
Impacts of Tick Parasitism on the Rodent Gut Microbiome

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Molecular Methods
2.2. Informatic and Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mackowiak, P.A. The normal microbial flora. N. Engl. J. Med. 1982, 307, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Bello, M.G.; Godoy-Vitorino, F.; Knight, R.; Blaser, M.J. Role of the microbiome in human development. Gut 2019, 68, 1108–1114. [Google Scholar] [CrossRef] [PubMed]
- Tremlett, H.; Bauer, K.C.; Appel-Cresswell, S.; Finlay, B.B.; Waubant, E. The gut microbiome in human neurological disease: A review. Ann. Neurol. 2017, 81, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Dinan, T.G.; Cryan, J.F. The microbiome-gut-brain axis in health and disease. Gastroenterol. Clin. 2017, 46, 77–89. [Google Scholar] [CrossRef]
- Flux, M.C.; Lowry, C.A. Finding intestinal fortitude: Integrating the microbiome into a holistic view of depression mechanisms, treatment, and resilience. Neurobiol. Dis. 2020, 135, 104578. [Google Scholar] [CrossRef]
- Wehkamp, J.; Frick, J.S. Microbiome and chronic inflammatory bowel diseases. J. Mol. Med. 2017, 95, 21–28. [Google Scholar] [CrossRef]
- Lleal, M.; Sarrabayrouse, G.; Willamil, J.; Santiago, A.; Pozuelo, M.; Manichanh, C. A single faecal microbiota transplantation modulates the microbiome and improves clinical manifestations in a rat model of colitis. eBioMedicine 2019, 48, 630–641. [Google Scholar] [CrossRef]
- Winter, G.; Hart, R.A.; Charlesworth, R.P.; Sharpley, C.F. Gut microbiome and depression: What we know and what we need to know. Rev. Neurosci. 2018, 29, 629–643. [Google Scholar] [CrossRef]
- Perry, R.J.; Peng, L.; Barry, N.A.; Cline, G.W.; Zhang, D.; Cardone, R.L.; Petersen, K.F.; Kibbey, R.G.; Goodman, A.L.; Shulman, G.I. Acetate mediates a microbiome–brain–β-cell axis to promote metabolic syndrome. Nature 2016, 534, 213–217. [Google Scholar] [CrossRef]
- Pilla, R.; Suchodolski, J.S. The role of the canine gut microbiome and metabolome in health and gastrointestinal disease. Front. Vet. Sci. 2020, 6, 498. [Google Scholar] [CrossRef]
- Libertucci, J.; Young, V.B. The role of the microbiota in infectious diseases. Nat. Microbiol. 2019, 4, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef]
- Yeoh, Y.K.; Zuo, T.; Lui, G.C.; Zhang, F.; Liu, Q.; Li, A.Y.; Chung, A.C.; Cheung, C.P.; Tso, E.Y.; Fung, K.S.; et al. Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID-19. Gut 2021, 70, 698–706. [Google Scholar] [CrossRef]
- Morrissette, M.; Pitt, N.; González, A.; Strandwitz, P.; Caboni, M.; Rebman, A.W.; Knight, R.; D’onofrio, A.; Aucott, J.N.; Soloski, M.J.; et al. A distinct microbiome signature in posttreatment Lyme disease patients. mBio 2020, 11, e02310-20. [Google Scholar] [CrossRef]
- Bonnet, S.I.; Binetruy, F.; Hernández-Jarguín, A.M.; Duron, O. The tick microbiome: Why non-pathogenic microorganisms matter in tick biology and pathogen transmission. Front. Cell. Infect. Microbiol. 2017, 7, 236. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente, J.; Villar, M.; Cabezas-Cruz, A.; Estrada-Peña, A.; Ayllon, N.; Alberdi, P. Tick–Host–Pathogen Interactions: Conflict and Cooperation. PLoS Pathog. 2016, 12, e1005488. [Google Scholar] [CrossRef] [PubMed]
- Cabezas-Cruz, A.; Estrada-Peña, A.; Rego, R.O.; De la Fuente, J. Tick-pathogen ensembles: Do molecular interactions lead ecological innovation? Front. Cell. Infect. Microbiol. 2017, 7, 74. [Google Scholar] [CrossRef] [PubMed]
- Faraone, N.; Light, M.; Scott, C.; MacPherson, S.; Hillier, N.K. Chemosensory and behavioural responses of Ixodes scapularis to natural products: Role of chemosensory organs in volatile detection. Insects 2020, 11, 502. [Google Scholar] [CrossRef] [PubMed]
- Lutz, H.L.; Gilbert, J.A.; Dick, C.W. Associations between Afrotropical bats, eukaryotic parasites, and microbial symbionts. Mol. Ecol. 2022, 31, 1939–1950. [Google Scholar] [CrossRef]
- Roodt, A.P.; Naudé, Y.; Stoltz, A.; Rohwer, E. Human skin volatiles: Passive sampling and GC× GC-ToFMS analysis as a tool to investigate the skin microbiome and interactions with anthropophilic mosquito disease vectors. J. Chromatogr. B 2018, 1097, 83–93. [Google Scholar] [CrossRef]
- Mann, E.A.; Bae, E.; Kostyuchek, D.; Chung, H.J.; McGee, J.S. The gut microbiome: Human health and inflammatory skin diseases. Ann. Dermatol. 2020, 32, 265. [Google Scholar] [CrossRef]
- Pessemier, B.D.; Grine, L.; Debaere, M.; Maes, A.; Paetzold, B.; Callewaert, C. Gut–skin axis: Current knowledge of the interrelationship between microbial dysbiosis and skin conditions. Microorganisms 2021, 9, 353. [Google Scholar] [CrossRef] [PubMed]
- Salem, I.; Ramser, A.; Isham, N.; Ghannoum, M.A. The gut microbiome as a major regulator of the gut-skin axis. Front. Microbiol. 2018, 9, 1459. [Google Scholar] [CrossRef] [PubMed]
- Frischknecht, F. The skin as interface in the transmission of arthropod-borne pathogens. Cell. Microbiol. 2007, 9, 1630–1640. [Google Scholar] [CrossRef]
- Fritzsche, A.; Allan, B.F. The ecology of fear: Host foraging behavior varies with the spatio-temporal abundance of a dominant ectoparasite. EcoHealth 2012, 9, 70–74. [Google Scholar] [CrossRef] [PubMed]
- André, A.; Mouton, A.; Millien, V.; Michaux, J. Liver microbiome of Peromyscus leucopus, a key reservoir host species for emerging infectious diseases in North America. Infect. Genet. Evol. 2017, 52, 10–18. [Google Scholar] [CrossRef]
- Hahn, M.A.; Piecyk, A.; Jorge, F.; Cerrato, R.; Kalbe, M.; Dheilly, N.M. Host phenotype and microbiome vary with infection status, parasite genotype, and parasite microbiome composition. Mol. Ecol. 2022, 31, 1577–1594. [Google Scholar] [CrossRef]
- Zaiss, M.M.; Harris, N.L. Interactions between the intestinal microbiome and helminth parasites. Parasite Immunol. 2016, 38, 5–11. [Google Scholar] [CrossRef]
- Bunikis, J.; Tsao, J.; Luke, C.J.; Luna, M.G.; Fish, D.; Barbour, A.G. Borrelia burgdorferi infection in a natural population of Peromyscus leucopus mice: A longitudinal study in an area where Lyme borreliosis is highly endemic. J. Infect. Dis. 2004, 189, 1515–1523. [Google Scholar] [CrossRef]
- Moraru, G.M.; Goddard, J.; Paddock, C.D.; Varela-Stokes, A. Experimental infection of cotton rats and bobwhite quail with Rickettsia parkeri. Parasites Vectors 2013, 6, 70. [Google Scholar] [CrossRef]
- Cumbie, A.N.; Espada, C.D.; Nadolny, R.M.; Rose, R.K.; Dueser, R.D.; Hynes, W.L.; Gaff, H.D. Survey of Rickettsia parkeri and Amblyomma maculatum associated with small mammals in southeastern Virginia. Ticks Tick-Borne Dis. 2020, 11, 101550. [Google Scholar] [CrossRef] [PubMed]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef]
- Apprill, A.; McNally, S.; Parsons, R.; Weber, L. Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecol. 2015, 75, 129–137. [Google Scholar] [CrossRef]
- Illumina. 16s Metagenomic Sequencing Library Preparation; Illumina: San Diego, CA, USA, 2013; p. 21. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Kaehler, B.D.; Bokulich, N.A.; McDonald, D.; Knight, R.; Caporaso, J.G.; Huttley, G.A. Species abundance information improves sequence taxonomy classification accuracy. Nat. Commun. 2019, 10, 4643. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H.; et al. Package ‘vegan’, version 2.6-10; Community Ecology Package; 2013; Volume 2, pp. 1–295. Available online: https://CRAN.R-project.org/package=vegan (accessed on 7 April 2025).
- Lin, H.; Peddada, S.D. Analysis of compositions of microbiomes with bias correction. Nat. Commun. 2020, 11, 3514. [Google Scholar] [CrossRef]
- Baxter, N.T.; Wan, J.J.; Schubert, A.M.; Jenior, M.L.; Myers, P.; Schloss, P.D. Intra-and interindividual variations mask interspecies variation in the microbiota of sympatric Peromyscus populations. Appl. Environ. Microbiol. 2015, 81, 396–404. [Google Scholar] [CrossRef]
- Menzel, M.A.; Carter, T.C.; Houston, A.T.; Longe, R.L. Notes on the microhabitat associations of the hispid cotton rat (Sigmodon hispidus) and white-footed mouse (Peromyscus leucopus) in the piedmont physiographic province. Ga. J. Sci. 1999, 57, 180. [Google Scholar]
- Lackey, J.A.; Huckaby, D.G.; Ormiston, B.G. Peromyscus leucopus. Mamm. Species 1985, 247, 1. [Google Scholar] [CrossRef]
- Randolph, J.C.; Cameron, G.N.; Wrazen, J.A. Dietary choice of a generalist grassland herbivore, Sigmodon hispidus. J. Mammal. 1991, 72, 300–313. [Google Scholar] [CrossRef]
- Anders, J.L.; Moustafa, M.A.; Mohamed, W.M.; Hayakawa, T.; Nakao, R.; Koizumi, I. Comparing the gut microbiome along the gastrointestinal tract of three sympatric species of wild rodents. Sci. Rep. 2021, 11, 19929. [Google Scholar] [CrossRef]
- Bowerman, K.L.; Knowles, S.C.; Bradley, J.E.; Baltrūnaitė, L.; Lynch, M.D.; Jones, K.M.; Hugenholtz, P. Effects of laboratory domestication on the rodent gut microbiome. ISME Commun. 2021, 1, 49. [Google Scholar] [CrossRef]
- Kohl, K.D.; Dieppa-Colón, E.; Goyco-Blas, J.; Peralta-Martínez, K.; Scafidi, L.; Shah, S.; Zawacki, E.; Barts, N.; Ahn, Y.; Hedayati, S.; et al. Gut microbial ecology of five species of sympatric desert rodents in relation to herbivorous and insectivorous feeding strategies. Integr. Comp. Biol. 2022, 62, 237–251. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, C.A.; Monteleone, G.; McLaughlin, J.T.; Paus, R. The gut-skin axis in health and disease: A paradigm with therapeutic implications. Bioessays 2016, 38, 1167–1176. [Google Scholar] [CrossRef]
- Sinha, S.; Lin, G.; Ferenczi, K. The skin microbiome and the gut-skin axis. Clin. Dermatol. 2021, 39, 829–839. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, M. Skin barrier function and the microbiome. Int. J. Mol. Sci. 2022, 23, 13071. [Google Scholar] [CrossRef]
- van Splunter, M.; Liu, L.; van Neerven, R.J.; Wichers, H.J.; Hettinga, K.A.; De Jong, N.W. Mechanisms underlying the skin-gut cross talk in the development of IgE-mediated food allergy. Nutrients 2020, 12, 3830. [Google Scholar] [CrossRef] [PubMed]
- Bosman, E.S.; Albert, A.Y.; Lui, H.; Dutz, J.P.; Vallance, B.A. Skin exposure to narrow band ultraviolet (UVB) light modulates the human intestinal microbiome. Front. Microbiol. 2019, 10, 477346. [Google Scholar] [CrossRef]
- Wikel, S.K.; Ramachandra, R.N.; Bergman, D.K. Tick-induced modulation of the host immune response. Int. J. Parasitol. 1994, 24, 59–66. [Google Scholar] [CrossRef]
- Brossard, M.; Wikel, S. Tick Immunobiology. In Ticks: Biology, Disease, and Control; Bowmand, A.S., Nuttall, P.A., Eds.; Cambridge University Press: Cambridge, UK, 2004; pp. 186–204. [Google Scholar]
- Skallová, A.; Iezzi, G.; Ampenberger, F.; Kopf, M.; Kopecký, J. Tick saliva inhibits dendritic cell migration, maturation, and function while promoting development of Th2 responses. J. Immunol. 2008, 180, 6186–6192. [Google Scholar] [CrossRef] [PubMed]
- Kotál, J.; Langhansová, H.; Lieskovská, J.; Andersen, J.F.; Francischetti, I.M.; Chavakis, T.; Kopecký, J.; Pedra, J.H.; Kotsyfakis, M.; Chmelař, J. Modulation of host immunity by tick saliva. J. Proteom. 2015, 128, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Kovář, L.; Kopecký, J.; Říhová, B. Salivary gland extract from Ixodes ricinus tick modulates the host immune response towards the Th2 cytokine profile. Parasitol. Res. 2002, 88, 1066–1072. [Google Scholar] [CrossRef]
- Sang, H.; Xie, Y.; Su, X.; Zhang, M.; Zhang, Y.; Liu, K.; Wang, J. Mushroom Bulgaria inquinans modulates host immunological response and gut microbiota in mice. Front. Nutr. 2020, 7, 144. [Google Scholar] [CrossRef]
- Chung, Y.W.; Gwak, H.J.; Moon, S.; Rho, M.; Ryu, J.H. Functional dynamics of bacterial species in the mouse gut microbiome revealed by metagenomic and metatranscriptomic analyses. PLoS ONE 2020, 15, e0227886. [Google Scholar] [CrossRef]
- Zhu, Y.; Chen, B.; Zhang, X.; Akbar, M.T.; Wu, T.; Zhang, Y.; Zhi, L.; Shen, Q. Exploration of the Muribaculaceae family in the gut microbiota: Diversity, metabolism, and function. Nutrients 2024, 16, 2660. [Google Scholar] [CrossRef]
- Pereira, F.C.; Wasmund, K.; Cobankovic, I.; Jehmlich, N.; Herbold, C.W.; Lee, K.S.; Sziranyi, B.; Vesely, C.; Decker, T.; Stocker, R.; et al. Rational design of a microbial consortium of mucosal sugar utilizers reduces Clostridiodes difficile colonization. Nat. Commun. 2020, 11, 5104. [Google Scholar] [CrossRef]
- Dabbagh, K.; Takeyama, K.; Lee, H.M.; Ueki, I.F.; Lausier, J.A.; Nadel, J.A. IL-4 induces mucin gene expression and goblet cell metaplasia in vitro and in vivo. J. Immunol. 1999, 162, 6233–6237. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.J.; Miller, R.A.; Ericsson, A.C.; Harrison, D.C.; Strong, R.; Schmidt, T.M. Changes in the gut microbiome and fermentation products concurrent with enhanced longevity in acarbose-treated mice. BMC Microbiol. 2019, 19, 130. [Google Scholar] [CrossRef]
- Ansaldo, E.; Slayden, L.C.; Ching, K.L.; Koch, M.A.; Wolf, N.K.; Plichta, D.R.; Brown, E.M.; Graham, D.B.; Xavier, R.J.; Moon, J.J.; et al. Akkermansia muciniphila induces intestinal adaptive immune responses during homeostasis. Science 2019, 364, 1179–1184. [Google Scholar] [CrossRef]
- Kreisinger, J.; Bastien, G.; Hauffe, H.C.; Marchesi, J.; Perkins, S.E. Interactions between multiple helminths and the gut microbiota in wild rodents. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140295. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-López, M.J. Mosquito behavior and vertebrate microbiota interaction: Implications for pathogen transmission. Front. Microbiol. 2020, 11, 573371. [Google Scholar] [CrossRef] [PubMed]
- Cozzarolo, C.S.; Glaizot, O.; Christe, P.; Pigeault, R. Enhanced attraction of arthropod vectors to infected vertebrates: A review of empirical evidence. Front. Ecol. Evol. 2020, 8, 296. [Google Scholar] [CrossRef]
- Franzin, A.M.; Maruyama, S.R.; Garcia, G.R.; Oliveira, R.P.; Ribeiro, J.M.; Bishop, R.; Maia, A.A.; Moré, D.D.; Ferreira, B.R.; de Miranda Santos, I.K. Immune and biochemical responses in skin differ between bovine hosts genetically susceptible and resistant to the cattle tick Rhipicephalus microplus. Parasites Vectors 2017, 10, 51. [Google Scholar] [CrossRef]
- Sherwin, E.; Bordenstein, S.R.; Quinn, J.L.; Dinan, T.G.; Cryan, J.F. Microbiota and the social brain. Science 2019, 366, eaar2016. [Google Scholar] [CrossRef]
- Sih, A.; Spiegel, O.; Godfrey, S.; Leu, S.; Bull, C.M. Integrating social networks, animal personalities, movement ecology and parasites: A framework with examples from a lizard. Anim. Behav. 2018, 136, 195–205. [Google Scholar] [CrossRef]
- Knutie, S.A. Food supplementation affects gut microbiota and immunological resistance to parasites in a wild bird species. J. Appl. Ecol. 2020, 57, 536–547. [Google Scholar] [CrossRef]
- Anderson, J.M.; Moore, I.N.; Nagata, B.M.; Ribeiro, J.M.; Valenzuela, J.G.; Sonenshine, D.E. Ticks, Ixodes scapularis, feed repeatedly on white-footed mice despite strong inflammatory response: An expanding paradigm for understanding tick–host interactions. Front. Immunol. 2017, 8, 1784. [Google Scholar] [CrossRef]
- Brunner, J.L.; Ostfeld, R.S. Multiple causes of variable tick burdens on small-mammal hosts. Ecology 2008, 89, 2259–2272. [Google Scholar] [CrossRef] [PubMed]
- Brehm, A.M.; Assis, V.R.; Martin, L.B.; Orrock, J.L. Individual variation underlies large-scale patterns: Host conditions and behavior affect parasitism. Ecology 2024, 106, e4478. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Ju, F. Evaluation of different 16S rRNA gene hypervariable regions and reference databases for profiling engineered microbiota structure and functional guilds in a swine wastewater treatment plant. Interface Focus 2023, 13, 20230012. [Google Scholar] [CrossRef]
- López-Aladid, R.; Fernández-Barat, L.; Alcaraz-Serrano, V.; Bueno-Freire, L.; Vázquez, N.; Pastor-Ibáñez, R.; Palomeque, A.; Oscanoa, P.; Torres, A. Determining the most accurate 16S rRNA hypervariable region for taxonomic identification from respiratory samples. Sci. Rep. 2023, 13, 3974. [Google Scholar] [CrossRef]
- Clooney, A.G.; Fouhy, F.; Sleator, R.D.; O’Driscoll, A.; Stanton, C.; Cotter, P.D.; Claesson, M.J. Comparing apples and oranges?: Next generation sequencing and its impact on microbiome analysis. PLoS ONE 2016, 11, e0148028. [Google Scholar] [CrossRef] [PubMed]
- Salamon, D.; Zapała, B.; Krawczyk, A.; Potasiewicz, A.; Nikiforuk, A.; Stój, A.; Gosiewski, T. Comparison of iSeq and MiSeq as the two platforms for 16S rRNA sequencing in the study of the gut of rat microbiome. Appl. Microbiol. Biotechnol. 2022, 106, 7671–7681. [Google Scholar] [CrossRef]
- Kim, J.Y.; Yi, M.H.; Mahdi, A.A.; Yong, T.S. iSeq 100 for metagenomic pathogen screening in ticks. Parasites Vectors 2021, 14, 346. [Google Scholar] [CrossRef]
- Illumina. 16S Metagenomics Sequencing with the iSeq 100 System; Illumina: San Diego, CA, USA, 2018; p. 4. [Google Scholar]
- Jupatanakul, N.; Sim, S.; Dimopoulos, G. The insect microbiome modulates vector competence for arboviruses. Viruses 2014, 6, 4294–4313. [Google Scholar] [CrossRef]
- Ring, K.; Couper, L.I.; Sapiro, A.L.; Yarza, F.; Yang, X.F.; Clay, K.; Mateusiak, C.; Chou, S.; Swei, A. Host blood meal identity modifies vector gene expression and competency. Mol. Ecol. 2022, 31, 2698–2711. [Google Scholar] [CrossRef]
- Wikel, S.K. Tick-host interactions. Biol. Ticks 2014, 2, 88–128. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
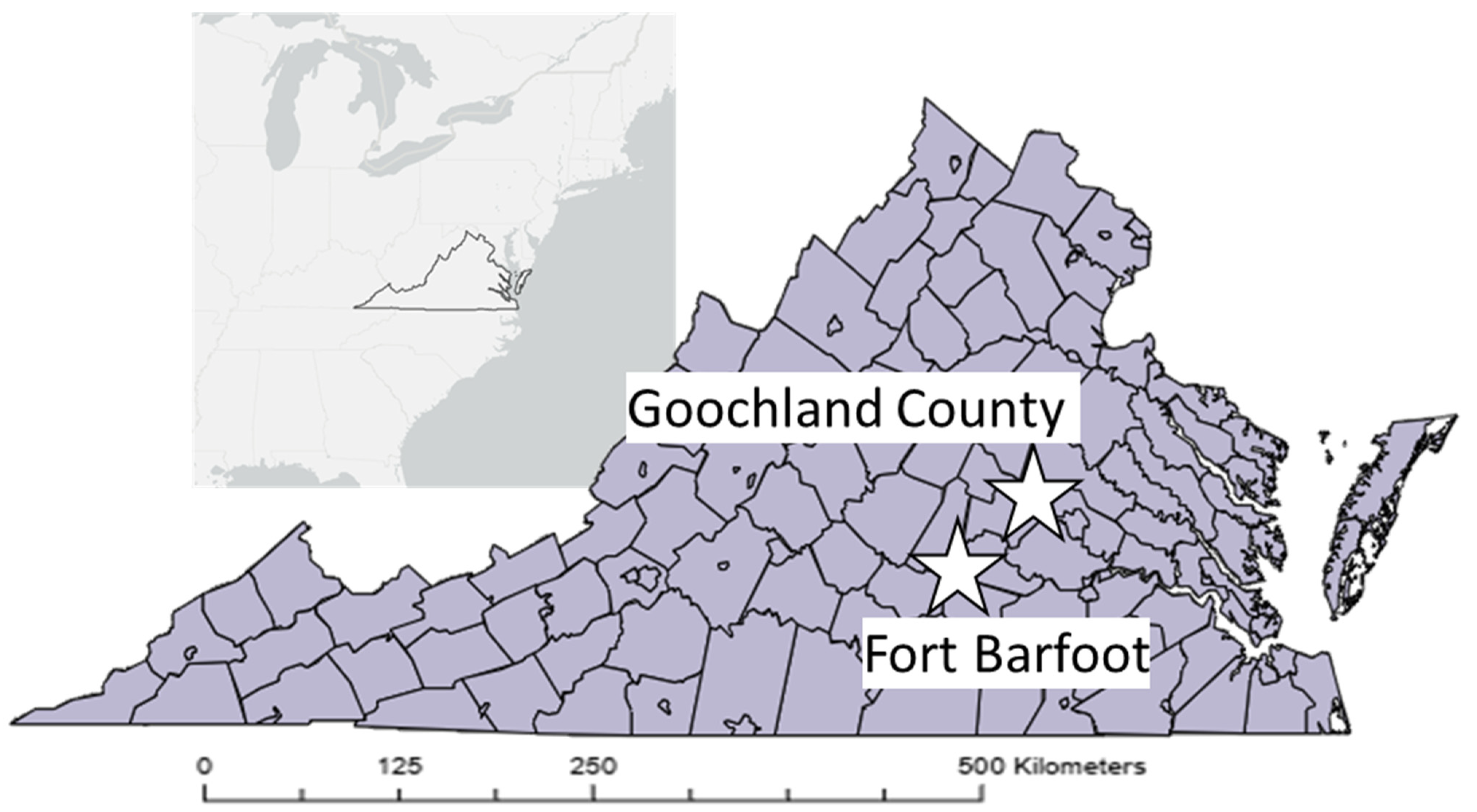
| Sampling Location | Rodent Species | Individuals Sampled | Individuals with Ticks |
|---|---|---|---|
| Goochland | Peromyscus leucopus | 24 | 9 |
| Fort Barfoot | Peromyscus leucopus | 14 | 3 |
| Sigmodon hispidus | 7 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brinkerhoff, R.J.; Pandian, J.; Leber, M.; Hauser, I.D.; Gaff, H.D. Impacts of Tick Parasitism on the Rodent Gut Microbiome. Microorganisms 2025, 13, 888. https://doi.org/10.3390/microorganisms13040888
Brinkerhoff RJ, Pandian J, Leber M, Hauser ID, Gaff HD. Impacts of Tick Parasitism on the Rodent Gut Microbiome. Microorganisms. 2025; 13(4):888. https://doi.org/10.3390/microorganisms13040888
Chicago/Turabian StyleBrinkerhoff, Robert Jory, Joshua Pandian, Meghan Leber, Isabella D. Hauser, and Holly D. Gaff. 2025. "Impacts of Tick Parasitism on the Rodent Gut Microbiome" Microorganisms 13, no. 4: 888. https://doi.org/10.3390/microorganisms13040888
APA StyleBrinkerhoff, R. J., Pandian, J., Leber, M., Hauser, I. D., & Gaff, H. D. (2025). Impacts of Tick Parasitism on the Rodent Gut Microbiome. Microorganisms, 13(4), 888. https://doi.org/10.3390/microorganisms13040888