Author Contributions
Conceptualization, W.L.S. and S.S.G.; methodology, C.R.Z., G.V.S.Z., E.C., L.M.M. and G.D.B.d.S.; investigation, C.R.Z., E.C., L.M.M. and G.D.B.d.S.; resources, E.S.K., W.L.S. and S.S.G.; data curation, C.R.Z.; writing—original draft preparation, C.R.Z.; writing—review and editing, F.H.d.F., M.d.M.F.P., E.S.K., W.L.S., S.S.G. and C.R.Z.; supervision, W.L.S. and S.S.G.; project administration, W.L.S. and S.S.G. All authors have read and agreed to the published version of the manuscript.
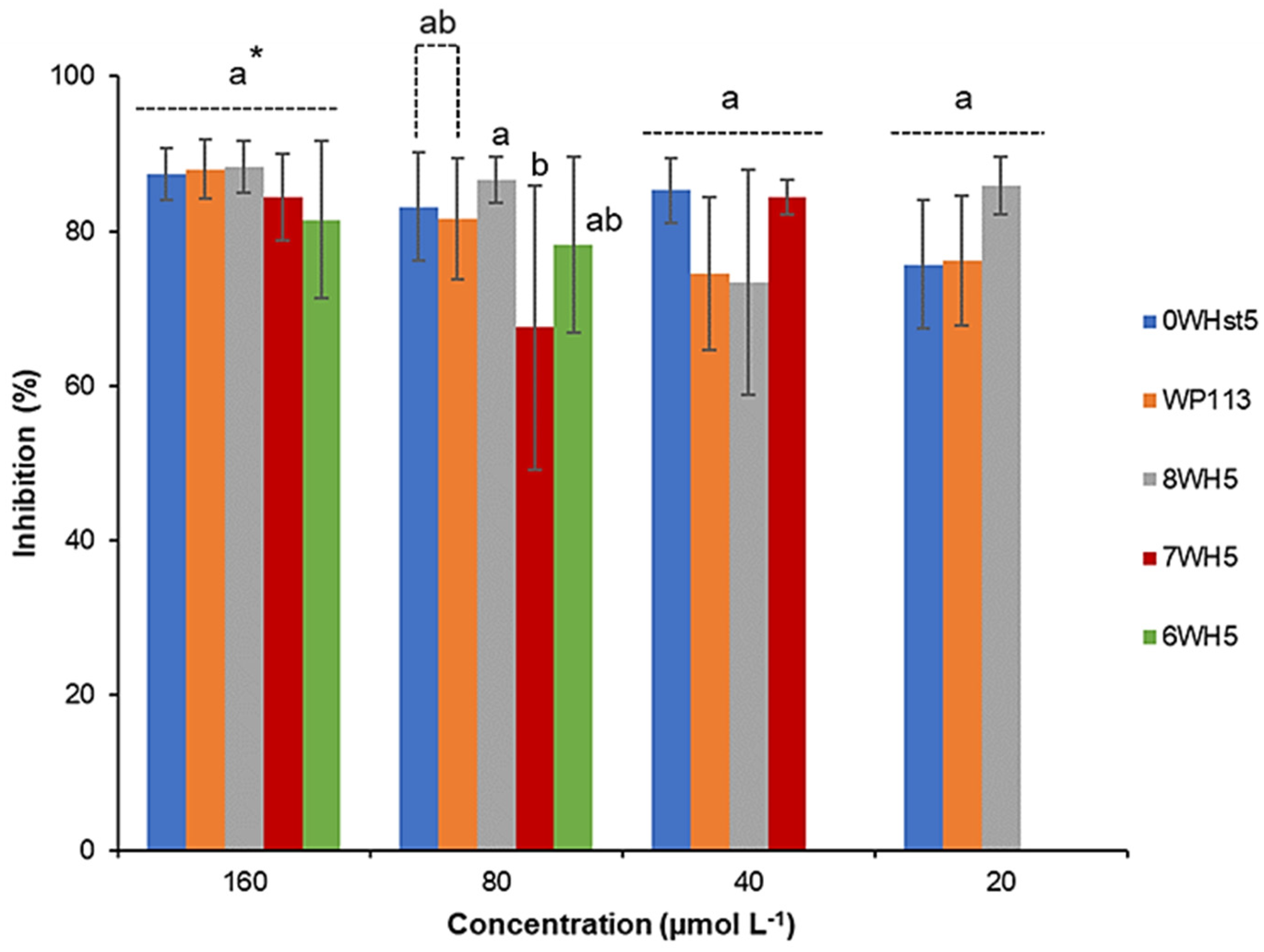
Figure 1.
Inhibition of C. albicans ATCC 90028 by 0WHst5, WP113, 8WH5, 7WH5 and 6WH5. The experiment was performed in triplicate. * Peptides were statistically compared between treatments of the same concentration. Equal letters indicate statistically equal means while different letters indicate statistically different means.
Figure 1.
Inhibition of C. albicans ATCC 90028 by 0WHst5, WP113, 8WH5, 7WH5 and 6WH5. The experiment was performed in triplicate. * Peptides were statistically compared between treatments of the same concentration. Equal letters indicate statistically equal means while different letters indicate statistically different means.
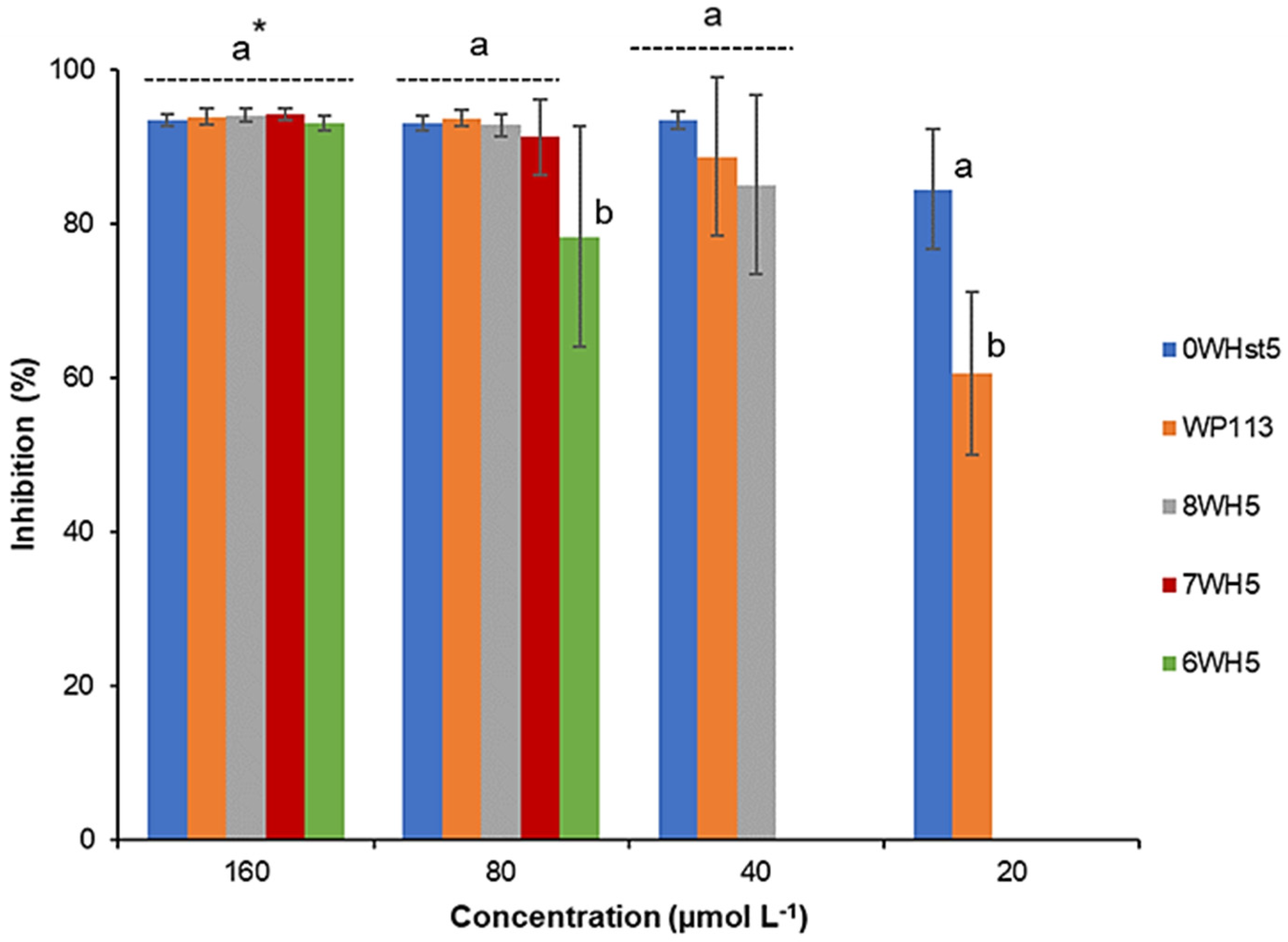
Figure 2.
Inhibition of C. albicans ATCC 18801 by 0WHst5, WP113, 8WH5, 7WH5 and 6WH5. The experiment was performed in triplicate. * Peptides were statistically compared between treatments of the same concentration. Equal letters indicate statistically equal means while different letters indicate statistically different means.
Figure 2.
Inhibition of C. albicans ATCC 18801 by 0WHst5, WP113, 8WH5, 7WH5 and 6WH5. The experiment was performed in triplicate. * Peptides were statistically compared between treatments of the same concentration. Equal letters indicate statistically equal means while different letters indicate statistically different means.
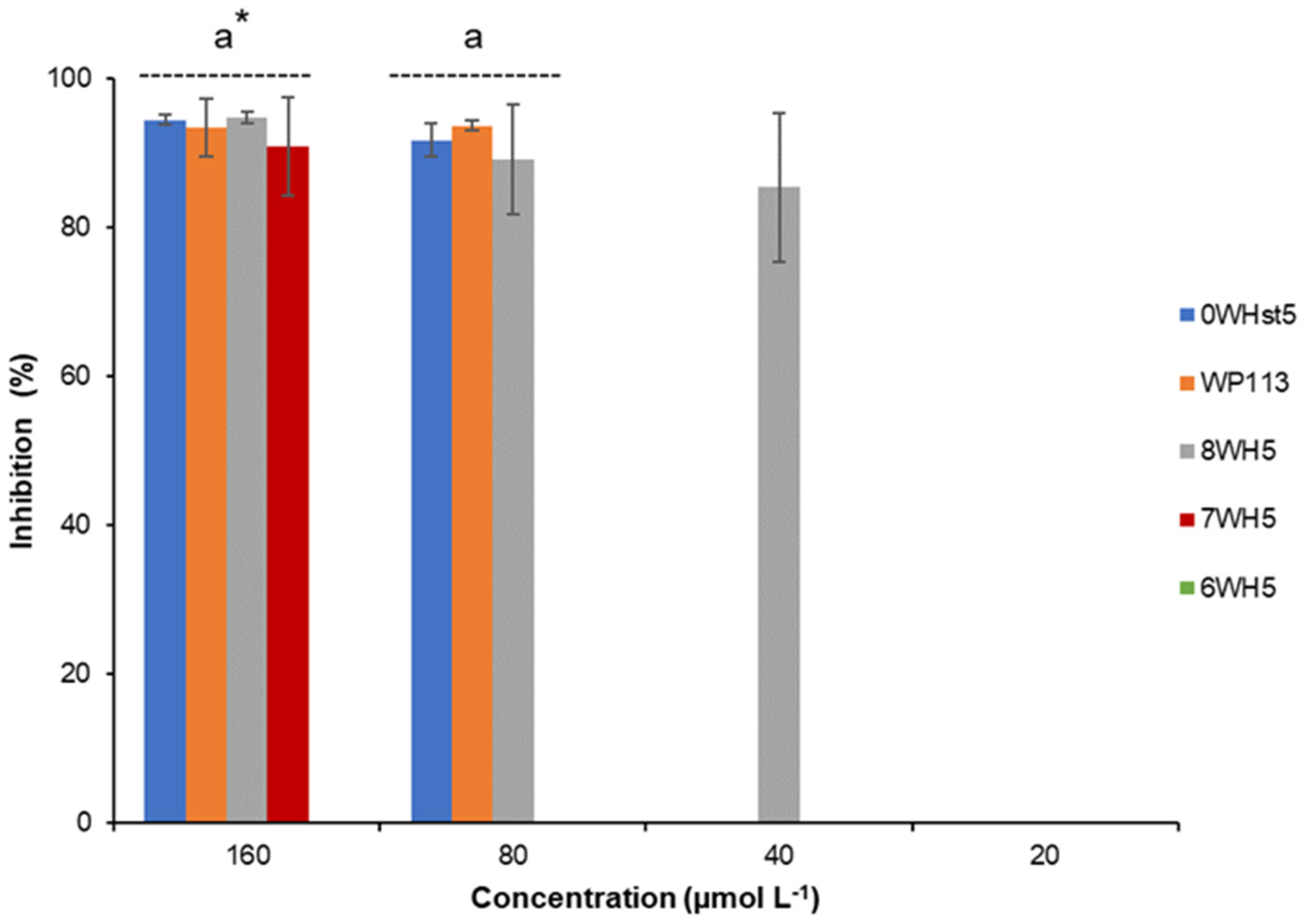
Figure 3.
Inhibition of C. albicans ATCC 10231 by 0WHst5, WP113, 8WH5, 7WH5 and 6WH5. The experiment was performed in triplicate. * Peptides were statistically compared between treatments of the same concentration. Equal letters indicate statistically equal means while different letters indicate statistically different means.
Figure 3.
Inhibition of C. albicans ATCC 10231 by 0WHst5, WP113, 8WH5, 7WH5 and 6WH5. The experiment was performed in triplicate. * Peptides were statistically compared between treatments of the same concentration. Equal letters indicate statistically equal means while different letters indicate statistically different means.
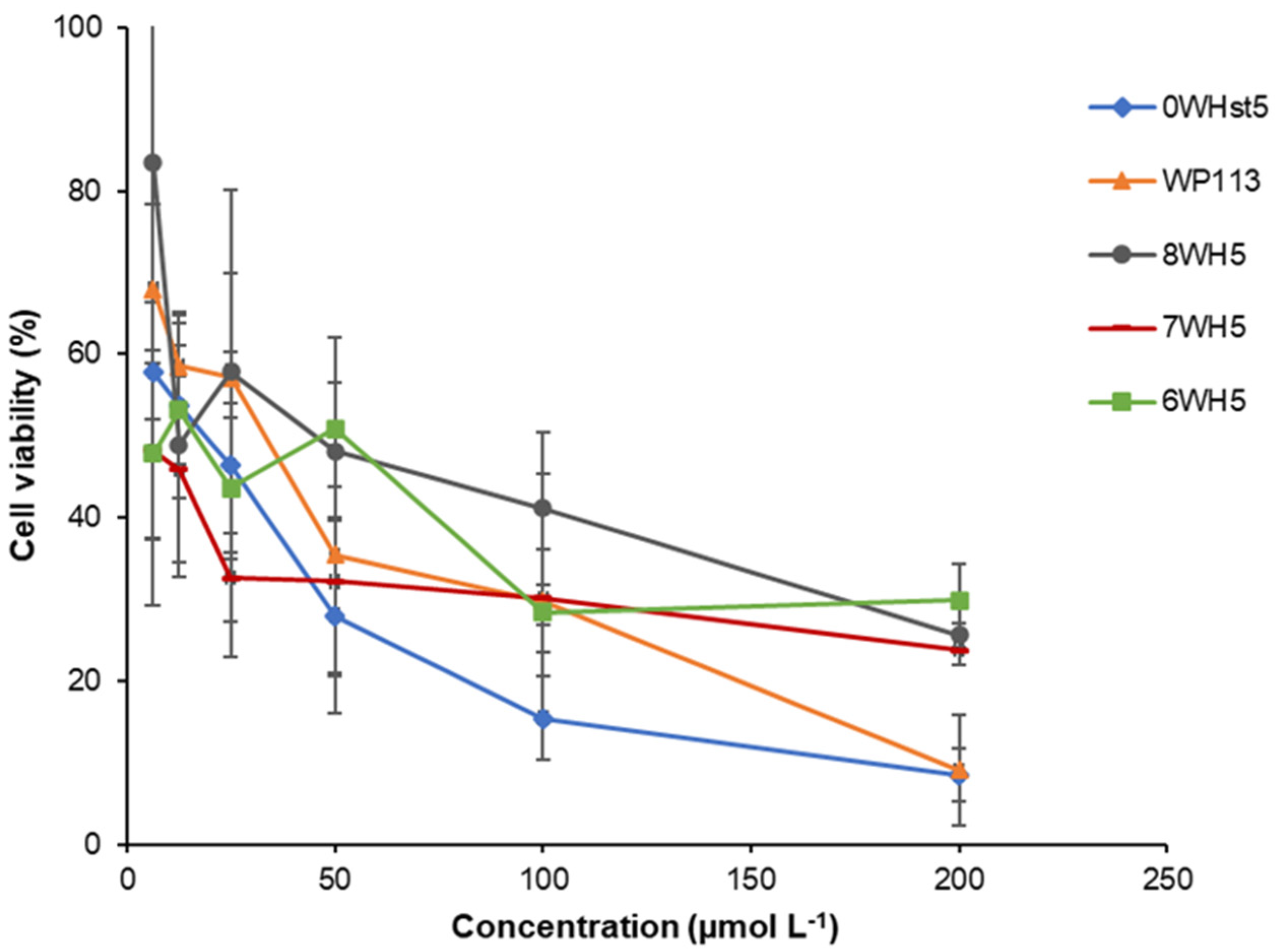
Figure 4.
Cell viability of C. albicans ATCC 90028 cells after 2 h incubation at 37 °C with dilution series of peptides 0WHst5, WP113, W8H5, 7WH5 and 6WH5. The dilutions were then plated in Sabouraud Dextrose Agar (SDA) media and the logarithm values of CFU mL−1 were calculated for the determination of cell viability.
Figure 4.
Cell viability of C. albicans ATCC 90028 cells after 2 h incubation at 37 °C with dilution series of peptides 0WHst5, WP113, W8H5, 7WH5 and 6WH5. The dilutions were then plated in Sabouraud Dextrose Agar (SDA) media and the logarithm values of CFU mL−1 were calculated for the determination of cell viability.
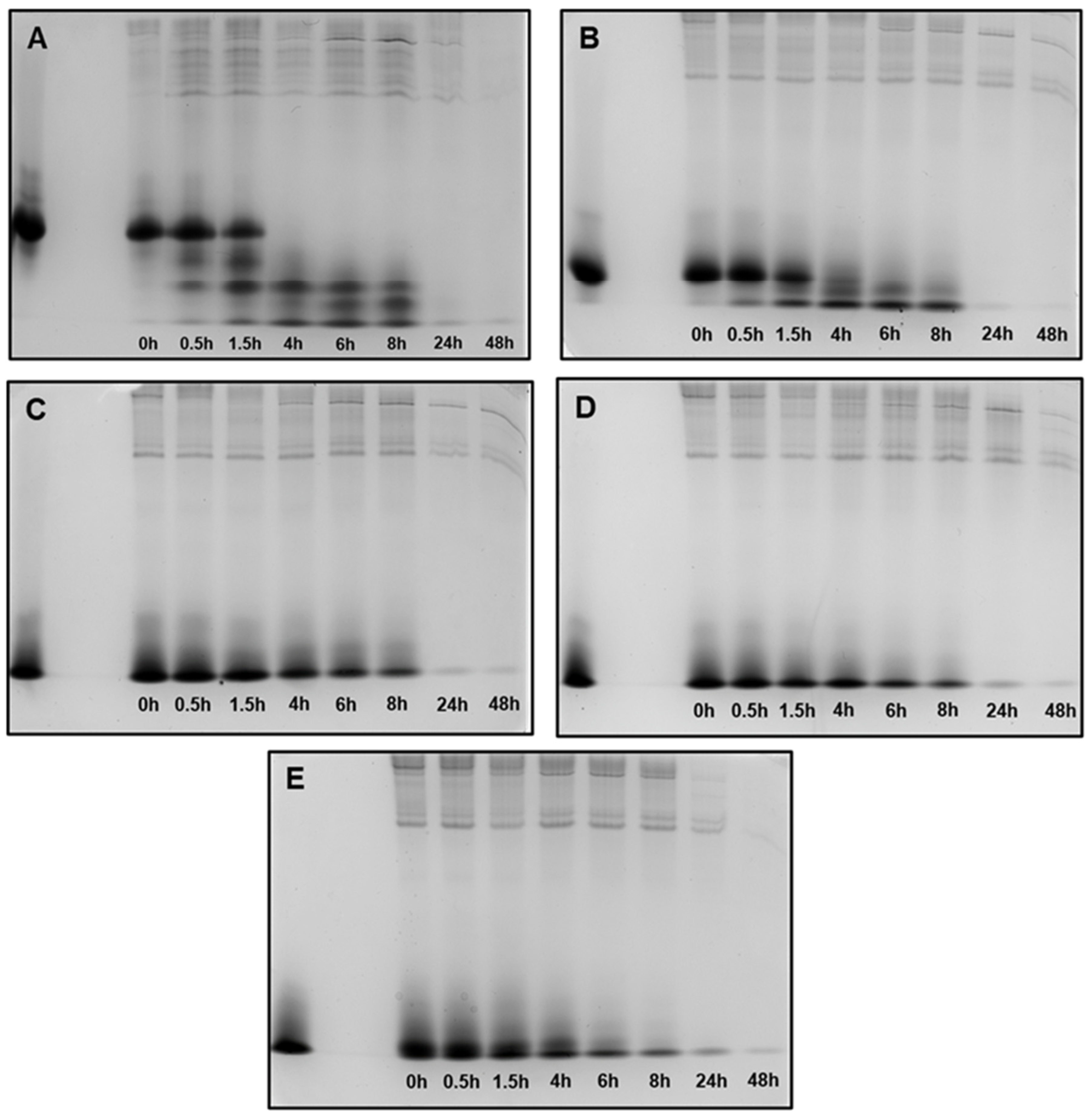
Figure 5.
Peptide degradation profile in human saliva (WSS): (A) 0WHst5; (B) WP113; (C) 8WH5; (D) 7WH5; (E) 6WH5. An aliquot of 100 µL was collected after each incubation period and evaluated by cationic-PAGE. The first column on the left (column 1) corresponds to the standard band for all peptides tested; columns 2 to 9 show the different incubation periods (t = 0, 0.5, 1.5, 4, 6, 8, 24 and 48 h).
Figure 5.
Peptide degradation profile in human saliva (WSS): (A) 0WHst5; (B) WP113; (C) 8WH5; (D) 7WH5; (E) 6WH5. An aliquot of 100 µL was collected after each incubation period and evaluated by cationic-PAGE. The first column on the left (column 1) corresponds to the standard band for all peptides tested; columns 2 to 9 show the different incubation periods (t = 0, 0.5, 1.5, 4, 6, 8, 24 and 48 h).
Figure 6.
Peptide degradation over time for 0WHst5, WP113, 8WH5, 7WH5 and 6WH5. Data were obtained after the cationic-PAGE and the amount of peptide in each aliquot was determined using the software Image Lab 6.1 (Bio-Rad Inc.). Error bars correspond to the standard deviations on duplicate samples.
Figure 6.
Peptide degradation over time for 0WHst5, WP113, 8WH5, 7WH5 and 6WH5. Data were obtained after the cationic-PAGE and the amount of peptide in each aliquot was determined using the software Image Lab 6.1 (Bio-Rad Inc.). Error bars correspond to the standard deviations on duplicate samples.
Figure 7.
Proteolytic cleavage sites in the peptides target to salivary proteases. The stars indicate all primary cleavage sites and the circle indicates preferred primary cleavage sites.
Figure 7.
Proteolytic cleavage sites in the peptides target to salivary proteases. The stars indicate all primary cleavage sites and the circle indicates preferred primary cleavage sites.
Figure 8.
0WHst5 and WP113 degradation profile in human saliva. Aliquots of 100 µL were boiled and analyzed by HPLC after 0 h, 1.5 h and 8 h of incubation. Black arrows indicate intact 0WHst5 and WP113, retention time in 15.35 and 15.02 min, respectively.
Figure 8.
0WHst5 and WP113 degradation profile in human saliva. Aliquots of 100 µL were boiled and analyzed by HPLC after 0 h, 1.5 h and 8 h of incubation. Black arrows indicate intact 0WHst5 and WP113, retention time in 15.35 and 15.02 min, respectively.
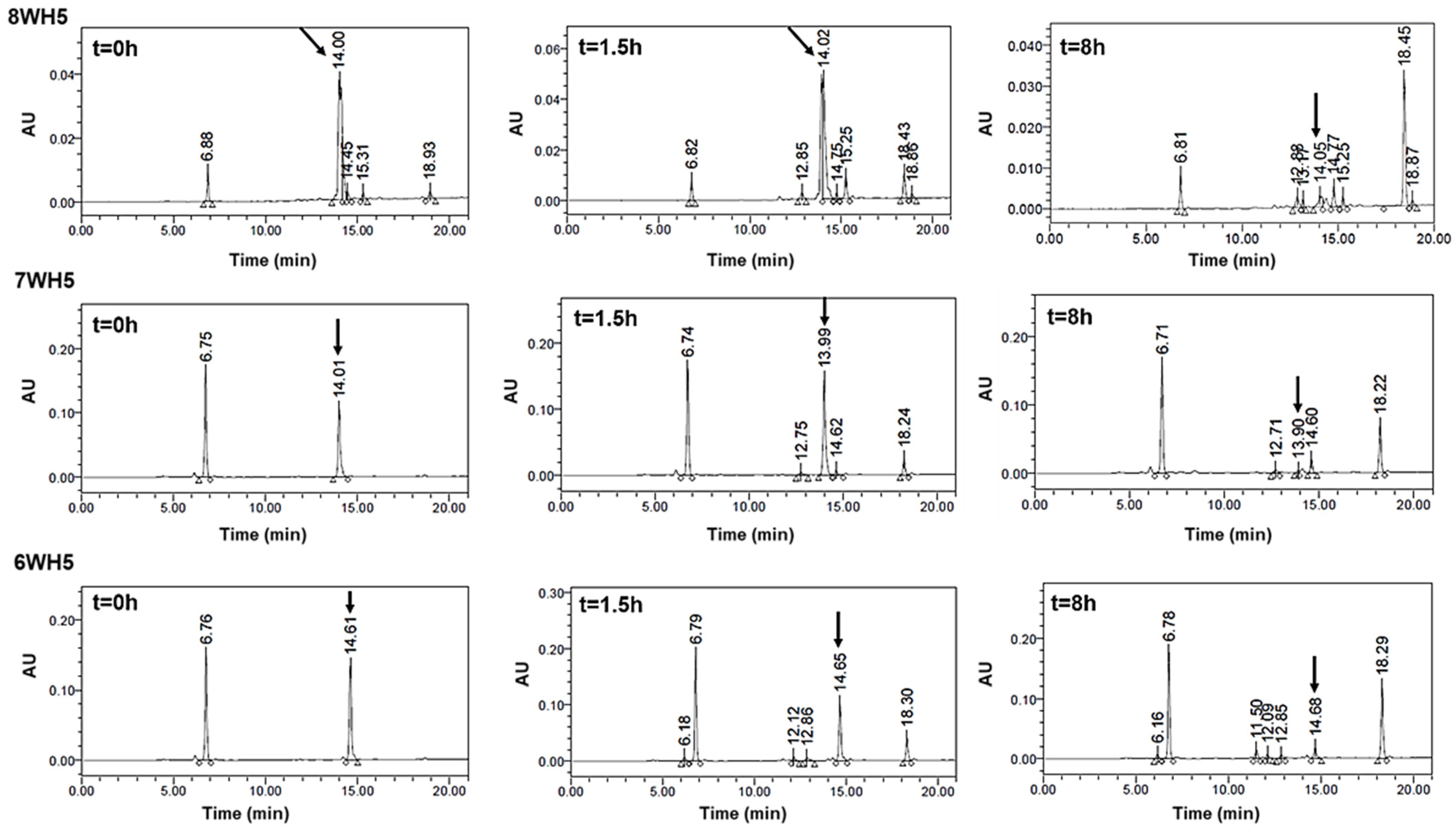
Figure 9.
8WH5, 7WH5 and 6WH5 degradation profile in human saliva. Aliquots of 100 µL were boiled and analyzed by HPLC after 0 h, 1.5 h and 8 h of incubation. Black arrows indicate intact 8WH5, 7WH5 and 6WH5, retention time at 14.00, 13.99 and 14.61 min, respectively.
Figure 9.
8WH5, 7WH5 and 6WH5 degradation profile in human saliva. Aliquots of 100 µL were boiled and analyzed by HPLC after 0 h, 1.5 h and 8 h of incubation. Black arrows indicate intact 8WH5, 7WH5 and 6WH5, retention time at 14.00, 13.99 and 14.61 min, respectively.
Table 1.
Amino acid sequence and physicochemical features of the synthetic peptides.
Table 1.
Amino acid sequence and physicochemical features of the synthetic peptides.
| Peptide | Amino Acid Sequence | Net Charge * | MW (g mol−1) |
|---|
| 0WHst5 | W D S H A K R H H G Y K R K F H E K H H S H R G Y | +5.7 | 3225.50 |
| WP113 | W A K R H H G Y K R K F H | +5.3 | 1751.01 |
| 8WH5 | W K R H H G Y K R | +4.2 | 1267.44 |
| 7WH5 | W K R H H G Y K | +3.2 | 1111.26 |
| 6WH5 | W K R H H G Y | +2.2 | 983.09 |
Table 2.
Quantification bands for the peptides 0WHst5, WP113, 8WH5, 7WH5 and 6WH5 remaining after withdrawal of aliquots at times 0, 0.5, 1.5, 4, 6, 8, 24 and 48 h. Bands were analyzed and quantified using Image Lab 6.1 software (Bio-Rad Inc., Hercules, CA, USA).
Table 2.
Quantification bands for the peptides 0WHst5, WP113, 8WH5, 7WH5 and 6WH5 remaining after withdrawal of aliquots at times 0, 0.5, 1.5, 4, 6, 8, 24 and 48 h. Bands were analyzed and quantified using Image Lab 6.1 software (Bio-Rad Inc., Hercules, CA, USA).
| Time (h) | 0WHst5 (%) | WP113 (%) | 8WH5 (%) | 7WH5 (%) | 6WH5 (%) |
|---|
| 0 | 100.0 ± 0.0 | 100.0 ± 0.0 | 100.0 ± 0.0 | 100.0 ± 0.0 | 100.0 ± 0.0 |
| 0.5 | 91.0 ± 17.0 | 89.5 ± 7.3 | 99.9 ± 8.0 | 100.0 ± 4.5 | 98.8 ± 1.1 |
| 1.5 | 61.0 ± 19.8 | 56.3 ± 30.9 | 78.2 ± 13.9 | 76.8 ± 15.7 | 96.6 ± 4.6 |
| 4 | - | 7.23 ± 8.17 | 80.3 ± 13.0 | 86.2 ± 3.1 | 79.3 ± 9.8 |
| 6 | - | - | 62.0 ± 8.4 | 55.9 ± 5.8 | 52.4 ± 7.0 |
| 8 | - | - | 44.2 ± 0.9 | 49.6 ± 2.4 | 30.6 ± 4.0 |
| 24 | - | - | 8.6 ± 4.4 | 19.1 ± 7.5 | 16.5 ± 5.9 |
| 48 | - | - | 3.5 ± 0.8 | 6.9 ± 1.0 | 8.3 ± 6.2 |
Table 3.
Amino acid sequences and properties of WHst5 fragments generated upon 48 h of incubation with diluted WSS.
Table 3.
Amino acid sequences and properties of WHst5 fragments generated upon 48 h of incubation with diluted WSS.
| 0WHst5 |
|---|
| Name | Observed (M + H)+ | Theoretical (M + H)+ | Sequence |
|---|
| Histatin 5 | - | 3036.29 | D S H A K R H H G Y K R K F H E K H H S H R G Y ** |
|---|
| his5 1/14 | 1766.93 | 1766.96 | D S H A K R H H G Y K R K F * |
| his5 7/24 | 2341.17 | 2341.55 | H H G Y K R K F H E K H H S H R G Y * |
| his5 8/24 | 2204.11 | 2204.41 | H G Y K R K F H E K H H S H R G Y * |
| his5 9/24 | 2067.05 | 2067.28 | G Y K R K F H E K H H S H R G Y * |
| his5 10/24 | 2010.03 | 2010.22 | Y K R K F H E K H H S H R G * |
| his5 11/24 | 1846.96 | 1847.05 | K R K F H E K H H S H R G Y |
| his5 12/24 | 1718.87 | 1718.88 | R K F H E K H H S H R G Y |
| his5 13/24 | 1562.77 | 1562.69 | K F H E K H H S H R G Y |
| his5 13/23 | 1399.70 | 1399.52 | K F H E K H H S H R G |
| his5 14/24 | 1434.67 | 1434.52 | F H E K H H S H R G Y |
Table 4.
Amino acid sequences and properties of WP113 fragments generated upon 48 h of incubation with diluted WSS.
Table 4.
Amino acid sequences and properties of WP113 fragments generated upon 48 h of incubation with diluted WSS.
| WP113 |
|---|
| Name | Observed (M + H)+ | Theoretical (M + H)+ | Sequence |
|---|
| P113 | - | 1564.8 | A K R H H G Y K R K F H ** |
|---|
| his5 1/21 | 2659.33 | 2659.88 | D S H A K R H H G Y K R K F H E K H H S H * |
| his5 1/15 | 1903.98 | 1904.13 | D S H A K R H H G Y K R K F H * |
| his5 1/14 | 1766.93 | 1766.96 | D S H A K R H H G Y K R K F * |
| his5 1/8 | 987.48 | 987.03 | D S H A K R H H |
| his5 2/15 | 1788.96 | 1789.02 | S H A K R H H G Y K R K F H * |
| his5 2/14 | 1651.90 | 1651.90 | S H A K R H H G Y K R K F * |
| his5 3/15 | 1701.93 | 1701.94 | H A K R H H G Y K R K F H * |
| his5 3/15 | 1564.87 | 1564.82 | H A K R H H G Y K R K F * |
| his5 3/8 | 785.42 | 784.87 | H A K R H H |
| his5 4/18 | 1959.06 | 1959.22 | A K R H H G Y K R K F H E K H * |
| his5 4/15 | 1564.87 | 1564.82 | A K R H H G Y K R K F H * |
| his5 4/14 | 1427.81 | 1427.66 | A K R H H G Y K R K F * |
| his5 5/24 | 2625.36 | 2625.91 | K R H H G Y K R K F H E K H H S H R G Y * |
| his5 5/15 | 1493.83 | 1493.72 | K R H H G Y K R K F H * |
| his5 5/13 | 1209.70 | 1209.41 | K R H H G Y K R K * |
| his5 6/21 | 2121.08 | 2121.33 | R H H G Y K R K F H E K H H S H * |
| his5 6/18 | 1759.93 | 1759.97 | R H H G Y K R K F H E K H * |
| his5 6/17 | 1622.87 | 1622.83 | R H H G Y K R K F H E K * |
| his5 6/15 | 1365.73 | 1365.55 | R H H G Y K R K F H * |
| his5 6/14 | 1228.68 | 1228.41 | R H H G Y K R K F * |
| his5 6/13 | 1081.61 | 1081.24 | R H H G Y K R K * |
| his5 7/18 | 1603.83 | 1603.79 | H H G Y K R K F H E K H * |
| his5 7/15 | 1209.63 | 1209.36 | H H G Y K R K F H * |
| his5 7/14 | 1072.57 | 1072.22 | H H G Y K R K F |
| his5 8/19 | 1603.83 | 1603.79 | H G Y K R K F H E K H H * |
| his5 8/15 | 1072.57 | 1072.22 | H G Y K R K F H * |
| his5 9/19 | 1466.77 | 1466.65 | G Y K R K F H E K H H * |
| his5 9/15 | 935.52 | 935.08 | G Y K R K F H * |
| his5 9/12 | 523.29 | 522.60 | G Y K R |
| his5 10/24 | 2010.03 | 2010.22 | Y K R K F H E K H H S H R G Y |
| his5 11/24 | 1846.96 | 1847.05 | K R K F H E K H H S H R G Y |
| his5 12/24 | 1718.87 | 1718.88 | R K F H E K H H S H R G Y |
| his5 12/22 | 1498.78 | 1498.65 | R K F H E K H H S H R |
| his5 13/24 | 1562.77 | 1562.69 | K F H E K H H S H R G Y |
| his5 14/24 | 1434.67 | 1434.52 | F H E K H H S H R G Y |
| his5 14/18 | 697.34 | 696.76 | F H E K H |
| his5 15/21 | 911.4 | 910.94 | H E K H H S H |
Table 5.
Amino acid sequences and properties of 8WH5 fragments generated upon 48 h of incubation with diluted WSS.
Table 5.
Amino acid sequences and properties of 8WH5 fragments generated upon 48 h of incubation with diluted WSS.
| 8WH5 |
|---|
| Name | Observed (M + H)+ | Theoretical (M + H)+ | Sequence |
|---|
| 8WH5 | - | 1081.24 | K R H H G Y K R ** |
|---|
| his5 1/12 | 1491.76 | 1491.62 | D S H A K R H H G Y K R * |
| his5 3/12 | 1289.70 | 1289.45 | H A K R H H G Y K R * |
| his5 4/7 | 511.30 | 510.59 | A K R H |
| his5 5/13 | 1209.70 | 1209.41 | K R H H G Y K R K * |
| his5 5/12 | 1081.61 | 1081.24 | K R H H G Y K R * |
| his5 6/19 | 1896.99 | 1897.11 | R H H G Y K R K F H E K H H |
Table 6.
Amino acid sequences and properties of 7WH5 fragments generated upon 48 h of incubation with diluted WSS.
Table 6.
Amino acid sequences and properties of 7WH5 fragments generated upon 48 h of incubation with diluted WSS.
| 7WH5 |
|---|
| Name | Observed (M + H)+ | Theoretical (M + H)+ | Sequence |
|---|
| 7WH5 | - | 925.05 | K R H H G Y K ** |
|---|
| his5 1/14 | 1766.93 | 1766.99 | D S H A K R H H G Y K R K F * |
| his5 5/13 | 1209.70 | 1209.42 | K R H H G Y K R K * |
| his5 5/11 | 925.51 | 925.06 | K R H H G Y K * |
| his5 6/11 | 797.41 | 796.88 | R H H G Y K * |
| his5 7/13 | 925.51 | 925.05 | H H G Y K R K |
| his5 12/17 | 844.47 | 843.97 | R K F H E K |
Table 7.
Amino acid sequences and properties of 6WH5 fragments generated upon 48 h of incubation with diluted WSS.
Table 7.
Amino acid sequences and properties of 6WH5 fragments generated upon 48 h of incubation with diluted WSS.
| 6WH5 |
|---|
| Name | Observed (M + H)+ | Theoretical (M + H)+ | Sequence |
|---|
| 6WH5 | - | 796.88 | K R H H G Y ** |
|---|
| his5 5/10 | 797.41 | 796.88 | K R H H G Y * |
| his5 10/24 | 2010.03 | 2010.22 | Y K R K F H E K H H S H R G Y |

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}