. 2025, 26, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Ren, H.; Wang, X.; Han, C.; Jin, Y.; Hu, X.; Shi, R.; Li, C.; Wang, Y.; Li, Y.; et al. Comparative genomics analysis to explore the biodiversity and mining novel target genes of Listeria monocytogenes strains from different regions. Front. Microbiol. 2024, 15, 1424868. [Google Scholar] [CrossRef] [PubMed]
- Okubo, T.; Ikeda, S.; Kaneko, T.; Eda, S.; Mitsui, H.; Sato, S.; Tabata, S.; Minamisawa, K. Nodulation-dependent communities of culturable bacterial endophytes from stems of field-grown soybeans. Microbes Environ. 2009, 24, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Amri, M.; Rjeibi, M.R.; Gatrouni, M.; Mateus, D.M.R.; Asses, N.; Pinho, H.J.O.; Abbes, C. Isolation, Identification, and Characterization of Phosphate-Solubilizing Bacteria from Tunisian Soils. Microorganisms 2023, 11, 783. [Google Scholar] [CrossRef]
- Kayasth, M.; Gera, R.; Dudeja, S.S.; Sharma, P.K.; Kumar, V. Studies on salinization in Haryana soils on free-living nitrogen-fixing bacterial populations and their activity. J. Basic. Microbiol. 2014, 54, 170–179. [Google Scholar] [CrossRef]
- Himpsl, S.D.; Mobley, H.L.T. Siderophore Detection Using Chrome Azurol S and Cross-Feeding Assays. Methods Mol. Biol. 2019, 2021, 97–108. [Google Scholar] [CrossRef]
- Moon, Y.S.; Ali, S. Isolation and identification of multi-trait plant growth-promoting rhizobacteria from coastal sand dune plant species of Pohang beach. Folia Microbiol. 2022, 67, 523–533. [Google Scholar] [CrossRef]
- Wang, J.; Qin, S.; Fan, R.; Peng, Q.; Hu, X.; Yang, L.; Liu, Z.; Baccelli, I.; Migheli, Q.; Berg, G.; et al. Plant Growth Promotion and Biocontrol of Leaf Blight Caused by Nigrospora sphaerica on Passion Fruit by Endophytic Bacillus subtilis Strain GUCC4. J. Fungi 2023, 9, 132. [Google Scholar] [CrossRef]
- Gang, S.; Sharma, S.; Saraf, M.; Buck, M.; Schumacher, J. Analysis of Indole-3-acetic Acid (IAA) Production in Klebsiellaby LC-MS/MS and the Salkowski Method. Bio Protoc. 2019, 9, e3230. [Google Scholar] [CrossRef]
- Kan, Y.; Zhang, L.; Wang, Y.; Ma, Q.; Zhou, Y.; Jiang, X.; Zhang, W.; Ruan, Z. Endophytic Bacterium Flexivirga meconopsidis sp. nov. with Plant Growth-Promoting Function, Isolated from the Seeds of Meconopsis integrifolia. Microorganisms 2023, 11, 2899. [Google Scholar] [CrossRef] [PubMed]
- Ntemafack, A.; Ahmed, S.; Kumar, A.; Chouhan, R.; Kapoor, N.; Bharate, S.B.; Hassan, Q.P.; Gandhi, S.G. Plant growth promoting potential of butyl isobutyl phthalate and Streptomyces sp. from Rumex dentatus on rice. Appl. Microbiol. Biotechnol. 2022, 106, 2603–2617. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Li, F.; Li, M.; He, Y.; Chen, Y.; Hu, F. Functional analysis of ARF1 from Cymbidium goeringii in IAA response during leaf development. PeerJ 2022, 10, e13077. [Google Scholar] [CrossRef]
- Porra, R.J. The chequered history of the development and use of simultaneous equations for the accurate determination of chlorophylls a and b. Photosynth. Res. 2002, 73, 149–156. [Google Scholar] [CrossRef]
- Zhang, Y.; Cao, X.; Liu, Q.; Chen, Y.; Wang, Y.; Cong, H.; Li, C.; Li, Y.; Wang, Y.; Jiang, J.; et al. Multi-omics analysis of Streptomyces djakartensis strain MEPS155 reveal a molecular response strategy combating Ceratocystis fimbriata causing sweet potato black rot. Food Microbiol. 2024, 122, 104557. [Google Scholar] [CrossRef]
- Corona Ramirez, A.; Symanczik, S.; Gallusser, T.; Bodenhausen, N. Quantification of arbuscular mycorrhizal fungi root colonization in wheat, tomato, and leek using absolute qPCR. Mycorrhiza 2023, 33, 387–397. [Google Scholar] [CrossRef] [PubMed]
- de Paula, C.C.P.; Barta, J.; Borovec, J.; Frouz, J.; Rychtecky, P.; Sirova, D. qPCR-based quantification reveals high plant host-specificity of endophytic colonization levels in leaves. Am. J. Bot. 2025, 112, e16448. [Google Scholar] [CrossRef]
- Liu, Y.; Stefanic, P.; Miao, Y.; Xue, Y.; Xun, W.; Zhang, N.; Shen, Q.; Zhang, R.; Xu, Z.; Mandic-Mulec, I. Housekeeping gene gyrA, a potential molecular marker for Bacillus ecology study. AMB Express 2022, 12, 133. [Google Scholar] [CrossRef]
- Yang, P.; Zeng, Q.; Jiang, W.; Wang, L.; Zhang, J.; Wang, Z.; Wang, Q.; Li, Y. Genome Sequencing and Characterization of Bacillus velezensis N23 as Biocontrol Agent against Plant Pathogens. Microorganisms 2024, 12, 294. [Google Scholar] [CrossRef]
- Fang, L.; Zheng, X.; Sun, Z.; Li, Y.; Deng, J.; Zhou, Y.I. Characterization of a Plant Growth-Promoting Endohyphal Bacillus subtilis in Fusarium acuminatum from Spiranthes sinensis. Pol. J. Microbiol. 2023, 72, 29–37. [Google Scholar] [CrossRef]
- Li, R.; Yang, P.; Zhang, H.; Wang, C.; Zhao, F.; Liu, J.; Wang, Y.; Liang, Y.; Sun, T.; Xie, X. Comparative Genomic and Functional Analysis of c-di-GMP Metabolism and Regulatory Proteins in Bacillus velezensis LQ-3. Microorganisms 2024, 12, 1724. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Liang, X.; Shi, J.C.; Luo, J.N.; Qiu, L.J.; Li, X.X.; Lu, L.J.; Wen, Y.Q.; Feng, J.Y. Pan-Genomic Analysis Identifies the Chinese Strain as a New Subspecies of Xanthomonas fragariae. Plant Dis. 2024, 108, 45–49. [Google Scholar] [CrossRef]
- Harman, G.E.; Uphoff, N. Symbiotic Root-Endophytic Soil Microbes Improve Crop Productivity and Provide Environmental Benefits. Scientifica 2019, 2019, 9106395. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.; Khan, Q.M.; Sessitsch, A. Endophytic bacteria: Prospects and applications for the phytoremediation of organic pollutants. Chemosphere 2014, 117, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Saikia, J.; Thakur, D. A review on endophytic bacteria of orchids: Functional roles toward synthesis of bioactive metabolites for plant growth promotion and disease biocontrol. Planta 2024, 260, 70. [Google Scholar] [CrossRef]
- Thoa, N.T.K.; Mai, D.T.H.; Hiu, B.L.; Duong, C.A.; Chau, N.N.B.; Nghiep, N.M.; Van Minh, N.; Quoc, N.B. Roles of β-Indole Acetic Acid (IAA) Producing Endophytic Bacteria on the Recovery of Plant Growth and Survival Ability of Sugarcane Infected White Leaf Disease (SWLD). Curr. Microbiol. 2022, 79, 389. [Google Scholar] [CrossRef]
- Etesami, H.; Alikhani, H.A.; Hosseini, H.M. Indole-3-acetic acid (IAA) production trait, a useful screening to select endophytic and rhizosphere competent bacteria for rice growth promoting agents. MethodsX 2015, 2, 72–78. [Google Scholar] [CrossRef]
- Yuan, Z.S.; Liu, F.; Zhang, G.F. Characteristics and biodiversity of endophytic phosphorus- and potassium-solubilizing bacteria in Moso Bamboo (Phyllostachys edulis). Acta Biol. Hung. 2015, 66, 449–459. [Google Scholar] [CrossRef]
- Varga, T.; Hixson, K.K.; Ahkami, A.H.; Sher, A.W.; Barnes, M.E.; Chu, R.K.; Battu, A.K.; Nicora, C.D.; Winkler, T.E.; Reno, L.R.; et al. Endophyte-Promoted Phosphorus Solubilization in Populus. Front. Plant Sci. 2020, 11, 567918. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Peng, J.; Chen, C.; Xiong, C.; Li, S.; Ge, A.; Wang, E.; Liesack, W. Harnessing biological nitrogen fixation in plant leaves. Trends Plant Sci. 2023, 28, 1391–1405. [Google Scholar] [CrossRef] [PubMed]
- Orozco-Mosqueda, M.D.C.; Glick, B.R.; Santoyo, G. ACC deaminase in plant growth-promoting bacteria (PGPB): An efficient mechanism to counter salt stress in crops. Microbiol. Res. 2020, 235, 126439. [Google Scholar] [CrossRef]
- Rosenblueth, M.; Ormeno-Orrillo, E.; Lopez-Lopez, A.; Rogel, M.A.; Reyes-Hernandez, B.J.; Martinez-Romero, J.C.; Reddy, P.M.; Martinez-Romero, E. Nitrogen Fixation in Cereals. Front. Microbiol. 2018, 9, 1794. [Google Scholar] [CrossRef]
- Khan, T.; Alzahrani, O.M.; Sohail, M.; Hasan, K.A.; Gulzar, S.; Rehman, A.U.; Mahmoud, S.F.; Alswat, A.S.; Abdel-Gawad, S.A. Enzyme Profiling and Identification of Endophytic and Rhizospheric Bacteria Isolated from Arthrocnemum macrostachyum. Microorganisms 2022, 10, 2112. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wei, S.; Chen, X.; Dong, Y.; Zeng, M.; Yan, C.; Hou, L.; Jiao, R. Isolation of cadmium-resistance and siderophore-producing endophytic bacteria and their potential use for soil cadmium remediation. Heliyon 2023, 9, e17661. [Google Scholar] [CrossRef]
- Khan, A.R.; Mustafa, A.; Hyder, S.; Valipour, M.; Rizvi, Z.F.; Gondal, A.S.; Yousuf, Z.; Iqbal, R.; Daraz, U. Bacillus spp. as Bioagents: Uses and Application for Sustainable Agriculture. Biology 2022, 11, 1763. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.O.; Bignon, J.; Rapoport, G.; Debarbouille, M. Regulation of the acetoin catabolic pathway is controlled by sigma L in Bacillus subtilis. J. Bacteriol. 2001, 183, 2497–2504. [Google Scholar] [CrossRef]
- Asif, M.; Li-Qun, Z.; Zeng, Q.; Atiq, M.; Ahmad, K.; Tariq, A.; Al-Ansari, N.; Blom, J.; Fenske, L.; Alodaini, H.A.; et al. Comprehensive genomic analysis of Bacillus paralicheniformis strain BP9, pan-genomic and genetic basis of biocontrol mechanism. Comput. Struct. Biotechnol. J. 2023, 21, 4647–4662. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Ayilara, M.S.; Ayangbenro, A.S.; Babalola, O.O. Genome Mining of Three Plant Growth-Promoting Bacillus Species from Maize Rhizosphere. Appl. Biochem. Biotechnol. 2021, 193, 3949–3969. [Google Scholar] [CrossRef]
- Pitiwittayakul, N.; Wongsorn, D.; Tanasupawat, S. Characterisation of Plant Growth-Promoting Endophytic Bacteria from Sugarcane and Their Antagonistic Activity against Fusarium moniliforme. Trop. Life Sci. Res. 2021, 32, 97–118. [Google Scholar] [CrossRef]
- Tashan, H.; Harighi, B.; Rostamzadeh, J.; Azizi, A. Characterization of Arsenic-Resistant Endophytic Bacteria From Alfalfa and Chickpea Plants. Front. Plant Sci. 2021, 12, 696750. [Google Scholar] [CrossRef] [PubMed]
- Johari, N.A.F.; Abidin, A.A.Z.; Ismail, N.F.N.; Yusof, Z.N.B. Endophytic Bacteria Induce Thiamine (Vitamin B1) Production in Oil Palm (Elaeis guineensis). Trop. Life Sci. Res. 2024, 35, 1–12. [Google Scholar] [CrossRef]
- Wojtaczka, P.; Ciarkowska, A.; Starzynska, E.; Ostrowski, M. The GH3 amidosynthetases family and their role in metabolic crosstalk modulation of plant signaling compounds. Phytochemistry 2022, 194, 113039. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Li, T.T.; Shi, W.M.; Ma, Q.; Di, D.W. The Roles of GRETCHEN HAGEN3 (GH3)-Dependent Auxin Conjugation in the Regulation of Plant Development and Stress Adaptation. Plants 2023, 12, 4111. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Hu, Y.; Aoi, Y.; Hira, H.; Ge, C.; Dai, X.; Kasahara, H.; Zhao, Y. Local conjugation of auxin by the GH3 amido synthetases is required for normal development of roots and flowers in Arabidopsis. Biochem. Biophys. Res. Commun. 2022, 589, 16–22. [Google Scholar] [CrossRef]
- Li, Y.; Han, S.; Qi, Y. Advances in structure and function of auxin response factor in plants. J. Integr. Plant Biol. 2023, 65, 617–632. [Google Scholar] [CrossRef]
- Singla, B.; Chugh, A.; Khurana, J.P.; Khurana, P. An early auxin-responsive Aux/IAA gene from wheat (Triticum aestivum) is induced by epibrassinolide and differentially regulated by light and calcium. J. Exp. Bot. 2006, 57, 4059–4070. [Google Scholar] [CrossRef]
- Lv, K.; Wu, W.; Wei, H.; Liu, G. A systems biology approach identifies a regulator, BplERF1, of cold tolerance in Betula platyphylla. For. Res. 2021, 1, 11. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, J.; Li, Z.; Qiao, J.; Quan, R.; Wang, J.; Huang, R.; Qin, H. SALT AND ABA RESPONSE ERF1 improves seed germination and salt tolerance by repressing ABA signaling in rice. Plant Physiol. 2022, 189, 1110–1127. [Google Scholar] [CrossRef]
- Fall, A.F.; Nakabonge, G.; Ssekandi, J.; Founoune-Mboup, H.; Apori, S.O.; Ndiaye, A.; Badji, A.; Ngom, K. Roles of Arbuscular Mycorrhizal Fungi on Soil Fertility: Contribution in the Improvement of Physical, Chemical, and Biological Properties of the Soil. Front. Fungal Biol. 2022, 3, 723892. [Google Scholar] [CrossRef]
- Chang, J.; van Veen, J.A.; Tian, C.; Kuramae, E.E. A review on the impact of domestication of the rhizosphere of grain crops and a perspective on the potential role of the rhizosphere microbial community for sustainable rice crop production. Sci. Total Environ. 2022, 842, 156706. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Rajkumar, M.; Zhang, C.; Freitas, H. Beneficial role of bacterial endophytes in heavy metal phytoremediation. J. Environ. Manag. 2016, 174, 14–25. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | FN | SP | CMC | ACC | Siderophore | IAA Concentration (mg/L, Average) |
|---|---|---|---|---|---|---|
| MEPW1 | + | − | − | − | − | 20.56 |
| MEPW2 | + | − | − | + | + | 13.32 |
| MEPW3 | + | − | − | + | + | 12.75 |
| MEPW4 | − | − | + | − | − | 10.82 |
| MEPW5 | + | − | + | + | + | 5.29 |
| MEPW6 | + | − | + | + | + | 7.54 |
| MEPW7 | + | − | − | − | − | 15.12 |
| MEPW8 | + | − | − | + | + | 16.91 |
| MEPW9 | + | − | + | + | + | 13.99 |
| MEPW10 | + | − | + | + | − | 12.22 |
| MEPW11 | − | − | − | + | + | 10.97 |
| MEPW12 | + | + | + | + | + | 13.94 |
| Functions | Results |
|---|---|
| V–P | + |
| Citrate | + |
| Propionate | − |
| D-xylose | + |
| L-arabinose | + |
| D-mannitol | − |
| Gelatin liquefaction | + |
| Growth with 7% NaCl | + |
| Growth with pH5.7 | + |
| Nitrate reduction | + |
| Starch hydrolysis | + |
| Anaerobic growth | + |
| PGP Traits | Genes |
|---|---|
| IAA | trpA trpB trpC trpD trpE trpF |
| Siderophore | fepC ftsY ftsX ftsE |
| Nitrogen generation | glnA glnN glnH glnP glnM glnT glnK glnL nasB nasD nasE gltB gltD gltX |
| Phosphate solubilization and uptake | pstA pstB pstC pstS phnP phoD phoB1 phoP phoR ugpQ |
| Class | Num | Descrition |
|---|---|---|
| GHs | 40 | Glycoside Hydrolases |
| GTs | 35 | Glycosyltransferases |
| PLs | 3 | Polysaccharide Lyases |
| CEs | 17 | Carbohydrate Esterases |
| AAs | 6 | Auxiliary Activities |
| CBMs | 4 | Carbohydrate-Binding Modules |
| Region | Type | From | To | Most Similar Known Cluster | Compound | Similarity |
|---|---|---|---|---|---|---|
| Region1 | NRPS | 304,470 | 369,877 | NRP:Lipopeptide | surfactin | 78% |
| Region2 | thiopeptide, LAP | 585,389 | 614,502 | Polyketide | kijanimicin | 4% |
| Region3 | RRE-containing, LAP | 697,719 | 720,896 | RiPP:LAP | plantazolicin | 91% |
| Region4 | PKS-like | 925,178 | 966,422 | Saccharide | butirosin A/butirosin B | 7% |
| Region5 | terpene | 1,048,459 | 1,069,199 | - | - | - |
| Region6 | transAT-PKS | 1,367,308 | 1,455,520 | Polyketide | macrolactin H | 100% |
| Region7 | transAT-PKS, T3PKS, NRPS | 1,674,324 | 1,784,435 | Polyketide + NRP | bacillaene | 100% |
| Region8 | NRPS, transAT-PKS, betalactone | 1,841,080 | 1,978,908 | NRP | fengycin | 100% |
| Region9 | terpene | 2,001,477 | 2,023,360 | - | - | - |
| Region10 | T3PKS | 2,096,195 | 2,137,295 | - | - | - |
| Region11 | transAT-PKS | 2,294,633 | 2,400,811 | Polyketide | difficidin | 100% |
| Region12 | NRP-metallophore | 3,030,529 | 3,082,321 | NRP | bacillibactin | 100% |
| Region13 | other | 3,599,564 | 3,640,982 | Other | bacilysin | 100% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Hao, J.; Gu, J.; Wu, J.; Zhang, Y.; Liang, T.; Bai, H.; Cao, Q.; Jiang, J.; Li, L.; et al. Genome-Driven Functional Validation of Bacillus amyloliquefaciens Strain MEPW12: A Multifunctional Endophyte for Sustainable Sweet Potato Cultivation. Microorganisms 2025, 13, 1322. https://doi.org/10.3390/microorganisms13061322
Wang Y, Hao J, Gu J, Wu J, Zhang Y, Liang T, Bai H, Cao Q, Jiang J, Li L, et al. Genome-Driven Functional Validation of Bacillus amyloliquefaciens Strain MEPW12: A Multifunctional Endophyte for Sustainable Sweet Potato Cultivation. Microorganisms. 2025; 13(6):1322. https://doi.org/10.3390/microorganisms13061322
Chicago/Turabian StyleWang, Yiming, Jingwen Hao, Jingsheng Gu, Jiaying Wu, Yongjing Zhang, Ting Liang, Haimeng Bai, Qinghe Cao, Jihong Jiang, Ludan Li, and et al. 2025. "Genome-Driven Functional Validation of Bacillus amyloliquefaciens Strain MEPW12: A Multifunctional Endophyte for Sustainable Sweet Potato Cultivation" Microorganisms 13, no. 6: 1322. https://doi.org/10.3390/microorganisms13061322
APA StyleWang, Y., Hao, J., Gu, J., Wu, J., Zhang, Y., Liang, T., Bai, H., Cao, Q., Jiang, J., Li, L., & Cao, X. (2025). Genome-Driven Functional Validation of Bacillus amyloliquefaciens Strain MEPW12: A Multifunctional Endophyte for Sustainable Sweet Potato Cultivation. Microorganisms, 13(6), 1322. https://doi.org/10.3390/microorganisms13061322