
Indole Acetic Acid: A Key Metabolite That Protects Marine Sulfitobacter mediterraneus Against Oxidative Stress


 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods and Materials
2.1. Bacterial Strains, Plasmids, and Growth Conditions
2.2. ROS Detection
2.3. Detection of IAA Produced by S. mediterraneus SC1-11 Under Oxidative Stress
2.4. RNA Extraction and RT-qPCR
2.5. Protective Effect of IAA Supplementation on H2O2-Induced Stress in S. mediterraneus SC1-11
2.6. Proteomics of S. mediterraneus SC1-11 Under Oxidative Stress
2.7. Quantification of S. mediterraneus SC1-11 Amino Acids Under Oxidative Stress
2.8. Protective Effect of Amino Acid Supplementations on H2O2-Induced Effects in S. mediterraneus SC1-11
2.9. Bioinformatics Analysis
2.10. Statistical Analyses
3. Results
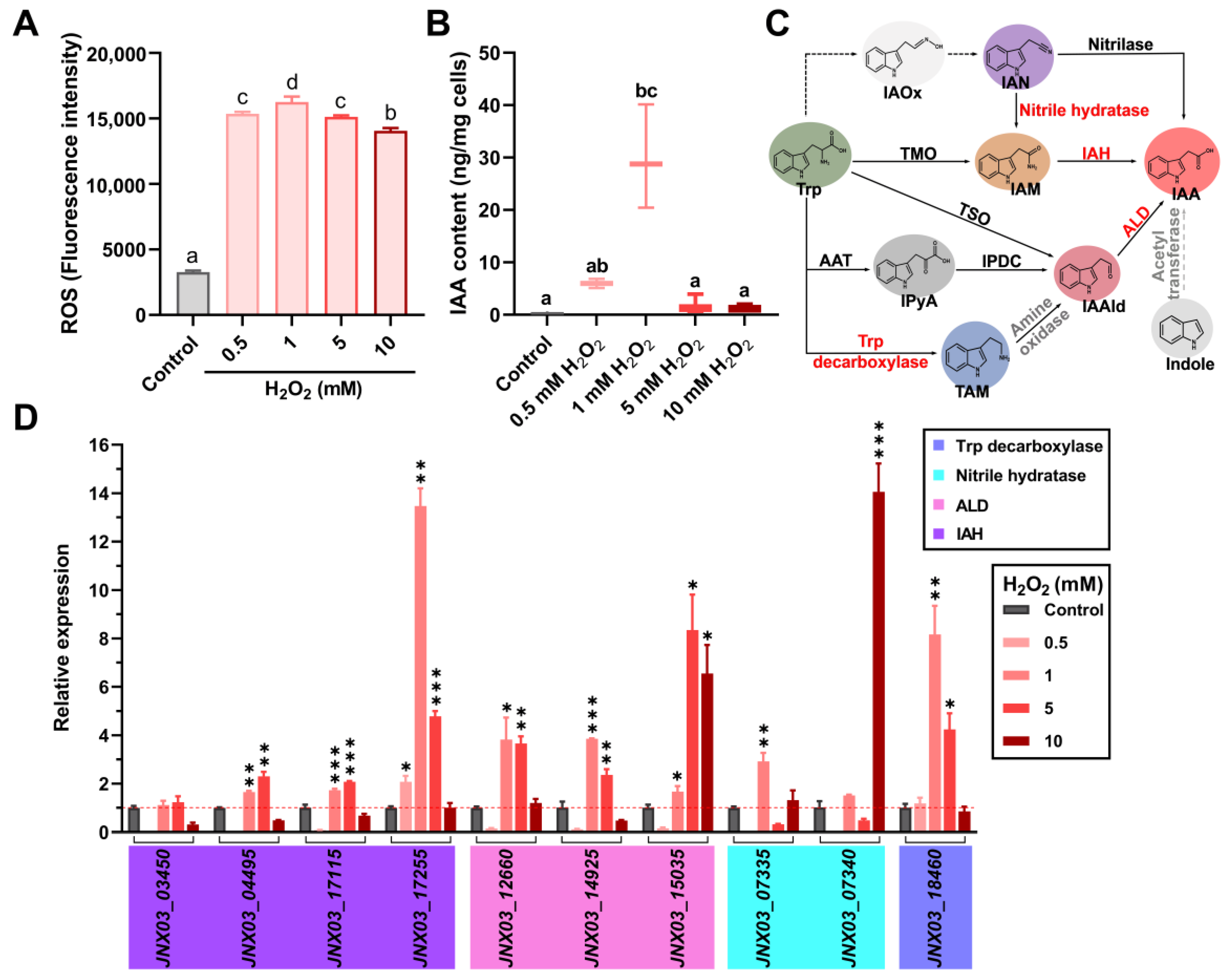
3.1. Oxidative Stress Stimulates the Biosynthesis of IAA in S. mediterraneus SC1-11
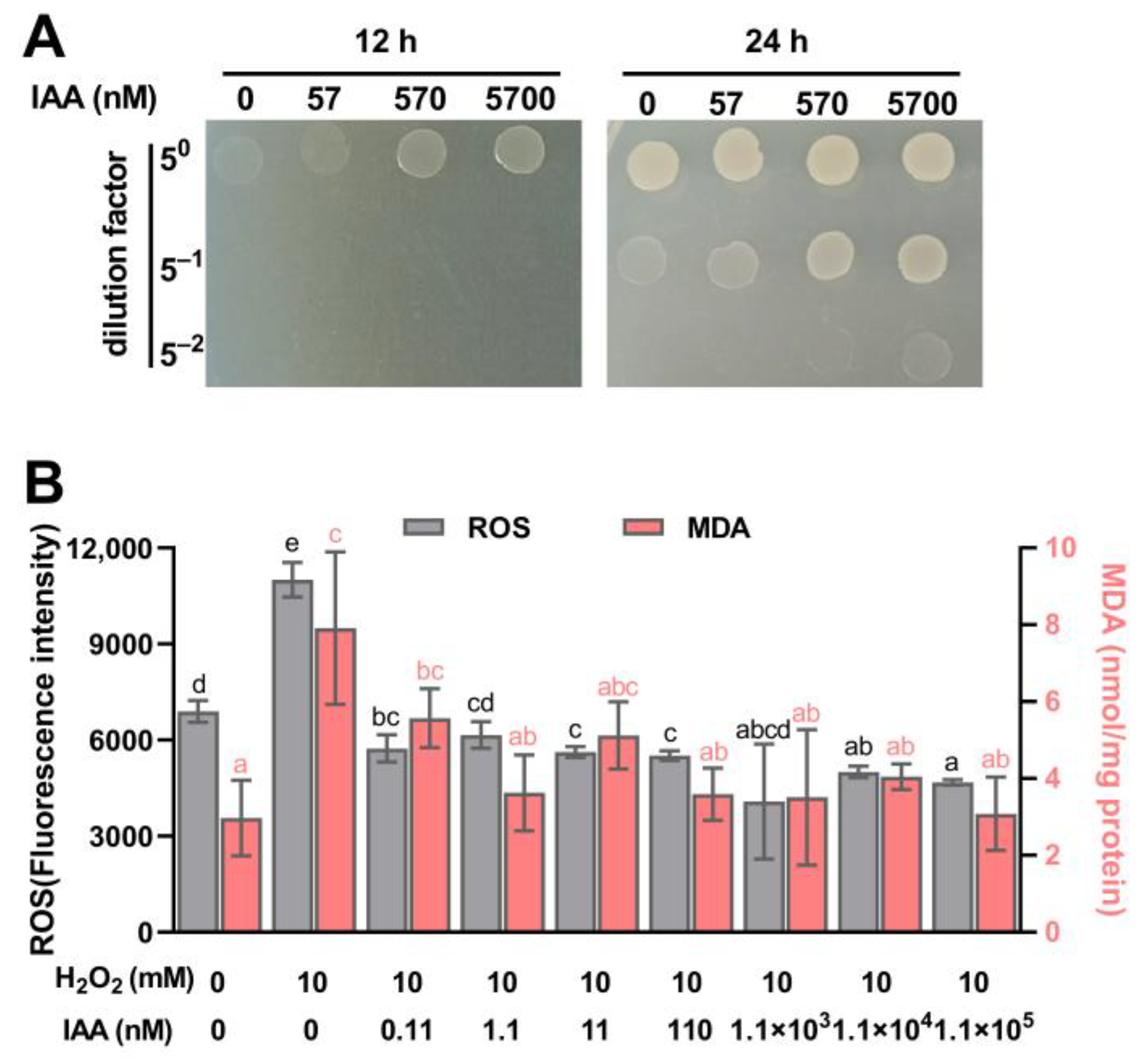
3.2. IAA Supplementation Protects S. mediterraneus SC1-11 from Oxidative Stress
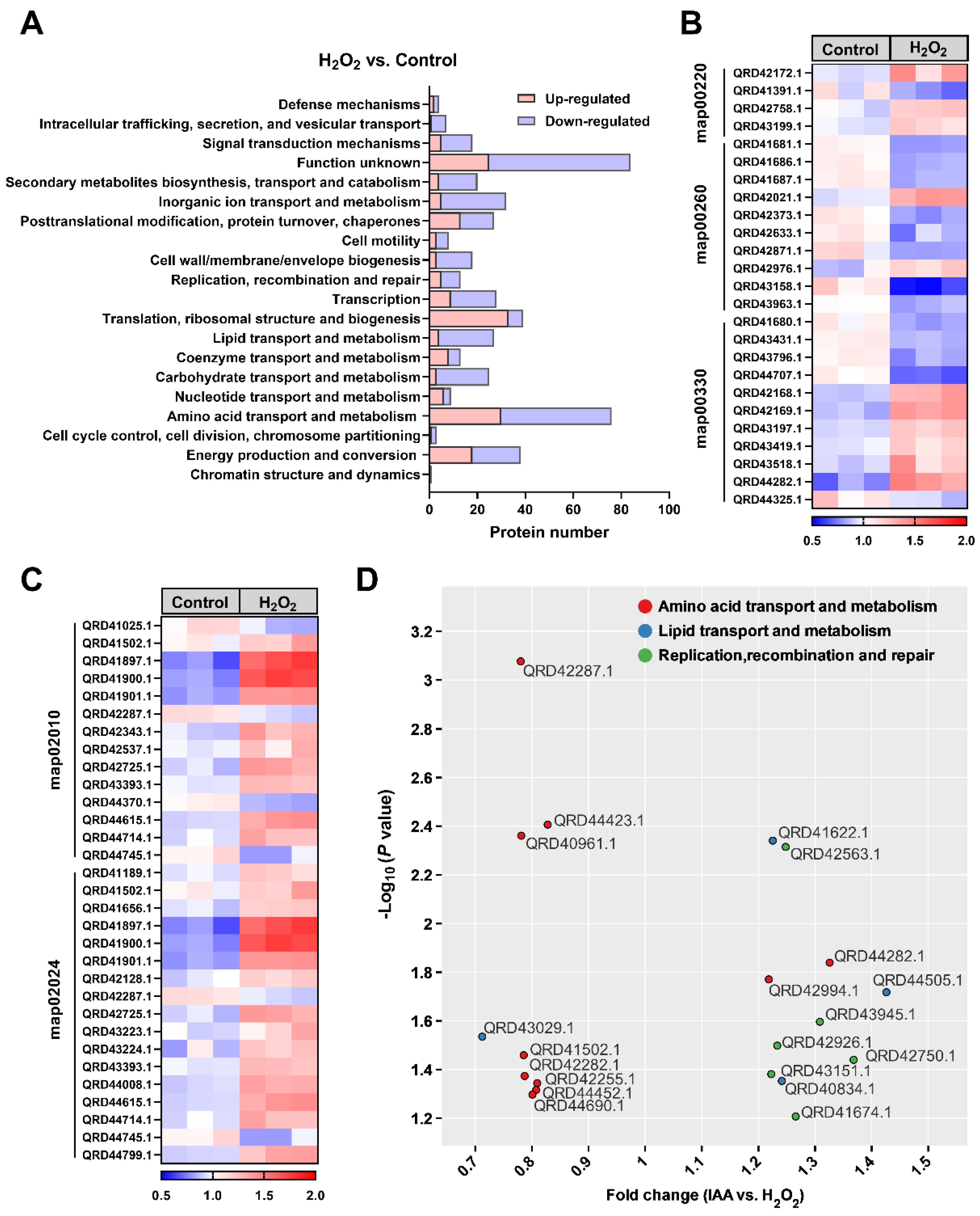
3.3. Oxidative Stress-Induced Proteomic and Metabolic Changes in SC1-11 Are Ameliorated by IAA Supplementation
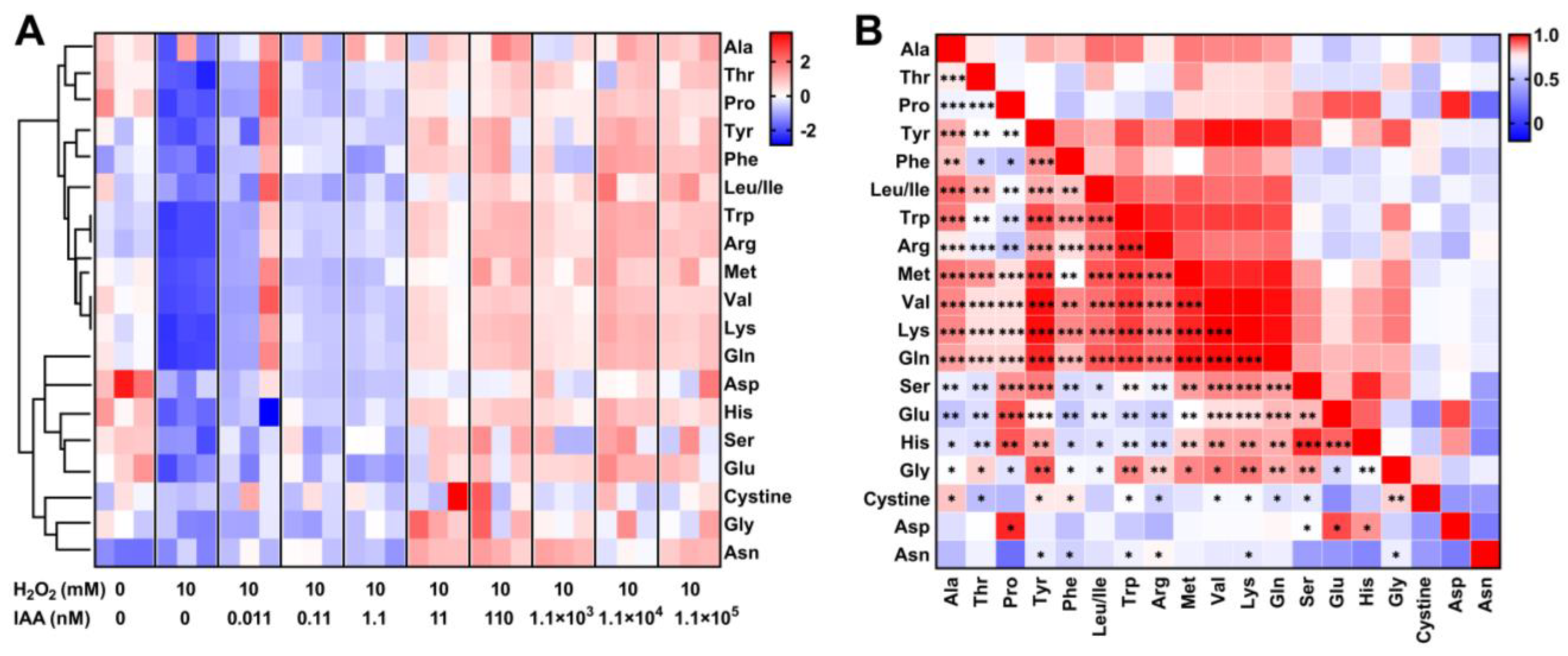
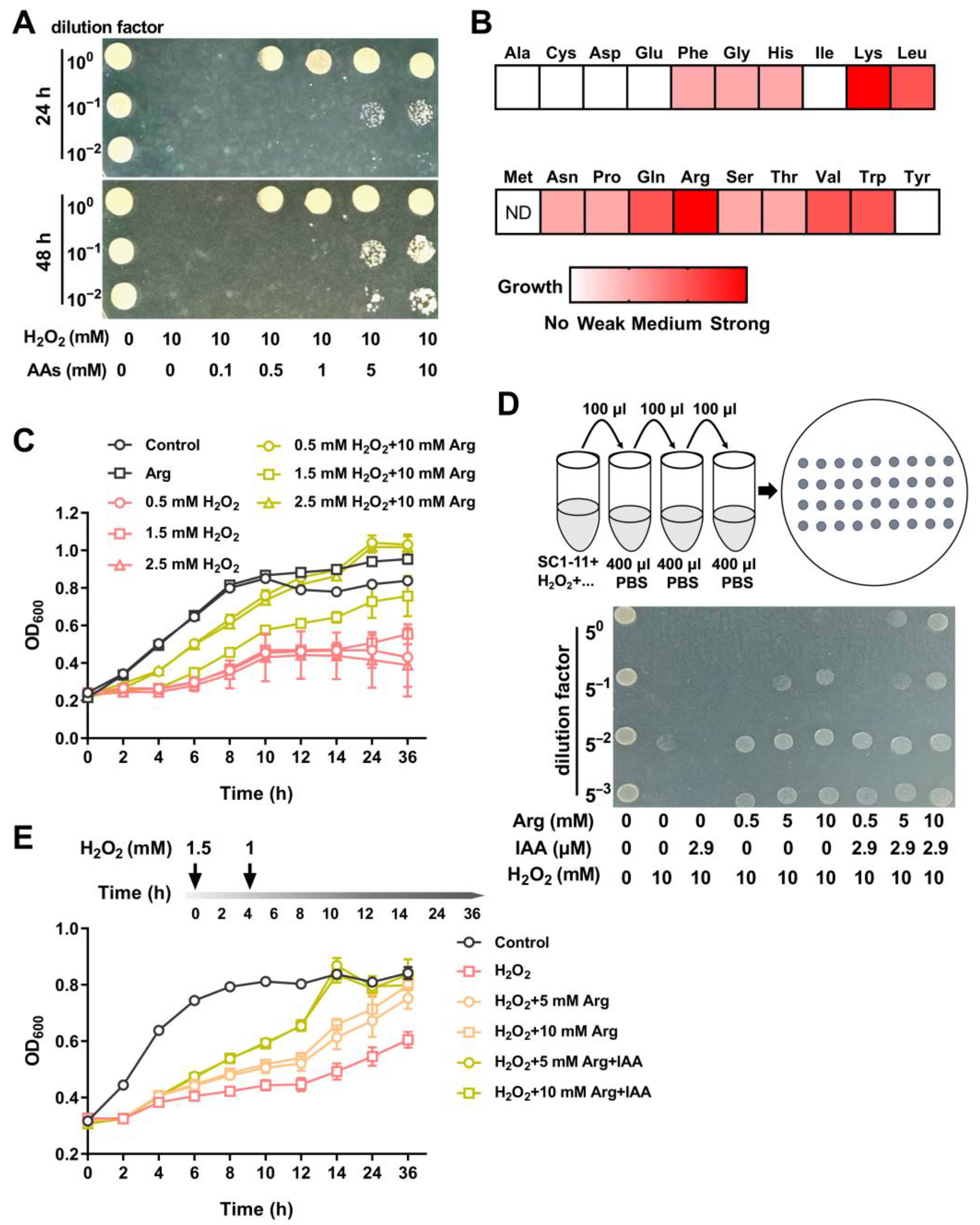
3.4. Amino Acids Supplementation Protects S. mediterraneus SC1-11 Against Oxidative Stress
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seymour, J.R.; Amin, S.A.; Raina, J.-B.; Stocker, R. Zooming in on the phycosphere: The ecological interface for phytoplankton–bacteria relationships. Nat. Microbiol. 2017, 2, 17065. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.B.; Kazamia, E.; Helliwell, K.E.; Kudahl, U.J.; Sayer, A.; Wheeler, G.L.; Smith, A.G. Cross-exchange of b-vitamins underpins a mutualistic interaction between Ostreococcus tauri and Dinoroseobacter shibae. ISME J. 2019, 13, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Croft, M.T.; Lawrence, A.D.; Raux-Deery, E.; Warren, M.J.; Smith, A.G. Algae acquire vitamin b12 through a symbiotic relationship with bacteria. Nature 2005, 438, 90–93. [Google Scholar] [CrossRef]
- Amin, S.A.; Hmelo, L.R.; Van Tol, H.M.; Durham, B.P.; Carlson, L.T.; Heal, K.R.; Morales, R.L.; Berthiaume, C.T.; Parker, M.S.; Djunaedi, B.; et al. Interaction and signalling between a cosmopolitan phytoplankton and associated bacteria. Nature 2015, 522, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Astafyeva, Y.; Gurschke, M.; Qi, M.; Bergmann, L.; Indenbirken, D.; De Grahl, I.; Katzowitsch, E.; Reumann, S.; Hanelt, D.; Alawi, M.; et al. Microalgae and bacteria interaction—Evidence for division of Diligence in the alga microbiota. Microbiol. Spectr. 2022, 10, e00633-22. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, W.; Wang, H. Physiological response and morphological changes of Heterosigma akashiwo to an algicidal compound prodigiosin. J. Hazard. Mater. 2020, 385, 121530. [Google Scholar] [CrossRef]
- Seyedsayamdost, M.R.; Case, R.J.; Kolter, R.; Clardy, J. The Jekyll-and-Hyde chemistry of Phaeobacter gallaeciensis. Nat. Chem. 2011, 3, 331–335. [Google Scholar] [CrossRef]
- Deng, Y.; Yu, R.; Grabe, V.; Sommermann, T.; Werner, M.; Vallet, M.; Zerfaß, C.; Werz, O.; Pohnert, G. Bacteria modulate microalgal aging physiology through the induction of extracellular vesicle production to remove harmful metabolites. Nat. Microbiol. 2024, 9, 2356–2368. [Google Scholar] [CrossRef]
- Sukenik, A.; Eshkol, R.; Livne, A.; Hadas, O.; Rom, M.; Tchernov, D.; Vardi, A.; Kaplan, A. Inhibition of growth and photosynthesis of the dinoflagellate Peridinium gatunense by Microcystis sp. (Cyanobacteria): A novel allelopathic mechanism. Limnol. Oceanogr. 2002, 47, 1656–1663. [Google Scholar] [CrossRef]
- Rijstenbil, J.W. Assessment of oxidative stress in the planktonic diatom Thalassiosira pseudonana in response to UVA and UVB radiation. J. Plankton Res. 2002, 24, 1277–1288. [Google Scholar] [CrossRef]
- Li, Z.; Wakao, S.; Fischer, B.B.; Niyogi, K.K. Sensing and responding to excess light. Annu. Rev. Plant Biol. 2009, 60, 239–260. [Google Scholar] [CrossRef] [PubMed]
- Van Creveld, S.G.; Rosenwasser, S.; Schatz, D.; Koren, I.; Vardi, A. Early perturbation in mitochondria redox homeostasis in response to environmental stress predicts cell fate in diatoms. ISME J. 2015, 9, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Thamatrakoln, K.; Korenovska, O.; Niheu, A.K.; Bidle, K.D. Whole-genome expression analysis reveals a role for death-related genes in stress acclimation of the diatom Thalassiosira pseudonana. Environ. Microbiol. 2012, 14, 67–81. [Google Scholar] [CrossRef]
- Thukral, M.; Allen, A.E.; Petras, D. Progress and challenges in exploring aquatic microbial communities using non-targeted metabolomics. ISME J. 2023, 17, 2147–2159. [Google Scholar] [CrossRef]
- Bell, W.; Mitchell, R. Chemotactic and growth responses of marine bacteria to algal extracellular products. Biol. Bull. 1972, 143, 265–277. [Google Scholar] [CrossRef]
- Fasnacht, M.; Polacek, N. Oxidative stress in bacteria and the central dogma of molecular biology. Front. Mol. Biosci. 2021, 8, 671037. [Google Scholar] [CrossRef] [PubMed]
- Ezraty, B.; Gennaris, A.; Barras, F.; Collet, J.-F. Oxidative stress, protein damage and repair in bacteria. Nat. Rev. Microbiol. 2017, 15, 385–396. [Google Scholar] [CrossRef]
- Kunkel, B.N.; Harper, C.P. The roles of auxin during interactions between bacterial plant pathogens and their hosts. J. Exp. Bot. 2018, 69, 245–254. [Google Scholar] [CrossRef]
- Mashiguchi, K.; Hisano, H.; Takeda-Kamiya, N.; Takebayashi, Y.; Ariizumi, T.; Gao, Y.; Ezura, H.; Sato, K.; Zhao, Y.; Hayashi, K.; et al. Agrobacterium tumefaciens enhances biosynthesis of two distinct auxins in the formation of crown galls. Plant Cell Physiol. 2019, 60, 29–37. [Google Scholar] [CrossRef]
- Tzipilevich, E.; Russ, D.; Dangl, J.L.; Benfey, P.N. Plant immune system activation is necessary for efficient root colonization by auxin-secreting beneficial bacteria. Cell Host Microbe 2021, 29, 1507–1520.e4. [Google Scholar] [CrossRef]
- Žižková, E.; Kubeš, M.; Dobrev, P.I.; Přibyl, P.; Šimura, J.; Zahajská, L.; Záveská Drábková, L.; Novák, O.; Motyka, V. Control of cytokinin and auxin homeostasis in cyanobacteria and algae. Ann. Bot. 2017, 119, 151–166. [Google Scholar] [CrossRef]
- Cheng, X.; Li, X.; Tong, M.; Wu, J.; Chan, L.L.; Cai, Z.; Zhou, J. Indole-3-acetic acid as a cross-talking molecule in algal-bacterial interactions and a potential driving force in algal bloom formation. Front. Microbiol. 2023, 14, 1236925. [Google Scholar] [CrossRef] [PubMed]
- Calatrava, V.; Hom, E.F.Y.; Guan, Q.; Llamas, A.; Fernández, E.; Galván, A. Genetic evidence for algal auxin production in Chlamydomonas and its role in algal-bacterial mutualism. iScience 2024, 27, 108762. [Google Scholar] [CrossRef] [PubMed]
- Khasin, M.; Cahoon, R.R.; Nickerson, K.W.; Riekhof, W.R. Molecular machinery of auxin synthesis, secretion, and perception in the unicellular chlorophyte alga Chlorella sorokiniana UTEX 1230. PLoS ONE 2018, 13, e0205227. [Google Scholar] [CrossRef]
- Matthews, J.L.; Khalil, A.; Siboni, N.; Bougoure, J.; Guagliardo, P.; Kuzhiumparambil, U.; DeMaere, M.; Le Reun, N.M.; Seymour, J.R.; Suggett, D.J.; et al. Coral endosymbiont growth is enhanced by metabolic interactions with bacteria. Nat. Commun. 2023, 14, 6864. [Google Scholar] [CrossRef] [PubMed]
- Khalil, A.; Bramucci, A.R.; Focardi, A.; Le Reun, N.; Willams, N.L.R.; Kuzhiumparambil, U.; Raina, J.-B.; Seymour, J.R. Widespread production of plant growth-promoting hormones among marine bacteria and their impacts on the growth of a marine diatom. Microbiome 2024, 12, 205. [Google Scholar] [CrossRef]
- Sison-Mangus, M.P.; Kempnich, M.W.; Appiano, M.; Mehic, S.; Yazzie, T. Specific bacterial microbiome enhances the sexual reproduction and auxospore production of the marine diatom, Odontella. PLoS ONE 2022, 17, e0276305. [Google Scholar] [CrossRef]
- Patidar, S.K. Metabolic interactions between microalgae and bacteria: Multifunctional ecological interplay and environmental applications. Algal Res. 2025, 86, 103904. [Google Scholar] [CrossRef]
- Segev, E.; Wyche, T.P.; Kim, K.H.; Petersen, J.; Ellebrandt, C.; Vlamakis, H.; Barteneva, N.; Paulson, J.N.; Chai, L.; Clardy, J.; et al. Dynamic metabolic exchange governs a marine algal-bacterial interaction. eLife 2016, 5, e17473. [Google Scholar] [CrossRef]
- Yang, Q.; Pande, G.S.J.; Wang, Z.; Lin, B.; Rubin, R.A.; Vora, G.J.; Defoirdt, T. Indole signalling and (micro)algal auxins decrease the virulence of Vibrio campbellii, a major pathogen of aquatic organisms. Environ. Microbiol. 2017, 19, 1987–2004. [Google Scholar] [CrossRef]
- Barak-Gavish, N.; Dassa, B.; Kuhlisch, C.; Nussbaum, I.; Brandis, A.; Rosenberg, G.; Avraham, R.; Vardi, A. Bacterial lifestyle switch in response to algal metabolites. Elife 2023, 12, e84400. [Google Scholar] [CrossRef] [PubMed]
- Hogle, S.L.; Brahamsha, B.; Barbeau, K.A. Direct heme uptake by phytoplankton-associated Roseobacter bacteria. mSystems 2017, 2, e00124-16. [Google Scholar] [CrossRef] [PubMed]
- Pukall, R.; Buntefuss, D.; Frühling, A.; Rohde, M.; Kroppenstedt, R.M.; Burghardt, J.; Lebaron, P.; Bernard, L.; Stackebrandt, E. Sulfitobacter mediterraneus sp. nov., a new sulfite-oxidizing member of the alpha-proteobacteria. Int. J. Syst. Bacteriol. 1999, 49 Pt 2, 513–519. [Google Scholar] [CrossRef]
- Qu, L.; Feng, X.; Chen, Y.; Li, L.; Wang, X.; Hu, Z.; Wang, H.; Luo, H. Metapopulation Structure of Diatom-Associated Marine Bacteria. bioRxiv 2021. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Z.; Zhao, J. Isolation, Characterization and Implications of the Bacterial Communities Associated with Established Cultures of Chattonella marina (Raphidophyceae) and Skeletonema costatum (Bacillariophyceae). Acta Oceanol. Sin. 2019, 38, 128–135. [Google Scholar] [CrossRef]
- Zhu, J.; Tang, S.; Cheng, K.; Cai, Z.; Chen, G.; Zhou, J. Microbial community composition and metabolic potential during a succession of algal blooms from Skeletonema sp. to Phaeocystis sp. Front. Microbiol. 2023, 14, 1147187. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, S.; Pratush, A.; Ye, X.; Xie, J.; Wei, H.; Sun, C.; Hu, Z. Acclimation of culturable bacterial communities under the stresses of different organic compounds. Front. Microbiol. 2018, 9, 225. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Olin-Sandoval, V.; Yu, J.S.L.; Miller-Fleming, L.; Alam, M.T.; Kamrad, S.; Correia-Melo, C.; Haas, R.; Segal, J.; Peña Navarro, D.A.; Herrera-Dominguez, L.; et al. Lysine harvesting is an antioxidant strategy and triggers underground polyamine metabolism. Nature 2019, 572, 249–253. [Google Scholar] [CrossRef]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. eggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Mas-Bargues, C.; Escrivá, C.; Dromant, M.; Borrás, C.; Viña, J. Lipid peroxidation as measured by chromatographic determination of malondialdehyde. Human plasma reference values in health and disease. Arch. Biochem. Biophys. 2021, 709, 108941. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. Diagnosing oxidative stress in bacteria: Not as easy as you might think. Curr. Opin. Microbiol. 2015, 24, 124–131. [Google Scholar] [CrossRef]
- Schmacht, M.; Lorenz, E.; Senz, M. Microbial production of glutathione. World J. Microbiol. Biotechnol. 2017, 33, 106. [Google Scholar] [CrossRef] [PubMed]
- Queval, G.; Thominet, D.; Vanacker, H.; Miginiac-Maslow, M.; Gakière, B.; Noctor, G. H2O2-activated up-regulation of glutathione in Arabidopsis involves induction of genes encoding enzymes involved in cysteine synthesis in the chloroplast. Mol. Plant 2009, 2, 344–356. [Google Scholar] [CrossRef]
- Krüger, A.; Vowinckel, J.; Mülleder, M.; Grote, P.; Capuano, F.; Bluemlein, K.; Ralser, M. Tpo1-mediated spermine and spermidine export controls cell cycle delay and times antioxidant protein expression during the oxidative stress response. EMBO Rep. 2013, 14, 1113–1119. [Google Scholar] [CrossRef]
- Gutsche, K.A.; Tran, T.B.T.; Vogel, R.F. Production of volatile compounds by Lactobacillus sakei from branched chain α-keto acids. Food Microbiol. 2012, 29, 224–228. [Google Scholar] [CrossRef]
- Bayliak, M.M.; Lylyk, M.P.; Vytvytska, O.M.; Lushchak, V.I. Assessment of antioxidant properties of alpha-keto acids in vitro and in vivo. Eur. Food Res. Technol. 2016, 242, 179–188. [Google Scholar] [CrossRef]
- Maqbool, A.; Horler, R.S.P.; Muller, A.; Wilkinson, A.J.; Wilson, K.S.; Thomas, G.H. The substrate-binding protein in bacterial ABC transporters: Dissecting roles in the evolution of substrate specificity. Biochem. Soc. Trans. 2015, 43, 1011–1017. [Google Scholar] [CrossRef]
- Ma, Y.; Li, X.; Wang, F.; Zhang, L.; Zhou, S.; Che, X.; Yu, D.; Liu, X.; Li, Z.; Sun, H.; et al. Structural and biochemical characterization of the key components of an auxin degradation operon from the rhizosphere bacterium Variovorax. PLoS Biol. 2023, 21, e3002189. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gan, Y.; Cai, R.; Cai, G.; Aweya, J.J.; Xie, J.; Chen, Z.; Wang, H. Indole Acetic Acid: A Key Metabolite That Protects Marine Sulfitobacter mediterraneus Against Oxidative Stress. Microorganisms 2025, 13, 1014. https://doi.org/10.3390/microorganisms13051014
Gan Y, Cai R, Cai G, Aweya JJ, Xie J, Chen Z, Wang H. Indole Acetic Acid: A Key Metabolite That Protects Marine Sulfitobacter mediterraneus Against Oxidative Stress. Microorganisms. 2025; 13(5):1014. https://doi.org/10.3390/microorganisms13051014
Chicago/Turabian StyleGan, Yongliang, Runlin Cai, Guanjing Cai, Jude Juventus Aweya, Jianmin Xie, Ziming Chen, and Hui Wang. 2025. "Indole Acetic Acid: A Key Metabolite That Protects Marine Sulfitobacter mediterraneus Against Oxidative Stress" Microorganisms 13, no. 5: 1014. https://doi.org/10.3390/microorganisms13051014
APA StyleGan, Y., Cai, R., Cai, G., Aweya, J. J., Xie, J., Chen, Z., & Wang, H. (2025). Indole Acetic Acid: A Key Metabolite That Protects Marine Sulfitobacter mediterraneus Against Oxidative Stress. Microorganisms, 13(5), 1014. https://doi.org/10.3390/microorganisms13051014