
Optimization of Fermentation Conditions for Endophytic Fungi from Schisandra chinensis and Investigation of Their Antibacterial Mechanisms Against Methicillin-Resistant Staphylococcus aureus

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Screening of Endophytic Fungi from Schisandra chinensis
2.2. Identification of Antagonistic Fungi
2.3. Inhibition Spectrum of the Antagonistic Fungi
2.4. Screening of Optimum Ingredient for Fermentation Medium
2.5. Effects of Fermentation Parameters on the Antibacterial Activity of S10
2.6. Response Surface Optimization Test
2.7. Preparation and Separation of Antibacterial Extracts
2.8. Determination of Minimum Inhibitory Concentration (MIC) of S10 Against MRSA
2.9. Effects of the Crude Extract on the Growth Curve of MRSA
2.10. Effects of Crude Extract on Alkaline Phosphatase (AKP) Content in Culture Medium
2.11. Effects of Crude Extract on the Membrane Potential of MRSA
2.12. Determination of Biofilm Eradication Potential
2.13. Determination of Effects of Crude Extract on Cell Morphology of MRSA
3. Results
3.1. Screening and Identification of MRSA Antagonistic Fungi
3.2. Inhibition Spectrum of S10
3.3. Optimization Results of Culture Medium Components
3.4. Experimental Analysis of Single-Factor Effects
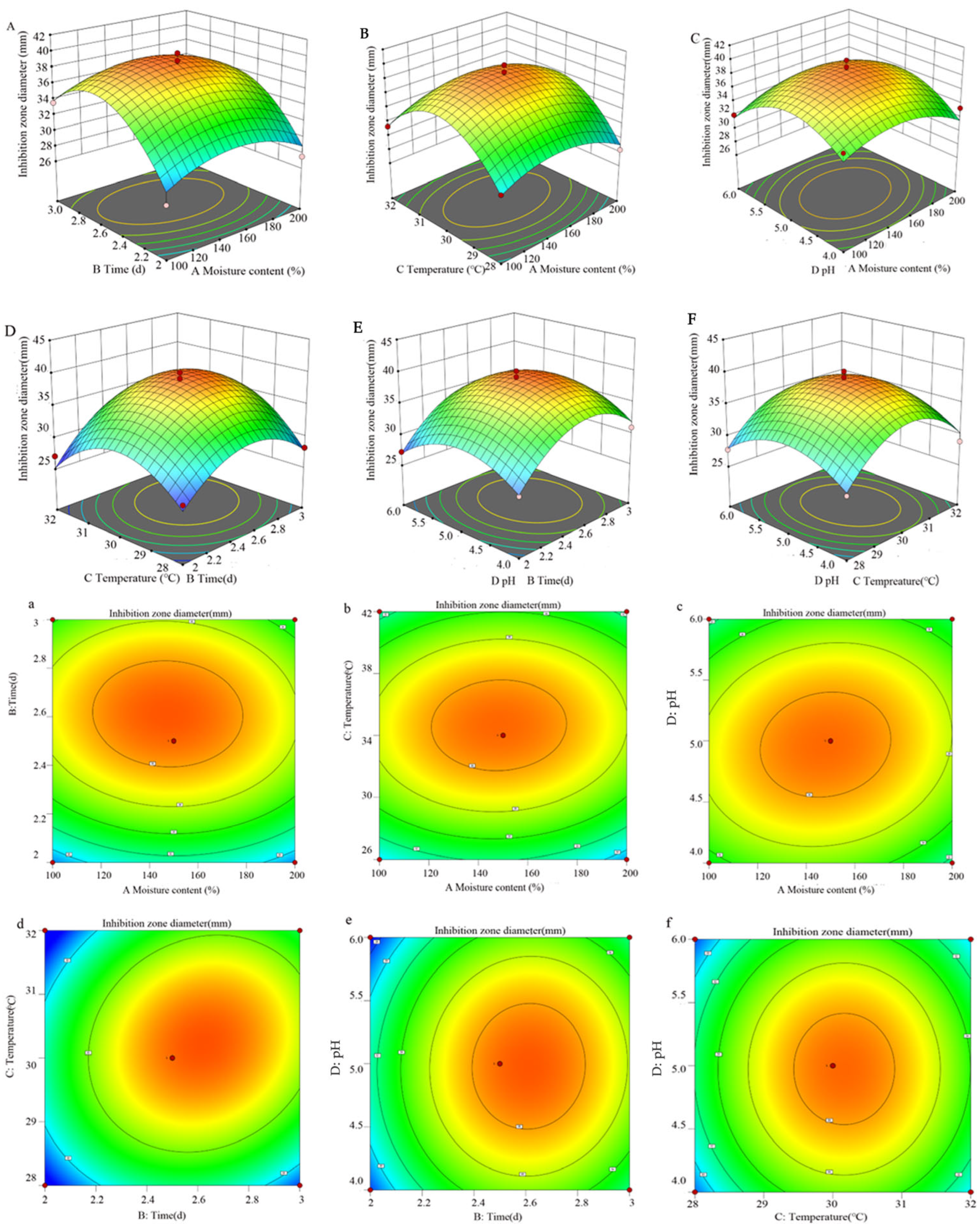
3.5. Optimization Outcomes from Response Surface Methodology Experiments
3.5.1. Regression Equation and Analysis of Variance
3.5.2. Response Surface Analysis of Interaction of Various Factors
3.6. MIC Determination of Crude Extract
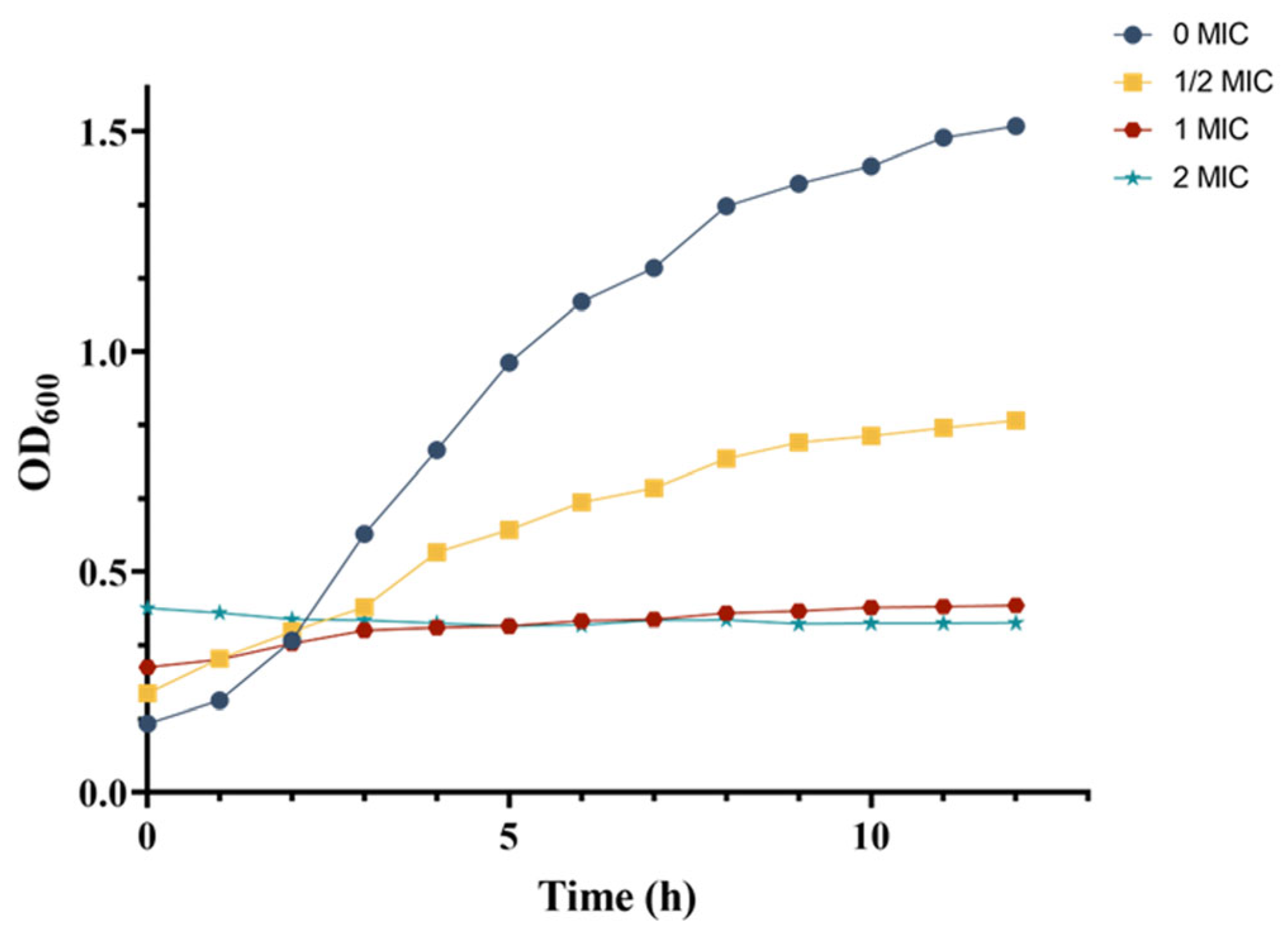
3.7. Effect of Crude Extract on Growth Curve of MRSA
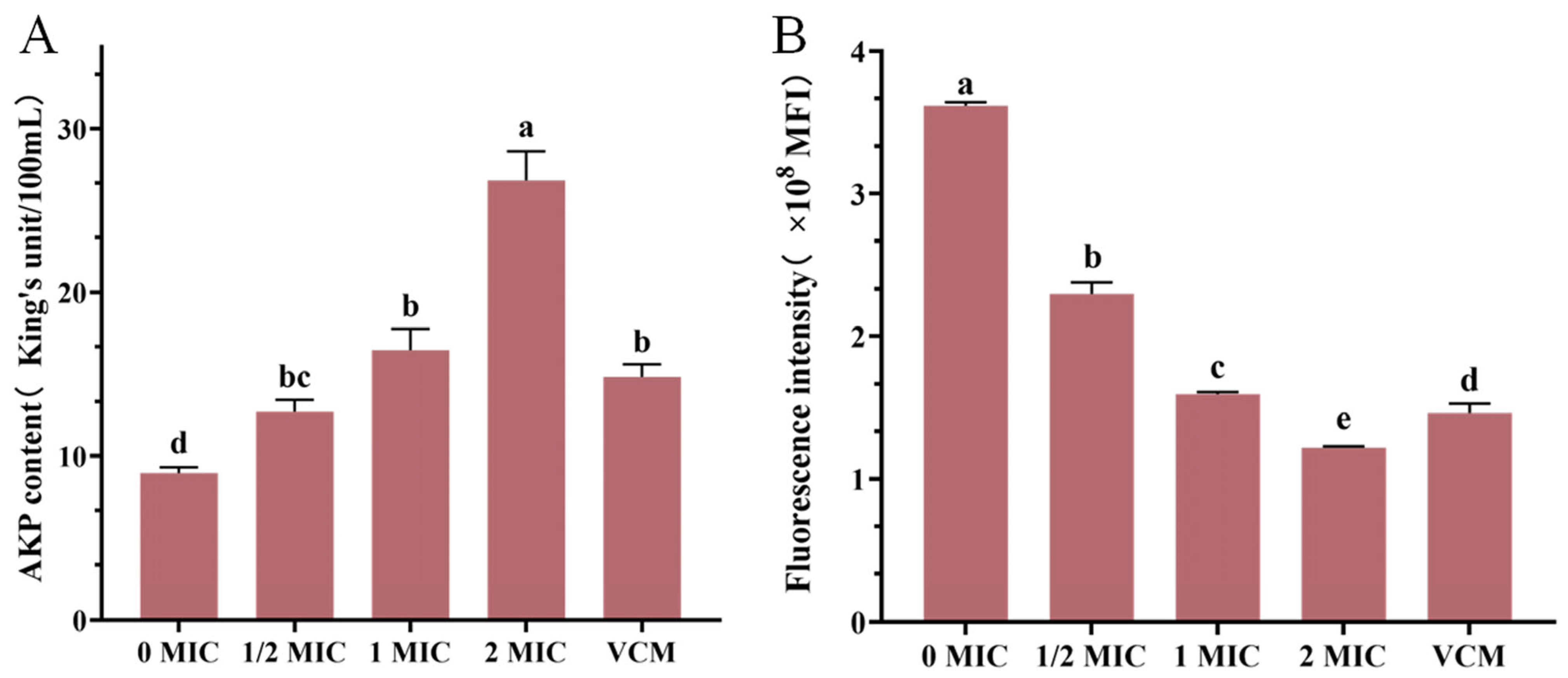
3.8. Results of AKP Content Determination
3.9. Results of Membrane Potential Measurement
3.10. Effects of Crude Extract on MRSA Biofilm Formation
3.11. Effects of Crude Extract on Cell Morphology of MRSA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Newsom, S.W.B. Ogston’s coccus. J. Hosp. Infect. 2008, 70, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Cameron, J.K.; Hall, L.; Tong, S.Y.C.; Paterson, D.L.; Halton, K. Incidence of community-onset MRSA in Australia: Least reported where it is most prevalent. Antimicrob. Resist. Infect. Control 2019, 8, 33. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Wu, Z.; McClements, D.J.; Zou, L.; Liu, W. Improvement on stability, loading capacity and sustained release of rhamnolipids modified curcumin liposomes. Colloids Surf. B Biointerfaces 2019, 183, 110460. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Poddar, N.; Panigrahi, K.; Pathi, B.; Nayak, S.R.; Dandapat, R.; Pattnaik, D.; Praharaj, A.K.; Patro, A.R.K. Evaluation of In-Vitro Activity of Ceftaroline Against Methicillin-Resistant Staphylococcus aureus Clinical Isolates. Cureus 2023, 15, e49859. [Google Scholar] [CrossRef]
- Alan, H.; Cian, O. Emerging nanomedicine therapies to counter the rise of methicillin-resistant Staphylococcus aureus. Materials 2018, 11, 321. [Google Scholar] [CrossRef]
- Ortwine, J.K.; Bhavan, K. Morbidity, mortality, and management of methicillin-resistant S. aureus bacteremia in the US: Update on antibacterial choices and understanding. Hosp. Pract. 2020, 46, 64–72. [Google Scholar] [CrossRef]
- Turnidge, J.D.; Kotsanas, D.; Munckhof, W.; Roberts, S.; Bennett, C.M.; Nimmo, G.R.; Coombs, G.W.; Murray, R.J.; Howden, B.; Johnson, P.D.R.; et al. Staphylococcus aureus bacteremia: A major cause of mortality in Australia and New Zealand. Med. J. Aust. 2009, 191, 368–373. [Google Scholar] [CrossRef]
- Peacock, S.J.; Paterson, G.K. Mechanisms of methicillin resistance in Staphylococcus aureus. Annu. Rev. Biochem. 2015, 84, 577–601. [Google Scholar] [CrossRef]
- Yoneda, A.; Thänert, R.; Burnham, C.A.D.; Dantas, G. In vitro activity of meropenem/piperacillin/tazobactam triple combination therapy against clinical isolates of Staphylococcus aureus, Staphylococcus epidermidis, Staphylococcus pseudintermedius, and vancomycin-resistant Enterococcus spp. Int. J. Antimicrob. Agents 2020, 55, 105864. [Google Scholar] [CrossRef]
- Chen, X.H.; Liu, P.P.; Luo, X.F.; Huang, A.L.; Wan, J.Q. Study on the antibacterial activity and mechanism of cinnamaldehyde against methicillin-resistant Staphylococcus aureus. Eur. Food Res. Technol. 2024, 250, 1069–1081. [Google Scholar] [CrossRef]
- Qin, D.; Wang, L.; Han, M.; Wang, J.; Song, H.; Yan, X.; Duan, X.; Dong, J. Effects of an Endophytic Fungus Umbelopsis dimorpha on the Secondary Metabolites of Host–Plant Kadsura angustifolia. Front. Microbiol. 2018, 9, 2845. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.A.; Abdessamad, D.; Peter, P. Fungal endophytes: Unique plant inhabitants with great promises. Appl. Microbiol. Biotechnol. 2011, 90, 1829–1845. [Google Scholar]
- Ying, G.; Jia, Z.; Hanli, R. Trichothecenes from an Endophytic Fungus Alternaria sp. sb23. Planta Medica 2020, 86, 976–982. [Google Scholar]
- Xu, W.; Zhang, L.-Y.; Goodwin, P.-H.; Xia, M.-C.; Zhang, J.; Wang, Q.; Liang, J.; Sun, R.-H.; Wu, C.; Yang, L.-R. Isolation, identification, and complete genome assembly of an endophytic Bacillus velezensis YB–130, potential biocontrol agent against Fusarium graminearum. Front. Microbiol. 2020, 11, 598285. [Google Scholar] [CrossRef]
- Abdelgawwad, M.R.; Mahmood, A.; Farraj DA, A.; El-Abedein AI, Z.; Mahmoud, A.H.; Bukhari, S.M. In-vitro Antimicrobial Activities of Solanum villosum (L.) Lam; Crude Extract Solvent Comparison. J. King Saud Univ.-Sci. 2020, 32, 2129–2133. [Google Scholar] [CrossRef]
- Santos, S.N.; Ferraris, F.K.; Souza, A.O.; Henriques MD, G.; Melo, I.S. Endophytic fungi from combretum leprosum with potential anticancer and antifungal activity. Symbiosis 2012, 58, 109–117. [Google Scholar] [CrossRef]
- Guenter, S.; Gorkiewicz, G.; Halwachs, B.; Kashofer, K.; Thueringer, A.; Wurm, P.; Zollner-Schwetz, I.; Valenti, T.; Prattes, J.; Wunsch, S. Impact of ITS-based sequencing on antifungal treatment of patients with suspected invasive fungal infections. J. Fungi 2020, 6, 43. [Google Scholar] [CrossRef]
- He, J.; Zhang, X.; Wang, Q.; Li, N.; Ding, D.; Wang, B. Optimization of the fermentation conditions of Metarhizium robertsii and its biological control of wolfberry root rot disease. Microorganisms 2023, 11, 2380. [Google Scholar] [CrossRef]
- Schn, T.; Werngren, J.; Machado, D.; Borroni, E.; Cambau, E. Multicentre testing of the EUCAST broth microdilution reference method for MIC determination on Mycobacterium tuberculosis. Clin. Microbiol. Infect. 2021, 27, 288.e1–288.e4. [Google Scholar] [CrossRef]
- Shu, Q.; Niu, Y.-W.; Zhao, W.-J.; Chen, Q.-H. Antibacterial activity and mannosyl erythritol lipids against vegetative cells and spores of Bacillus cereus. Food Control 2019, 106, 106711. [Google Scholar] [CrossRef]
- Xu, Z.B.; Liang, Y.R.; Lin, S.Q.; Chen, D.Q.; Li, B.; Li, L.; Deng, Y. Crystal violet and XTT assays on Staphylococcus aureus biofilm quantification. Curr. Microbiol. 2016, 73, 474–482. [Google Scholar] [CrossRef]
- Bakkiyaraj, D.; Nandhini, J.R.; Malathy, B.; Pandian, S.K. The anti-biofilm potential of pomegranate (Punica granatum L.) extract against human bacterial and fungal pathogens. Biofouling 2013, 29, 929–937. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.-Y.; Bai, M.; Lin, L. Plasma–treated poly (ethylene oxide) nanofibers containing tea tree oil/beta-cyclodextrin inclusion complex for antibacterial packaging. Carbohydr. Polym. 2018, 179, 360–369. [Google Scholar] [CrossRef]
- Zheng, H.F.; Liu, Y.; Cai, J.; Zhang, M.; Wen, Y.; Guo, L. The exploration of anti-Vibrio parahaemolyticus substances from Phellodendri Chinensis Cortex as a preservative for shrimp storage. Front. Microbiol. 2022, 13, 1004262. [Google Scholar] [CrossRef]
- Lei, G.; Jiacai, G.; Fuquan, X. Optimized extraction process and identification of antibacterial substances from Rhubarb against aquatic pathogenic Vibrio harveyi. 3 Biotech 2017, 7, 377. [Google Scholar]
- Kim, T.I.; Choi, E.J.; Chung, C.P.; Han, S.B.; Ku, Y. Antimicrobial effect of Zea mays L. and Magnoliae cortex extract mixtures on periodontal pathogen and effect on human gingival fibroblast cellular activity. J. Korean Acad. Periodontol. 2002, 32, 249–255. [Google Scholar] [CrossRef]
- Jeong, P.H.; Kim, Y.S.; Shin, D.H. Changes of physicochemical characteristics of during postharvest ripening at various temperatures. Korean J. Food Sci. Technol. 2006, 38, 469–474. [Google Scholar]
- Nam, H.H.; Kim, H.J.; Choi, N.J.; Roh, S.S.; Choo, B.K. A comparison of antioxidant activity from Schisandra chinensis water extracts depending on stir-frying and stir-frying with liquids process. Korean J. Org. Agric. 2015, 23, 987–997. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, M.J.; Choi, S.K.; Bae, S.H.; An, S.K.; Yoon, Y.M. Antioxidant and antimicrobial effects of lemon and eucalyptus essential oils against skin floras. J. Soc. Cosmet. Sci. Korean 2011, 37, 303–308. [Google Scholar]
- Jeong, J.H.; Kim, S.H.; Huh, C.K. Separation and identification of an antimicrobial substance from Schisandra chinensis extract against Streptococcus mutans KCCM 40105 Strain. Molecules 2023, 28, 3417. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Z.; Cooper, S.L. Interactions between dendrimer biocides and bacterial membranes. Biomaterials 2002, 23, 3359–3368. [Google Scholar] [CrossRef]
- Xing, K.; Chen, X.-G.; Kong, M.; Liu, C.-S.; Cha, D.-S.; Park, H.-J. Effect of oleoyl–chitosan nanoparticles as a novel antibacterial dispersion system on viability, membrane permeability and cell morphology of Escherichia coli and Staphylococcus aureus. Carbohyd. Polym. 2009, 76, 17–22. [Google Scholar] [CrossRef]
- Bensassi, F.; Gallerne, C.; Dein OS, E.; Hajlaoui, M.R.; Bacha, H.; Lemaire, C. Mechanism of alternariol monomethyl ether-induced mitochondrial apoptosis in human colon carcinoma cells. Toxicology 2011, 290, 231–241. [Google Scholar] [CrossRef]
- Xue, Y.-B.; Yang, M.-G.; Li, S.-H.; Li, Z.-J.; Liu, H.-H.; Guo, Q.-B.; Wang, C.-L. The antibiotic activity and mechanisms of active metabolites (Streptomyces alboflavus TD–1) against Ralstonia solanacearum. Biotechnol. Lett. 2019, 41, 1213–1222. [Google Scholar] [CrossRef]
- Wang, B.C.; Guo, Y.S.; Chen, X.T.; Ma, J.L.; Lei, X.; Wang, W.Z.; Long, Y.H. Assessment of the Biocontrol Potential of Bacillus velezensis WL–23 against Kiwifruit Canker Caused by Pseudomonas syringae pv. actinidiae. Int. J. Mol. Sci. 2023, 24, 11541. [Google Scholar] [CrossRef]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef] [PubMed]
- Atiencia-Carrera, M.B.; Cabezas-Mera, F.S.; Tejera, E.; Machado, A. Prevalence of biofilms in Candida spp. bloodstream infections: A meta-analysis. PLoS ONE 2022, 17, e0263522. [Google Scholar] [CrossRef]
- Marsh, P.D.; Head, D.A.; Devine, D.A. Dental plaque as a biofilm and a microbial community—Implications for treatment. BMC Oral Health 2015, 57, 185–191. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogen | Type of Culture Medium |
|---|---|
| Staphylococcus aureus | LB Broth |
| Escherichia coli | LB Broth |
| Vibrio parahaemolyticus | LB Broth |
| ESBL E. coli | LB Broth |
| Pseudomonas aeruginosa | LB Broth |
| Salmonella typhimurium | LB Broth |
| Klebsiella pneumoniae | Nutrient Broth |
| Acinetobacter baumannii | Brian Heart Infusion |
| Levels | Factors | |||
|---|---|---|---|---|
| A Moisture Content (%) | B Time (d) | C Temperature (°C) | D pH | |
| −1 | 100 | 2 | 28 | 4.0 |
| 0 | 150 | 2.5 | 30 | 5.0 |
| 1 | 200 | 3 | 32 | 6.0 |
| Strain Number | Inhibition Zone Diameter (mm) | Strain Number | Inhibition Zone Diameter (mm) |
|---|---|---|---|
| S10 | 18.69 ± 0.82 | S1 | 17.02 ± 0.79 |
| W1 | 15.57 ± 1.67 | S2 | 14.73 ± 0.81 |
| W2 | 17.33 ± 0.46 | W8 | 16.73 ± 0.24 |
| W4 | 17.89 ± 1.12 | W7 | 13.62 ± 0.18 |
| Pathogen | Inhibition Zone Diameter (mm) | Pathogen | Inhibition Zone Diameter (mm) |
|---|---|---|---|
| Staphylococcus aureus | 14.15 ± 0.29 | ESBL E. coli | 17.55 ± 1.22 |
| Acinetobacter baumannii | 13.35 ± 0.98 | Escherichia coli | 13.93 ± 0.42 |
| Vibrio parahaemolyticus | 16.64 ± 0.78 | Klebsiella pneumoniae | 17.37 ± 0.32 |
| Pseudomonas aeruginosa | 16.85 ± 0.19 | Salmonella typhimurium | 18.43 ± 0.92 |
| Concentration of Crude Extract (μg/mL) | OD600 | Concentration of Crude Extract (μg/mL) | OD600 |
|---|---|---|---|
| 50.00 | 0.030 ± 0.011 | 1.56 | 0.371 ± 0.031 |
| 25.00 | 0.080 ± 0.007 | 0.78 | 0.482 ± 0.078 |
| 12.5 | 0.122 ± 0.019 | Negative control | 0.047 ± 0.003 |
| 6.25 | 0.165 ± 0.009 | Positive control | 0.827 ± 0.011 |
| Concentration of Crude Extract (μg/mL) | OD590 | Inhibition Rate (%) |
|---|---|---|
| 0 (Control) | 1.233 ± 0.041 | - |
| 1/2 MIC | 1.043 ± 0.038 | 15.4% |
| 1 MIC | 0.694 ± 0.052 | 42.7% |
| 2 MIC | 0.635 ± 0.047 | 48.4% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Shi, Y.; Ma, W.; Cai, S.; Yang, X.; Xu, L.; Hou, X.; Wang, L.; Jin, L.; Quan, C. Optimization of Fermentation Conditions for Endophytic Fungi from Schisandra chinensis and Investigation of Their Antibacterial Mechanisms Against Methicillin-Resistant Staphylococcus aureus. Microorganisms 2025, 13, 982. https://doi.org/10.3390/microorganisms13050982
Li M, Shi Y, Ma W, Cai S, Yang X, Xu L, Hou X, Wang L, Jin L, Quan C. Optimization of Fermentation Conditions for Endophytic Fungi from Schisandra chinensis and Investigation of Their Antibacterial Mechanisms Against Methicillin-Resistant Staphylococcus aureus. Microorganisms. 2025; 13(5):982. https://doi.org/10.3390/microorganisms13050982
Chicago/Turabian StyleLi, Mengyu, Yuewei Shi, Wenwei Ma, Shouyuan Cai, Xinyuan Yang, Lukai Xu, Xiyan Hou, Lulu Wang, Liming Jin, and Chunshan Quan. 2025. "Optimization of Fermentation Conditions for Endophytic Fungi from Schisandra chinensis and Investigation of Their Antibacterial Mechanisms Against Methicillin-Resistant Staphylococcus aureus" Microorganisms 13, no. 5: 982. https://doi.org/10.3390/microorganisms13050982
APA StyleLi, M., Shi, Y., Ma, W., Cai, S., Yang, X., Xu, L., Hou, X., Wang, L., Jin, L., & Quan, C. (2025). Optimization of Fermentation Conditions for Endophytic Fungi from Schisandra chinensis and Investigation of Their Antibacterial Mechanisms Against Methicillin-Resistant Staphylococcus aureus. Microorganisms, 13(5), 982. https://doi.org/10.3390/microorganisms13050982