
The Effects of Varying Concentrations of Didecyl Methylpropyl Ammonium Iodide (DMPAI) on the Structure and Function of Soil Bacterial Communities in the Lake–Terrestrial Ecotone
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Methods
2.2.1. Determination of DMPAI Concentration
2.2.2. 16S rRNA Sequencing
2.2.3. Function Prediction
3. Results
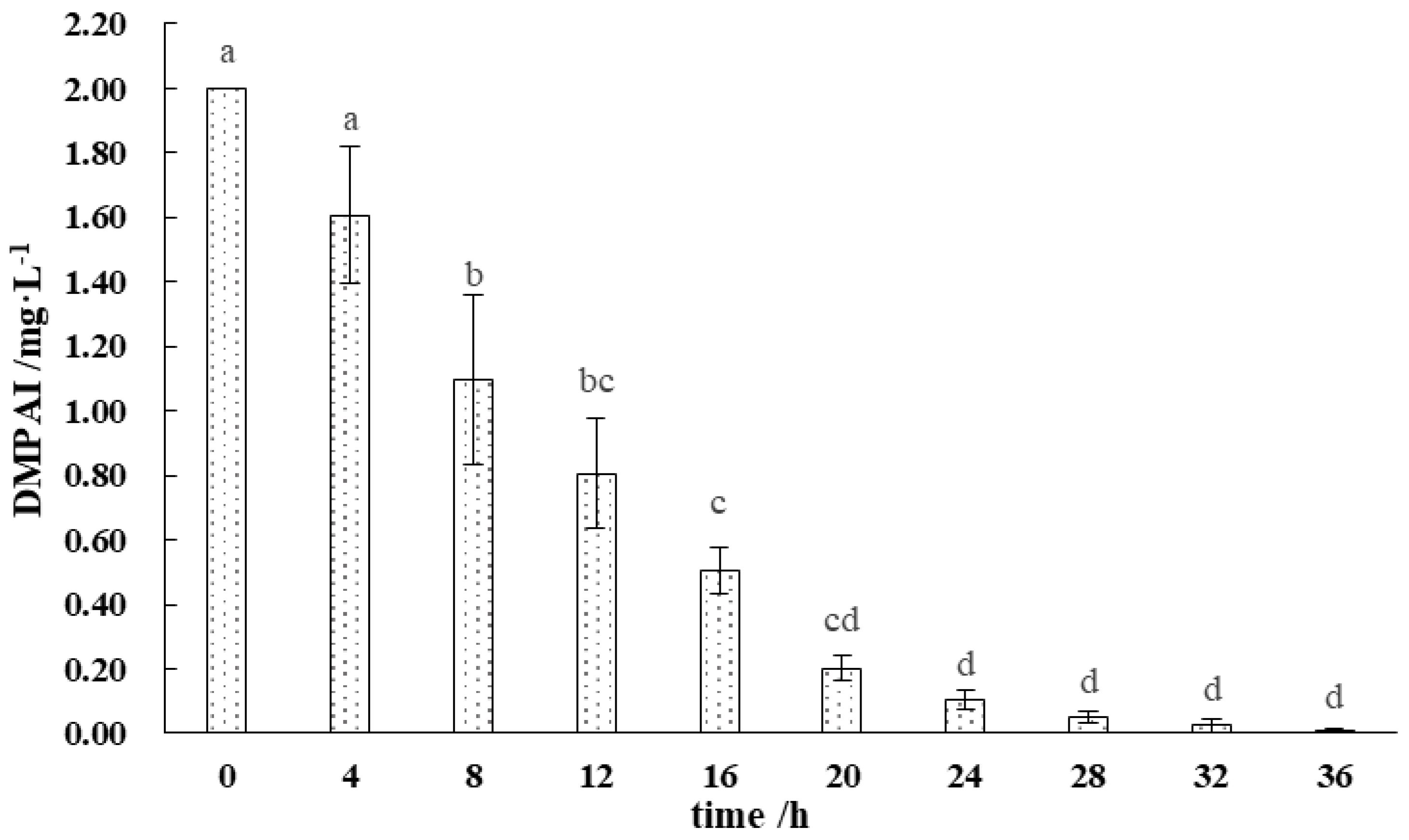
3.1. Degradation Characteristics of DMPAI in the Lake–Terrestrial Ecotone Waters
3.2. Response of Soil Bacteria to DMPAI in the Lake–Terrestrial Ecotone
3.2.1. Effect of DMPAI on Bacterial Community Species Diversity
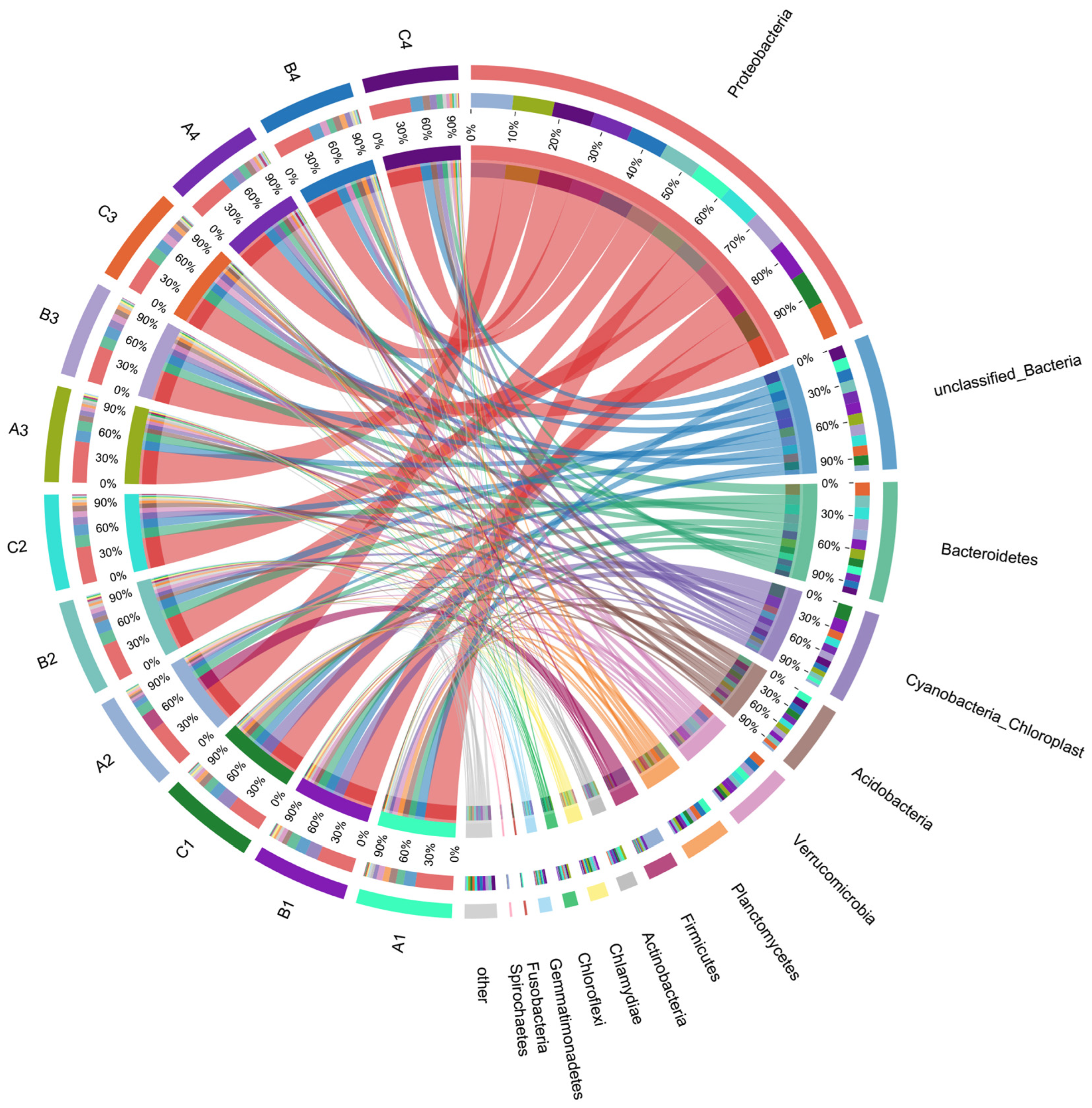
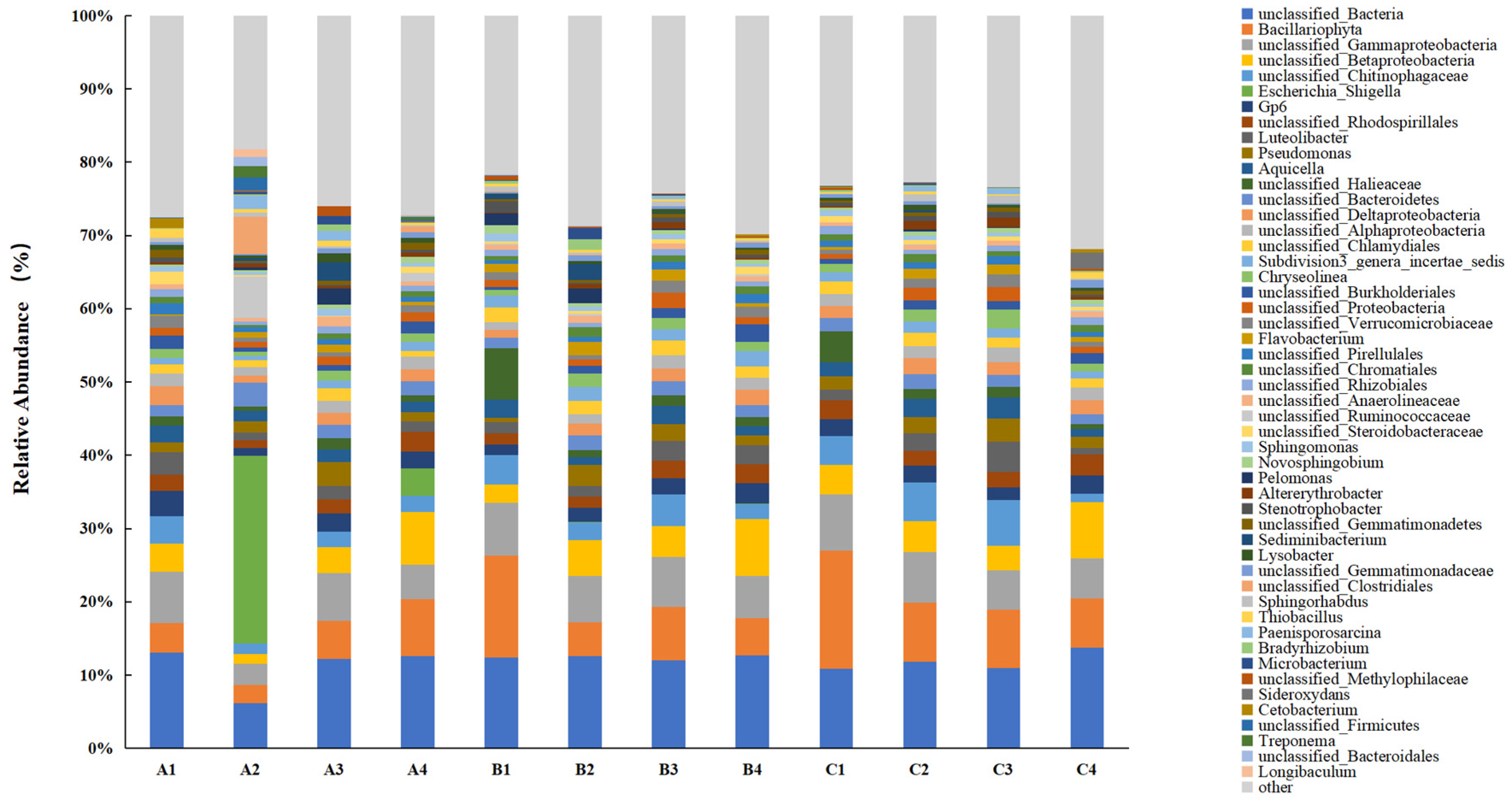
3.2.2. Soil Bacterial Community Evolution in the Lake–Terrestrial Ecotone
3.2.3. Functional Gene Abundance in Bacteria
4. Discussion
5. Conclusions
- (1)
- DMPAI at concentrations less than 2 mg L−1 is rapidly degraded by microorganisms in the lake–terrestrial ecotone and completely disappears in 36 h.
- (2)
- The addition of DMPAI altered the microbial community structure in the lake–terrestrial ecotone, as evidenced by an increase in the diversity index and the proliferation of microorganisms capable of tolerating and degrading DMPAI, such as Pseudomonas and Flavobacterium, within a short period. These changes were typically observed after 10 d and generally recovered, not persisting for extended periods.
- (3)
- Functional genes involved in carbon, nitrogen, and sulfur cycling are more impaired in a lake–terrestrial ecotone after DMPAI addition. The destabilization of the microbial community may lead to a short-term increase in pathogenic bacteria during the recovery process. This phenomenon was more pronounced in environments with higher concentrations of DMPAI. Therefore, the concentration of DMPAI should be controlled within the range of 0.5 to 2.0 mg L−1.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, S.L.; Li, J.S.; Zhang, B.; Spyrakos, E.; Tyler, A.N.; Shen, Q.; Zhang, F.F.; Kuster, T.; Lehmann, M.K.; Wu, Y.H.; et al. Trophic state assessment of global inland waters using a MODIS-derived Forel-Ule index. Remote Sens. Environ. 2018, 217, 444–460. [Google Scholar] [CrossRef]
- Ho, J.C.; Michalak, A.M.; Pahlevan, N. Widespread global increase in intense lake phytoplankton blooms since the 1980s. Nature 2019, 574, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Sheng, H.; Guo, H.C.; Liu, H.; Yang, Y.H. Reversion and analysis on cyanobacteria bloom in Waihai of Lake Dianchi. Acta Ecol. Sin. 2012, 32, 56–63. [Google Scholar] [CrossRef]
- Liu, J.T.; Yang, Y.S.; Jiang, J.H.; Gao, J.F. Risk evaluation method of cyanobacteria bloom hazard in Taihu Lake. China Environ. Sci. 2011, 31, 498–503. [Google Scholar]
- Yuan, Q.; Yu, L.; Li, W.H.; Yang, H.Y.; Wang, R.; Xu, D.M. Dynamic differences of microcyslins and their relationship with environmental faclors in spring in Lake Chaohu and Lake Taihu. J. Environ. Eng. Technol. 2021, 11, 267–277. [Google Scholar]
- Zhang, Z.X.; Wang, Y.C.; Liu, Q.H.; Jiao, X.D.; Wang, L. Research progress on biological function of microcystins. Asian J. Ecotoxicol. 2023, 18, 128–140. [Google Scholar]
- Hu, L.L.; Shan, K.; Lin, L.Z.; Shen, W.; Huang, L.C.; Gan, N.Q.; Song, L.R. Multi-year assessment of toxic genotypes and microcystin concentration in northern Lake Taihu, China. Toxins 2016, 8, 23. [Google Scholar] [CrossRef]
- Yang, Z.; Kong, F.X.; Zhang, M. Groundwater contamination by microcystin from toxic cyanobacteria blooms in Lake Chaohu, China. Environ. Monit. Assess. 2016, 188, 280. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Boyer, G.L. Health impacts from cyanobacteria harmful algae blooms: Implications for the North American Great Lakes. Harmful Algae 2016, 54, 194–212. [Google Scholar] [CrossRef]
- Wei, Q.; Wang, L.; Ma, X.M.; Gan, Y.H.; Zheng, S.M. Research status and prospect of management countermeasures for cyanobacteria blooms in freshwater lakes. J. North China Univ. Water Resour. Electr. Power (Nat. Sci. Ed.) 2021, 42, 22–30. [Google Scholar]
- Liu, j.; Zhang, H.; Yang, W.; Jie, G.; Qiong, K. Studies on biquaternary ammonium salt algaecide for removing red tide. Mar. Environ. Sci. 2003, 22, 68–71. [Google Scholar]
- Sütterlin, H.; Alexy, R.; Coker, A.; Kümmerer, K. Mixtures of quaternary ammonium compounds and anionic organic compounds in the aquatic environment: Elimination and biodegradability in the closed bottle test monitored by LC-MS/MS. Chemosphere 2008, 72, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Lavorgna, M.; Russo, C.; D’Abrosca, B.; Parrella, A.; Isidori, M. Toxicity and genotoxicity of the quaternary ammonium compound benzalkonium chloride (BAC) using Daphnia magna and Ceriodaphnia dubia as model systems. Environ. Pollut. 2016, 210, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Fan, Z.Y.; Zeng, L.Q.; Li, C.H.; Ye, C. Studies on the algae removal effect and toxic effects of new algae removal agent didecyl methylpropyl ammonium iodide (DMPAI) on aquatic organisms. J. Environ. Eng. Technol. 2024, 14, 1357–1363. [Google Scholar]
- Bodelier, P.L.E. Toward understanding, managing, and protecting microbial ecosystems. Front. Microbiol. 2020, 11, 653. [Google Scholar] [CrossRef]
- Zhang, C.; Tezel, U.; Li, K.X.; Liu, D.F.; Ren, R.; Du, J.X.; Pavlostathis, S.G. Evaluation and modeling of benzalkonium chloride inhibition and biodegradation in activated sludge. Water Res. 2011, 45, 1238–1246. [Google Scholar] [CrossRef]
- Agathokleous, E.; Barceló, D.; Iavicoli, I.; Tsatsakis, A.; Calabrese, E.J. Disinfectant-induced hormesis: An unknown environmental threat of the application of disinfectants to prevent SARS-CoV-2 infection during the COVID-19 pandemic? Environ. Pollut. 2022, 292, 118429. [Google Scholar] [CrossRef]
- Deng, M.; Yu, L.Y.; Peng, Q. Synthesis and research on flocculation performance of cationic polyacrylamide with ultrahigh molecular weight. Chem. Res. Appl. 2023, 35, 654–661. [Google Scholar]
- Zhang, C.; Zhao, X.; Liang, A.J.; Li, Y.Y.; Song, Q.Y.; Li, X.Y.; Li, D.P.; Hou, N. Insight into the soil aggregate-mediated restoration mechanism of degraded black soil via biochar addition: Emphasizing the driving role of core microbial communities and nutrient cycling. Environ. Res. 2023, 228, 115895. [Google Scholar] [CrossRef]
- Xu, F.; Cao, F.Q.; Kong, Q.; Zhou, L.L.; Yuan, Q.; Zhu, Y.J.; Wang, Q.; Du, Y.D.; Wang, Z.D. Electricity production and evolution of microbial community in the constructed wetland-microbial fuel cell. Chem. Eng. J. 2018, 339, 479–486. [Google Scholar] [CrossRef]
- Ji, B.; Zhang, M.; Gu, J.; Ma, Y.Q.; Liu, Y. A self-sustaining synergetic microalgal-bacterial granular sludge process towards energy-efficient and environmentally sustainable municipal wastewater treatment. Water Res. 2020, 179, 115884. [Google Scholar] [CrossRef] [PubMed]
- Nixon, S.L.; Daly, R.A.; Borton, M.A.; Solden, L.M.; Welch, S.A.; Cole, D.R.; Mouser, P.J.; Wilkins, M.J.; Wrighton, K.C. Genome-Resolved Metagenomics Extends the Environmental Distribution of the Verrucomicrobia Phylum to the Deep Terrestrial Subsurface. Msphere 2019, 4, e00613-19. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.S.; Zhao, Y.Q.; Pan, F.X.; Yang, B.S.; Wang, H.; Wang, S.Z.; Yuan, Y.R. Enhanced nitrogen removal in filled-and-drained vertical flow constructed wetlands: Microbial responses to aeration mode and carbon source. Environ. Sci. Pollut. Res. 2020, 27, 37650–37659. [Google Scholar] [CrossRef] [PubMed]
- Li, C.H.; Zhang, C.X.; Tang, L.S.; Xiong, Z.Q.; Wang, B.Z.; Jia, Z.J.; Li, Y. Effect of long-term fertilizing regime on soil microbial diversity and soil property. Acta Microbiol. Sin. 2014, 54, 319–329. [Google Scholar]
- Onalenna, O.; Rahube, T.O. Assessing bacterial diversity and antibiotic resistance dynamics in wastewater effluent-irrigated soil and vegetables in a microcosm setting. Heliyon 2022, 8, e09089. [Google Scholar] [CrossRef]
- Chen, L.; Li, F.; Li, W.; Ning, Q.; Li, J.W.; Zhang, J.B.; Ma, D.H.; Zhang, C.Z. Organic amendment mitigates the negative impacts of mineral fertilization on bacterial communities in Shajiang black soil. Appl. Soil Ecol. 2020, 150, 103457. [Google Scholar] [CrossRef]
- Campos, M.; Rilling, J.I.; Acuña, J.J.; Valenzuela, T.; Larama, G.; Peña-Cortés, F.; Ogram, A.; Jaisi, D.P. Spatiotemporal variations and relationships of phosphorus, phosphomonoesterases, and bacterial communities in sediments from two Chilean rivers. Sci. Total Environ. 2021, 776, 145782. [Google Scholar] [CrossRef]
- Su, P.Z.; Gao, P.; Sun, W.M.; Gao, W.L.; Xu, F.Q.; Wang, Q.; Xiao, E.Z.; Soleimani, M.; Sun, X.X. Keystone taxa and functional analysis in arsenic and antimony co-contaminated rice terraces. Environ. Sci. Pollut. Res. 2022, 29, 61236–61246. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Zhang, Z.G.; Yuen, G.Y. The Role of Chitinase Production by Stenotrophomonas maltophilia Strain C3 in Biological Control of Bipolaris sorokiniana. Phytopathology 2000, 90, 384–389. [Google Scholar] [CrossRef]
- Hagemann, M.; Ribbeck-Busch, K.; Klähn, S.; Hasse, D.; Steinbruch, R.; Berg, G. The plant-associated bacterium Stenotrophomonas rhizophila expresses a new enzyme for the synthesis of the compatible solute glucosylglycerol. J. Bacteriol. 2008, 190, 5898–5906. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.T.; Wang, R.Y.; Zhao, X.; Wang, Y.; Guo, Z.H.; Zhang, Y.B.; Xie, Z.K. Effect of Compost on Rhizosphere Microbial Community of Lanzhou-lily Based on High Throughput Sequencing. Acta Agric. Boreali-Occident. Sin. 2017, 26, 437–447. [Google Scholar]
- Du, S.Y.; Yu, M.; Liu, F.H.; Xiao, L.L.; Zhang, H.X.; Tao, J.; Gu, W.; Gu, J.Y.; Chen, X. Effect of facility management regimes on soil bacterial diversity and community structure. Chin. J. Eco-Agric. 2017, 25, 1615–1625. [Google Scholar]
- Lin, B.H.; Dai, Y.C.; Zhu, C.J.; Su, Z.Q.; Zhang, X.F. Analysis on characteristics of gut microbiota of children with Norovirus acute gastroenteritis. J. Pathog. Biol. 2023, 18, 519–524. [Google Scholar]
- Nalecz, J.G.; Grabinska, S.E.; Narkiewicz, P. The toxicity of cationic surfactants in four bioassays. Ecotoxicol. Environ. Saf. 2003, 54, 87–91. [Google Scholar] [CrossRef]
- Zhu, M.J.; Ge, F.; Zhu, R.L.; Wang, X.Y.; Zheng, X.Y. A DFT-based QSAR study of the toxicity of quaternary ammonium compounds on Chlorella vulgaris. Chemosphere 2010, 80, 46–52. [Google Scholar] [CrossRef]
- Carbajo, J.B.; Perdigón-Melón, J.A.; Peeter, A.L.; Rosal, R.; Letón, P.; García-Calvo, E. Personal care product preservatives: Risk assessment and mixture toxicities with an industrial wastewater. Water Res. 2015, 72, 174–185. [Google Scholar] [CrossRef]
- Tandukar, M.; Oh, S.; Tezel, U.; Konstantinidis, K.T.; Pavlostathis, S.G. Long-term exposure to benzalkonium chloride disinfectants results in change of microbial community structure and increased antimicrobial resistance. Environ. Sci. Technol. 2013, 47, 9730–9738. [Google Scholar] [CrossRef]
- Zhang, C.; Cui, F.; Zeng, G.M.; Jiang, M.; Yang, Z.Z.; Yu, Z.J.; Zhu, M.Y.; Shen, L.Q. Quaternary ammonium compounds (QACs): A review on occurrence, fate and toxicity in the environment. Sci. Total Environ. 2015, 15, 352–362. [Google Scholar] [CrossRef]
- Clara, M.; Scharf, S.; Scheffknecht, C.; Gans, O. Occurrence of selected surfactants in untreated and treated sewage. Water Res. 2007, 41, 4339–4348. [Google Scholar] [CrossRef]
- Garcia, M.T.; Ribosa, I.; Guindulain, T.; Sánchez-Leal, J.; Vives-Rego, J. Fate and effect of monoalkyl quaternary ammonium surfactants in the aquatic environment. Environ. Pollut. 2001, 111, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Nica, V.D.; Gallet, J.; Villa, S.; Mezzanotte, V. Toxicity of quaternary ammonium compounds (QACs) as single compounds and mixtures to aquatic non-target microorganisms: Experimental data and predictive models. Ecotoxicol. Environ. Saf. 2017, 142, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Fan, Z.Y.; Zeng, L.Q.; Li, C.H.; Ye, C.; Zheng, R.W. Effect of sodium dodecyl sulfate (SDS) on microbial community structure and function in lake–terrestrial ecotones: A simulation experiment. Environ. Technol. Innov. 2024, 34, 103594. [Google Scholar] [CrossRef]
- Worthington, R.J.; Blackledge, M.S.; Melander, C. Small-molecule inhibition of bacterial two-component systems to combat antibiotic resistance and virulence. Future Med. Chem. 2013, 5, 1265–1284. [Google Scholar] [CrossRef]
- Umar, Z.; Chen, Q.; Tang, B.; Xu, Y.; Wang, J.; Zhang, H.; Ji, K.; Jia, X.; Feng, Y. The poultry pathogen Riemerella anatipestifer appears as a reservoir for Tet(X) tigecycline resistance. Environ. Microbiol. 2021, 23, 7465–7482. [Google Scholar] [CrossRef]
- Chen, Q.W.; Gong, X.W.; Zheng, F.Y.; Ji, G.; Li, S.D.; Stipkovits, L.; Szathmary, S.; Liu, Y. Interplay between the phenotype and genotype, and efflux pumps in drug-resistant strains of Riemerella anatipestifer. Front. Microbiol. 2018, 9, 2136. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | PH | TN (‰) | TP (‰) | SOM (‰) |
|---|---|---|---|---|
| A | 8.75 | 0.46 | 0.78 | 5.58 |
| B | 8.80 | 0.35 | 0.77 | 5.31 |
| C | 8.88 | 0.31 | 0.73 | 5.03 |
| Functional Genes | A1 | B1 | C1 | A2 | B2 | C2 | A3 | B3 | C3 | A4 | B4 | C4 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Nitrogen cycle | Aerobic nitrite oxidation | 351 | 343 | 1049 | 36 | 270 | 179 | 134 | 142 | 67 | 152 | 137 | 239 |
| Nitrification | 397 | 357 | 1086 | 51 | 306 | 315 | 139 | 157 | 92 | 293 | 147 | 329 | |
| Nitrogen fixation | 178 | 275 | 354 | 131 | 852 | 218 | 478 | 154 | 59 | 300 | 173 | 271 | |
| Nitrate respiration | 101 | 195 | 144 | 53 | 267 | 130 | 103 | 135 | 57 | 302 | 179 | 240 | |
| Nitrate reduction | 279 | 267 | 466 | 9645 | 430 | 187 | 205 | 212 | 72 | 1483 | 290 | 278 | |
| Nitrogen respiration | 101 | 195 | 144 | 53 | 279 | 130 | 103 | 135 | 57 | 302 | 179 | 240 | |
| Ureolysis | 382 | 100 | 228 | 60 | 267 | 231 | 324 | 104 | 103 | 103 | 48 | 68 | |
| Carbon cycle | Methanotrophy | 165 | 26 | 100 | 3 | 19 | 3 | 27 | 19 | 5 | 13 | 15 | 32 |
| Methanogenesis | 11 | 0 | 12 | 37 | 45 | 2 | 6 | 0 | 0 | 16 | 0 | 9 | |
| Methanol oxidation | 63 | 410 | 430 | 24 | 108 | 62 | 658 | 52 | 21 | 71 | 66 | 108 | |
| Methylotrophy | 228 | 436 | 530 | 46 | 127 | 65 | 685 | 71 | 26 | 91 | 81 | 140 | |
| Chitinolysis | 1207 | 74 | 465 | 144 | 204 | 368 | 597 | 254 | 102 | 205 | 84 | 75 | |
| Aromatic compound degradation | 52 | 139 | 43 | 47 | 142 | 54 | 148 | 5 | 0 | 19 | 1 | 3 | |
| Hydrocarbon degradation | 165 | 32 | 100 | 3 | 19 | 3 | 27 | 19 | 5 | 13 | 15 | 32 | |
| Chloroplasts | 6903 | 8815 | 23,691 | 1075 | 2393 | 4309 | 2608 | 3667 | 2613 | 2597 | 1565 | 1837 | |
| Chemoheterotrophy | 21,276 | 7079 | 12,891 | 13,907 | 7614 | 7437 | 8470 | 7073 | 5295 | 4728 | 3203 | 2546 | |
| Sulfur cycle | Sulfate respiration | 220 | 519 | 95 | 16 | 31 | 17 | 51 | 23 | 6 | 110 | 104 | 22 |
| Sulfur respiration | 310 | 49 | 94 | 15 | 44 | 25 | 39 | 12 | 2 | 8 | 13 | 26 | |
| Sulfite respiration | 5 | 123 | 19 | 0 | 21 | 4 | 0 | 4 | 1 | 1 | 2 | 1 | |
| Thiosulfate respiration | 0 | 2 | 0 | 4 | 22 | 10 | 19 | 0 | 0 | 3 | 0 | 0 | |
| Respiration of sulfur compounds | 530 | 568 | 189 | 31 | 75 | 42 | 90 | 35 | 8 | 118 | 117 | 48 | |
| Dark oxidation of sulfur compounds | 12 | 0 | 18 | 0 | 6 | 0 | 5 | 3 | 4 | 5 | 1 | 0 | |
| Others | Human pathogens: all | 336 | 179 | 158 | 9628 | 208 | 111 | 150 | 36 | 20 | 1213 | 37 | 25 |
| Dark hydrogen oxidation | 56 | 25 | 55 | 33 | 41 | 36 | 58 | 42 | 12 | 72 | 43 | 58 | |
| Iron respiration | 341 | 116 | 96 | 13 | 43 | 16 | 27 | 13 | 3 | 9 | 19 | 45 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Q.; Zeng, L.; Li, C.; Ye, C. The Effects of Varying Concentrations of Didecyl Methylpropyl Ammonium Iodide (DMPAI) on the Structure and Function of Soil Bacterial Communities in the Lake–Terrestrial Ecotone. Microorganisms 2025, 13, 934. https://doi.org/10.3390/microorganisms13040934
Zhu Q, Zeng L, Li C, Ye C. The Effects of Varying Concentrations of Didecyl Methylpropyl Ammonium Iodide (DMPAI) on the Structure and Function of Soil Bacterial Communities in the Lake–Terrestrial Ecotone. Microorganisms. 2025; 13(4):934. https://doi.org/10.3390/microorganisms13040934
Chicago/Turabian StyleZhu, Qi, Lingquan Zeng, Chunhua Li, and Chun Ye. 2025. "The Effects of Varying Concentrations of Didecyl Methylpropyl Ammonium Iodide (DMPAI) on the Structure and Function of Soil Bacterial Communities in the Lake–Terrestrial Ecotone" Microorganisms 13, no. 4: 934. https://doi.org/10.3390/microorganisms13040934
APA StyleZhu, Q., Zeng, L., Li, C., & Ye, C. (2025). The Effects of Varying Concentrations of Didecyl Methylpropyl Ammonium Iodide (DMPAI) on the Structure and Function of Soil Bacterial Communities in the Lake–Terrestrial Ecotone. Microorganisms, 13(4), 934. https://doi.org/10.3390/microorganisms13040934