

Effects of Dietary Clostridium butyricum on Growth and Intestinal Mucosal Barrier Functions of Juvenile Channel Catfish (Ictalurus punctatus)

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Feeding Trial and Sample Collection
2.2. Intestinal Digestive and Antioxidant Enzyme Activities
2.3. Intestinal Mucosal Barrier Functions-Related Gene Expression
2.4. Extraction of Gut Microbial Genomic DNA and 16S rRNA Gene Amplicon Sequencing
2.5. Analysis of 16S rRNA Gene Amplicon Sequencing Data
2.6. Intestinal LC-MS Untargeted Metabolomics
2.7. Statistical Analysis
3. Results
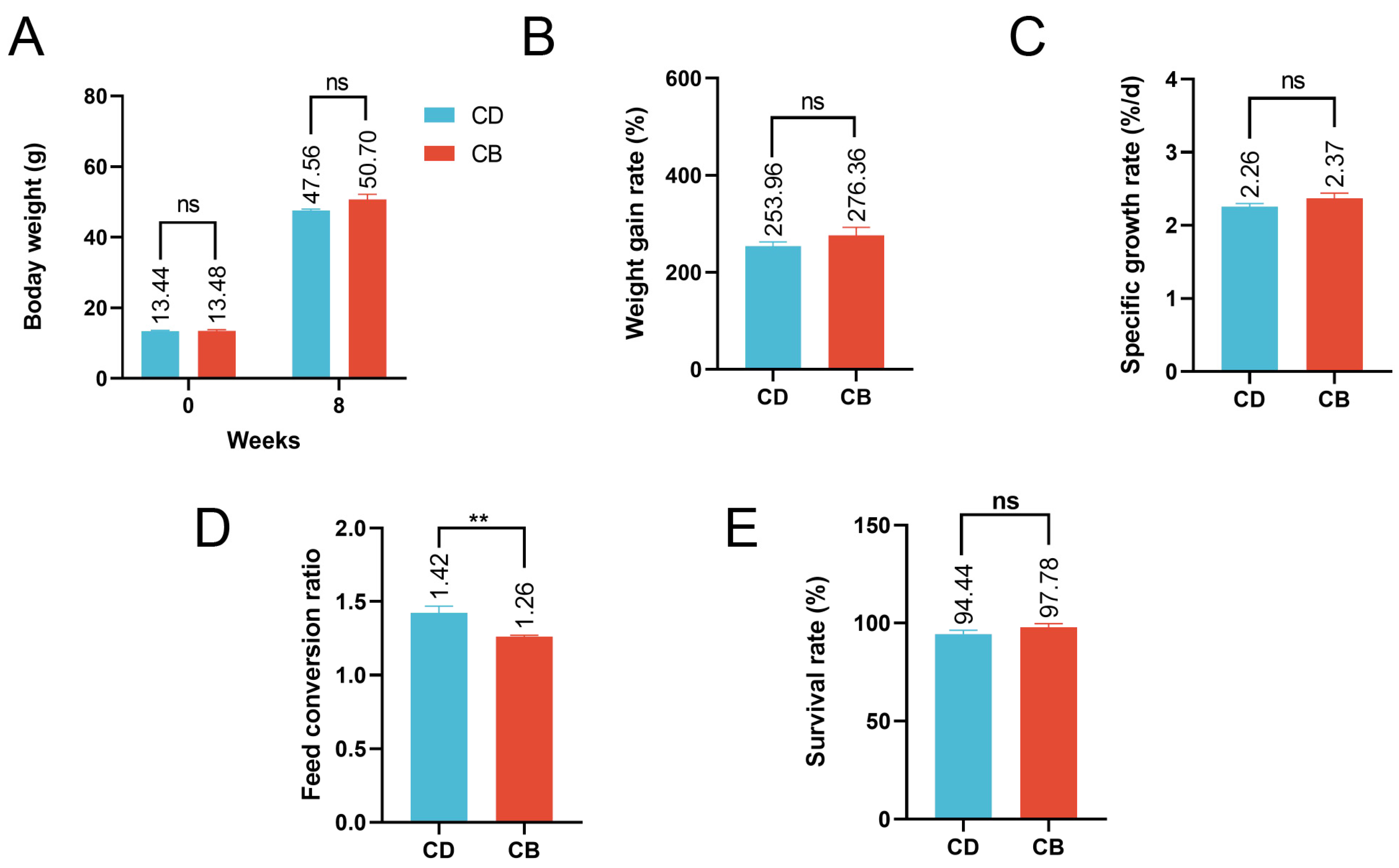
3.1. Growth Performance
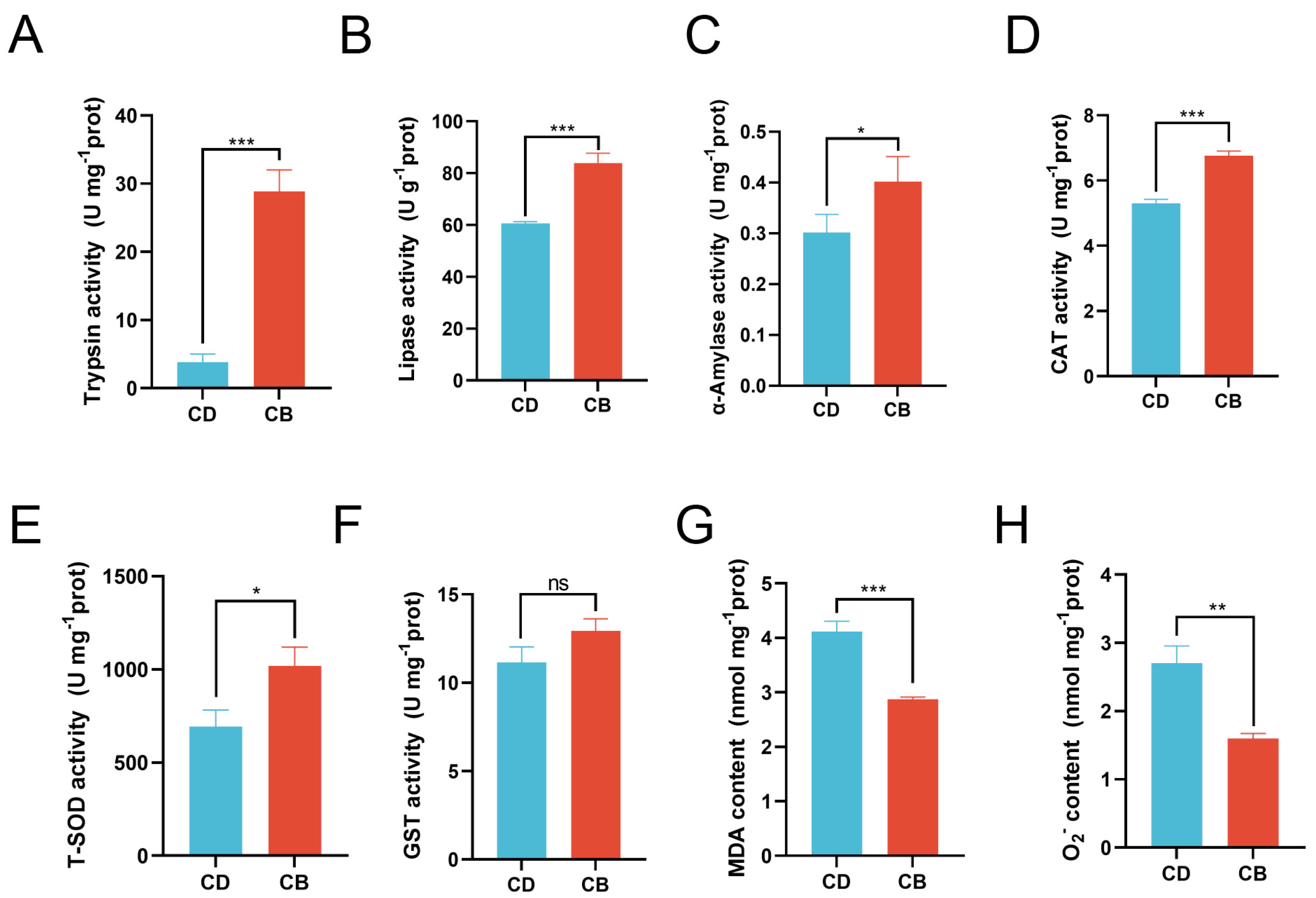
3.2. Intestinal Digestion and Antioxidant Capacity
3.3. Gene Expression Associated with Intestinal Mucosal Barrier Functions
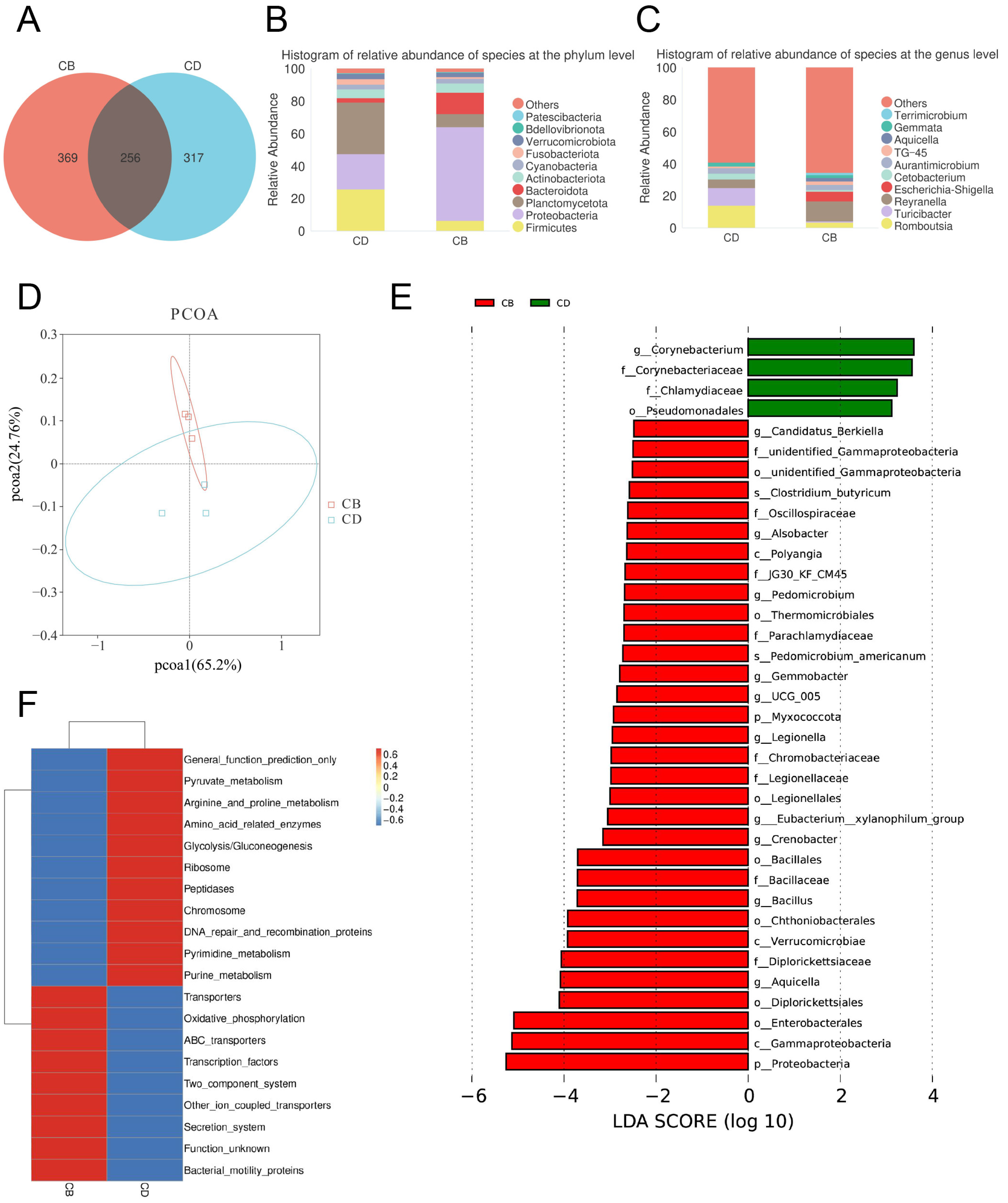
3.4. Intestinal Microbiota
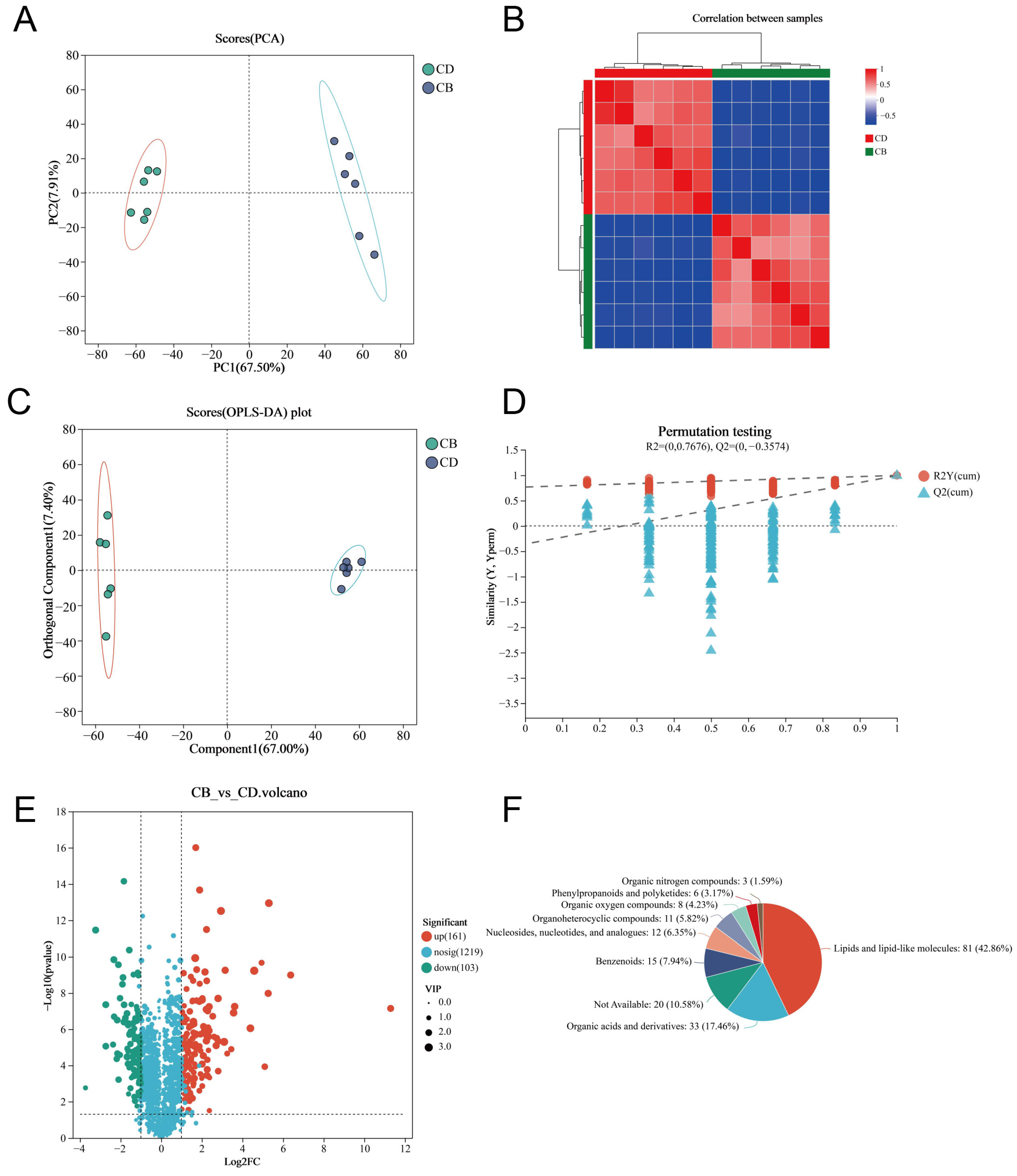
3.5. LC-MS Untargeted Metabolomics of the Intestine
4. Discussion
4.1. Dietary C. butyricum Enhanced the Growth Performance and Intestinal Digestive Enzyme Activities of Juvenile Channel Catfish
4.2. Dietary C. butyricum Elevated Intestinal Antioxidant Enzyme Activities of Juvenile Channel Catfish
4.3. Dietary C. butyricum Increased the Expression of Genes Linked to Intestinal Mucosal Barrier Functions of Juvenile Channel Catfish
4.4. Dietary C. butyricum Improved the Gut Microbiota of Juvenile Channel Catfish
4.5. Dietary C. butyricum Improved the Intestinal Metabolic Profile of Juvenile Channel Catfish
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Y.; Xu, G.; Nie, Z.; Shao, N.; Li, Q.; Xu, P. Growth Performance of Bluntnose Black Bream, Channel Catfish, Yellow Catfish, and Largemouth Bass Reared in the in-Pond Raceway Recirculating Culture System. N. Am. J. Aquac. 2019, 81, 153–159. [Google Scholar] [CrossRef]
- Rodger, H.D. Fish Disease Causing Economic Impact in Global Aquaculture. In Fish Vaccines; Adams, A., Ed.; Springer: Basel, Switzerland, 2016; pp. 1–34. [Google Scholar]
- Celi, P.; Verlhac, V.; Calvo, E.P.; Schmeisser, J.; Kluenter, A.-M. Biomarkers of Gastrointestinal Functionality in Animal Nutrition and Health. Anim. Feed Sci. Technol. 2019, 250, 9–31. [Google Scholar] [CrossRef]
- Thoo, L.; Noti, M.; Krebs, P. Keep Calm: The Intestinal Barrier at the Interface of Peace and War. Cell Death Dis. 2019, 10, 849. [Google Scholar] [CrossRef] [PubMed]
- Vancamelbeke, M.; Vermeire, S. The Intestinal Barrier: A Fundamental Role in Health and Disease. Expert. Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef]
- Romero, E.S.; Cotoner, C.A.; Camacho, C.P.; Bedmar, M.C.; Vicario, M. The Intestinal Barrier Function and Its Involvement in Digestive Disease. Rev. Esp. Enferm. Dig. 2015, 107, 686–696. [Google Scholar]
- Nalle, S.C.; Turner, J.R. Intestinal Barrier Loss as a Critical Pathogenic Link between Inflammatory Bowel Disease and Graft-Versus-Host Disease. Mucosal Immunol. 2015, 8, 720–730. [Google Scholar] [CrossRef]
- Yingying, H.U.; Juying, W.; Deyi, M.A. Research Progress on Environmental Effect of Antibiotic Agents in Marine Aquaculture. Mar. Environ. Sci. 2004, 23, 76–80. [Google Scholar]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert Consensus Document. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Shi, S.; Liu, J.; Dong, J.; Hu, J.; Liu, Y.; Feng, J.; Zhou, D. Research Progress on the Regulation Mechanism of Probiotics on the Microecological Flora of Infected Intestines in Livestock and Poultry. Lett. Appl. Microbiol. 2022, 74, 647–655. [Google Scholar] [CrossRef]
- Koga, Y. Microbiota in the Stomach and Application of Probiotics to Gastroduodenal Diseases. World J. Gastroenterol. 2022, 28, 6702–6715. [Google Scholar] [CrossRef]
- Ou, W.; Yu, G.; Zhang, Y.; Mai, K. Recent Progress in the Understanding of the Gut Microbiota of Marine Fishes. Mar. Life Sci. Technol. 2021, 3, 434–448. [Google Scholar] [CrossRef] [PubMed]
- Ariyoshi, T.; Hagihara, M.; Takahashi, M.; Mikamo, H. Effect of Clostridium butyricum on Gastrointestinal Infections. Biomedicines 2022, 10, 483. [Google Scholar] [CrossRef] [PubMed]
- Junghare, M.; Subudhi, S.; Lal, B. Improvement of Hydrogen Production under Decreased Partial Pressure by Newly Isolated Alkaline Tolerant Anaerobe, Clostridium butyricum Tm-9a: Optimization of Process Parameters. Int. J. Hydrog. Energy 2012, 37, 3160–3168. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, Y. Gut Microbiota Derived Metabolites in Cardiovascular Health and Disease. Protein Cell 2018, 9, 416–431. [Google Scholar] [CrossRef]
- Tang, X. Probiotic Roles of Clostridium butyricum in Piglets: Considering Aspects of Intestinal Barrier Function. Animals 2024, 14, 1069. [Google Scholar] [CrossRef]
- Meng, X.; Wu, S.; Hu, W.; Zhu, Z.; Yang, G.; Zhang, Y.; Qin, C.; Yang, L.; Nie, G. Clostridium butyricum Improves Immune Responses and Remodels the Intestinal Microbiota of Common Carp (Cyprinus carpio L.). Aquaculture 2021, 530, 735753. [Google Scholar] [CrossRef]
- Li, P.; Hou, D.; Zhao, H.; Wang, H.; Peng, K.; Cao, J. Dietary Clostridium butyricum Improves Growth Performance and Resistance to Ammonia Stress in Yellow Catfish (Pelteobagrus fulvidraco). Aquac. Nutr. 2022, 2022, 6965174. [Google Scholar] [CrossRef]
- Bi, X.; Lv, C.; Wang, M.; Chen, Y.; Lv, X.; Hua, S.; Zhang, X.; Yang, D.; Zhao, J.; Mu, C. Effects of Dietary Supplementation of Clostridium butyricum H129 on Growth Performance and Intestinal Health of Turbot, Scophthalmus maximus. Aquac. Rep. 2023, 30, 101544. [Google Scholar] [CrossRef]
- Gao, Q.; Qi, L.; Wu, T.; Wang, J. Ability of Clostridium butyricum to Inhibit Escherichia coli-Induced Apoptosis in Chicken Embryo Intestinal Cells. Vet. Microbiol. 2012, 160, 395–402. [Google Scholar] [CrossRef]
- Wang, T.; Fu, J.; Xiao, X.; Lu, Z.; Wang, F.; Jin, M.; Wang, Y.; Zong, X. Cbp22, a Novel Bacteriocin Isolated from Clostridium butyricum Zju-F1, Protects against Lps-Induced Intestinal Injury through Maintaining the Tight Junction Complex. Mediat. Inflamm. 2021, 2021, 8032125. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, X.; Wang, Y.; Liu, H.; Yu, X.; Li, L.; Ye, H.; Wang, S.; Gai, C.; Xu, L.; et al. Potential Effects of Dietary Probiotics with Chinese Herb Polysaccharides on the Growth Performance, Immunity, Disease Resistance, and Intestinal Microbiota of Rainbow Trout (Oncorhynchus mykiss). J. World Aquac. Soc. 2021, 52, 1194–1208. [Google Scholar] [CrossRef]
- Hayashi, A.; Sato, T.; Kamada, N.; Mikami, Y.; Matsuoka, K.; Hisamatsu, T.; Hibi, T.; Roers, A.; Yagita, H.; Ohteki, T.; et al. A Single Strain of Clostridium butyricum Induces Intestinal Il-10-Producing Macrophages to Suppress Acute Experimental Colitis in Mice. Cell Host Microbe 2013, 13, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Gaudier, E.; Jarry, A.; Blottière, H.M.; de Coppet, P.; Buisine, M.P.; Aubert, J.P.; Laboisse, C.; Cherbut, C.; Hoebler, C. Butyrate Specifically Modulates Muc Gene Expression in Intestinal Epithelial Goblet Cells Deprived of Glucose. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G1168–G1174. [Google Scholar] [CrossRef] [PubMed]
- Segain, J.P.; De La Blétiere, D.R.; Bourreille, A.; Leray, V.; Gervois, N.; Rosales, C.; Ferrier, L.; Bonnet, C.; Blottière, H.M.; Galmiche, J.-P. Butyrate Inhibits Inflammatory Responses through Nfkappab Inhibition: Implications for Crohn’s Disease. Gut 2000, 47, 397–403. [Google Scholar] [CrossRef]
- Pan, X.; Wu, T.; Zhang, L.; Song, Z.; Tang, H.; Zhao, Z. In Vitro Evaluation on Adherence and Antimicrobial Properties of a Candidate Probiotic Clostridium butyricum Cb2 for Farmed Fish. J. Appl. Microbiol. 2008, 105, 1623–1629. [Google Scholar] [CrossRef]
- Maren, N.A.; Duduit, J.R.; Huang, D.; Zhao, F.; Ranney, T.G.; Liu, W. Stepwise Optimization of Real-Time Rt-Pcr Analysis. Methods Mol. Biol. 2023, 2653, 317–332. [Google Scholar]
- Doyle, J.J. A Rapid DNA Isolation Procedure for Small Quantities of Fresh Leaf Tissue. Phytochem. Bull 1987, 19, 11–15. [Google Scholar]
- Magoč, T.; Salzberg, S.L. Flash: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-Filtering Vastly Improves Diversity Estimates from Illumina Amplicon Sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. Uchime Improves Sensitivity and Speed of Chimera Detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, H.; Gao, X.; Wang, J. The Intratumor Microbiota Signatures Associate with Subtype, Tumor Stage, and Survival Status of Esophageal Carcinoma. Front. Oncol. 2021, 11, 754788. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Chen, X.; Hou, D.; Chen, B.; Peng, K.; Huang, W.; Cao, J.; Zhao, H. Positive Effects of Dietary Clostridium butyricum Supplementation on Growth Performance, Antioxidant Capacity, Immunity and Viability against Hypoxic Stress in Largemouth Bass. Front. Immunol. 2023, 14, 1190592. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, J.; Huang, J.; Jiang, S. Effects of Dietary Clostridium butyricum on the Growth, Digestive Enzyme Activity, Antioxidant Capacity, and Resistance to Nitrite Stress of Penaeus monodon. Probiotics Antimicrob. Proteins 2019, 11, 938–945. [Google Scholar] [CrossRef]
- Chapman, M.A.; Grahn, M.F.; Boyle, M.A.; Hutton, M.; Rogers, J.; Williams, N.S. Butyrate Oxidation Is Impaired in the Colonic Mucosa of Sufferers of Quiescent Ulcerative Colitis. Gut 1994, 35, 73–76. [Google Scholar] [CrossRef]
- Kim, S.-K.; Guevarra, R.B.; Kim, Y.-T.; Kwon, J.; Kim, H.; Cho, J.H.; Kim, H.B.; Lee, J.-H. Role of Probiotics in Human Gut Microbiome-Associated Diseases. J. Microbiol. Biotechnol. 2019, 29, 1335–1340. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Chattopadhyay, R.; Mitra, S.; Crowe, S.E. Oxidative Stress: An Essential Factor in the Pathogenesis of Gastrointestinal Mucosal Diseases. Physiol. Rev. 2014, 94, 329–354. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.; Zhang, X.; Lu, Y.; Chen, H. New Insights in Intestinal Oxidative Stress Damage and the Health Intervention Effects of Nutrients: A Review. J. Funct. Foods 2020, 75, 104248. [Google Scholar] [CrossRef]
- Han, H.; He, N.; Pan, E.; Tan, X.; Yang, Z.; Li, X.; Shi, D.; Dong, J. Disruption of the Intestinal Barrier by Avermectin in Carp Involves Oxidative Stress and Apoptosis and Leads to Intestinal Inflammation. Pestic. Biochem. Physiol. 2023, 195, 105531. [Google Scholar] [CrossRef]
- Del Rio, D.; Stewart, A.J.; Pellegrini, N. A Review of Recent Studies on Malondialdehyde as Toxic Molecule and Biological Marker of Oxidative Stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Turrens, J.F. Mitochondrial Formation of Reactive Oxygen Species. J. Physiol. 2003, 552 Pt 2, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Tadese, D.A.; Wangari, M.R.; Zhou, Q.; Zheng, X.; Liu, B.; Tamiru, M.; Dagne, A.; Janssens, G.P.J.; Zhao, Y. Amelioration of Ammonia-Induced Intestinal Oxidative Stress by Dietary Clostridium butyricum in Giant Freshwater Prawn (Macrobrachium rosenbergii). Fish Shellfish Immunol. 2022, 131, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Cai, M.; Wang, Y.; Zhong, L.; Hu, Y.; Fu, G. Dietary Clostridium butyricum Metabolites Mitigated the Disturbances in Growth, Immune Response and Gut Health Status of Ctenopharyngodon Idella Subjected to High Cottonseed and Rapeseed Meal Diet. Fish Shellfish Immunol. 2024, 154, 109934. [Google Scholar] [CrossRef] [PubMed]
- Chasiotis, H.; Kolosov, D.; Bui, P.; Kelly, S.P. Tight Junctions, Tight Junction Proteins and Paracellular Permeability across the Gill Epithelium of Fishes: A Review. Respir. Physiol. Neurobiol. 2012, 184, 269–281. [Google Scholar] [CrossRef]
- Chen, W.; Song, T.; Li, D.; Chen, M.; Wang, P.; Ye, J. Effect of Dietary Clostridium butyricum Supplementation on Growth Performance, Immune Function, and Intestinal Health of Hybrid Grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). Front. Immunol. 2025, 16, 1557256. [Google Scholar] [CrossRef]
- Yang, M.; Wang, Y.; Patel, G.; Xue, Q.; Njateng, G.S.S.; Cai, S.; Cheng, G.; Kai, G. In Vitro and in Vivo Anti-Inflammatory Effects of Different Extracts from Epigynum Auritum through down-Regulation of Nf-Κb and Mapk Signaling Pathways. J. Ethnopharmacol. 2020, 261, 113105. [Google Scholar] [CrossRef]
- Li, H.; Zhou, Y.; Ling, H.; Luo, L.; Qi, D.; Feng, L. The Effect of Dietary Supplementation with Clostridium butyricum on the Growth Performance, Immunity, Intestinal Microbiota and Disease Resistance of Tilapia (Oreochromis niloticus). PLoS ONE 2019, 14, e0223428. [Google Scholar] [CrossRef]
- Shan, Q.; Wang, X.; Yang, H.; Zhu, Y.; Wang, J.; Yang, G. Bacillus Cereus Cwpfm Induces Colonic Tissue Damage and Inflammatory Responses through Oxidative Stress and the Nlrp3/Nf-Κb Pathway. Sci. Total Environ. 2024, 933, 173079. [Google Scholar] [CrossRef]
- Ye, D.; Ma, T.Y. Cellular and Molecular Mechanisms That Mediate Basal and Tumour Necrosis Factor-Alpha-Induced Regulation of Myosin Light Chain Kinase Gene Activity. J. Cell Mol. Med. 2008, 12, 1331–1346. [Google Scholar] [CrossRef]
- Shirazi, T.; Longman, R.; Corfield, A.; Probert, C. Mucins and Inflammatory Bowel Disease. Postgrad. Med. J. 2000, 76, 473–478. [Google Scholar] [CrossRef]
- Pothuraju, R.; Pai, P.; Chaudhary, S.; Siddiqui, J.A.; Cox, J.L.; Kaur, S.; Rachagani, S.; Roy, H.K.; Bouvet, M.; Batra, S.K. Depletion of Transmembrane Mucin 4 (Muc4) Alters Intestinal Homeostasis in a Genetically Engineered Mouse Model of Colorectal Cancer. Aging 2022, 14, 2025–2046. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.-T.; Stowell, S.R. The Role of Galectins in Immunity and Infection. Nat. Rev. Immunol. 2023, 23, 479–494. [Google Scholar] [CrossRef] [PubMed]
- Pietrzak, E.; Mazurkiewicz, J.; Slawinska, A. Innate Immune Responses of Skin Mucosa in Common Carp (Cyprinus carpio) Fed a Diet Supplemented with Galacto-oligosaccharides. Animals 2020, 10, 438. [Google Scholar] [CrossRef]
- Pereiro, P.; Varela, M.; Diaz-Rosales, P.; Romero, A.; Dios, S.; Figueras, A.; Novoa, B. Zebrafish Nk-Lysins: First Insights About Their Cellular and Functional Diversification. Dev. Comp. Immunol. 2015, 51, 148–159. [Google Scholar] [CrossRef]
- Larsen, A.; Mohammed, H.; Arias, C. Characterization of the Gut Microbiota of Three Commercially Valuable Warmwater Fish Species. J. Appl. Microbiol. 2014, 116, 1396–1404. [Google Scholar] [CrossRef]
- Gao, Q.; Liu, P.; Li, Y.; Song, D.; Long, W.; Wang, Z.; Yi, S.; Jiang, L. Gut Microbiota, Host Genetics and Phenotypes in Aquatic Animals: A Review. Aquac. Rep. 2023, 31, 101648. [Google Scholar] [CrossRef]
- Stoeva, M.K.; Garcia-So, J.; Justice, N.; Myers, J.; Tyagi, S.; Nemchek, M.; McMurdie, P.J.; Kolterman, O.; Eid, J. Butyrate-Producing Human Gut Symbiont, Clostridium butyricum, and Its Role in Health and Disease. Gut Microbes 2021, 13, 1907272. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira—A Candidate for the Next-Generation Probiotics. Gut Microbes 2021, 13, 1987783. [Google Scholar] [CrossRef]
- Bernardeau, M.; Lehtinen, M.J.; Forssten, S.D.; Nurminen, P. Importance of the Gastrointestinal Life Cycle of Bacillus for Probiotic Functionality. J. Food Sci. Technol. 2017, 54, 2570–2584. [Google Scholar] [CrossRef]
- Mukherjee, A.; Lordan, C.; Ross, R.P.; Cotter, P.D. Gut Microbes from the Phylogenetically Diverse Genus Eubacterium and Their Various Contributions to Gut Health. Gut Microbes 2020, 12, 1802866. [Google Scholar] [CrossRef]
- Wood, S.J.; Goldufsky, J.W.; Seu, M.Y.; Dorafshar, A.H.; Shafikhani, S.H. Pseudomonas aeruginosa Cytotoxins: Mechanisms of Cytotoxicity and Impact on Inflammatory Responses. Cells 2023, 12, 195. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.F.; Chambers, J.P.; Gupta, R.; Arulanandam, B.P. Chlamydia and Its Many Ways of Escaping the Host Immune System. J. Pathog. 2019, 2019, 8604958. [Google Scholar] [CrossRef] [PubMed]
- Litvak, Y.; Byndloss, M.X.; Bäumler, A.J. Colonocyte Metabolism Shapes the Gut Microbiota. Science 2018, 362, eaat9076. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.J.; Song, S.; Mason, K.; Pinkett, H.W. Selective Substrate Uptake: The Role of Atp-Binding Cassette (Abc) Importers in Pathogenesis. Biochim. Biophys. Acta Biomembr. 2018, 1860, 868–877. [Google Scholar] [CrossRef]
- Manson, M.D. Bacterial Motility and Chemotaxis. In Advances in Microbial Physiology; Rose, A.H., Ed.; Academic Press: London, UK, 1992; pp. 277–346. [Google Scholar]
- Palma, V.; Gutiérrez, M.S.; Vargas, O.; Parthasarathy, R.; Navarrete, P. Methods to Evaluate Bacterial Motility and Its Role in Bacterial–Host Interactions. Microorganisms 2022, 10, 563. [Google Scholar] [CrossRef]
- Marin, J.J.; Macias, R.I.; Briz, O.; Banales, J.M.; Monte, M.J. Bile Acids in Physiology, Pathology and Pharmacology. Curr. Drug Metab. 2015, 17, 4–29. [Google Scholar] [CrossRef]
- Rahman, S.; O’connor, A.L.; Becker, S.L.; Patel, R.K.; Martindale, R.G.; Tsikitis, V.L. Gut Microbial Metabolites and Its Impact on Human Health. Ann. Gastroenterol. 2023, 36, 360–368. [Google Scholar] [CrossRef]
- Brown, E.M.; Clardy, J.; Xavier, R.J. Gut Microbiome Lipid Metabolism and Its Impact on Host Physiology. Cell Host Microbe 2023, 31, 173–186. [Google Scholar] [CrossRef]
- Bakar, R.B.; Reimann, F.; Gribble, F.M. The Intestine as an Endocrine Organ and the Role of Gut Hormones in Metabolic Regulation. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 784–796. [Google Scholar] [CrossRef]
- Peng, F.; Zhang, H.; He, X.; Song, Z. Effects of Ursolic Acid on Intestinal Health and Gut Bacteria Antibiotic Resistance in Mice. Front. Physiol. 2021, 12, 650190. [Google Scholar] [CrossRef]
- Kou, R.-W.; Li, Z.-Q.; Wang, J.-L.; Jiang, S.-Q.; Zhang, R.-J.; He, Y.-Q.; Xia, B.; Gao, J.-M. Ganoderic Acid a Mitigates Inflammatory Bowel Disease through Modulation of Ahr Activity by Microbial Tryptophan Metabolism. J. Agric. Food Chem. 2024, 72, 17912–17923. [Google Scholar] [CrossRef]
- Decara, J.; Rivera, P.; López-Gambero, A.J.; Serrano, A.; Pavón, F.J.; Baixeras, E.; De Fonseca, F.R.; Suárez, J. Peroxisome Proliferator-Activated Receptors: Experimental Targeting for the Treatment of Inflammatory Bowel Diseases. Front. Pharmacol. 2020, 11, 730. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Z.; Qian, Y.; Peng, X.; Qin, C.; Ren, H.; Du, J.; Huang, C.; Pan, M.; Ou, W. Effects of Dietary Clostridium butyricum on Growth and Intestinal Mucosal Barrier Functions of Juvenile Channel Catfish (Ictalurus punctatus). Microorganisms 2025, 13, 1061. https://doi.org/10.3390/microorganisms13051061
Guo Z, Qian Y, Peng X, Qin C, Ren H, Du J, Huang C, Pan M, Ou W. Effects of Dietary Clostridium butyricum on Growth and Intestinal Mucosal Barrier Functions of Juvenile Channel Catfish (Ictalurus punctatus). Microorganisms. 2025; 13(5):1061. https://doi.org/10.3390/microorganisms13051061
Chicago/Turabian StyleGuo, Zihe, Ye Qian, Xiao Peng, Chanxia Qin, Huige Ren, Jingyi Du, Chengrui Huang, Mingzhu Pan, and Weihao Ou. 2025. "Effects of Dietary Clostridium butyricum on Growth and Intestinal Mucosal Barrier Functions of Juvenile Channel Catfish (Ictalurus punctatus)" Microorganisms 13, no. 5: 1061. https://doi.org/10.3390/microorganisms13051061
APA StyleGuo, Z., Qian, Y., Peng, X., Qin, C., Ren, H., Du, J., Huang, C., Pan, M., & Ou, W. (2025). Effects of Dietary Clostridium butyricum on Growth and Intestinal Mucosal Barrier Functions of Juvenile Channel Catfish (Ictalurus punctatus). Microorganisms, 13(5), 1061. https://doi.org/10.3390/microorganisms13051061