
Cas5 Regulates the Exposure of β-Glucan, the Cell Surface Hydrophobicity, and the Expression of Cell Wall Proteins to Remodel the Candida albicans Cell Wall and Participates in the Recruitment of Neutrophils


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.1.1. Type of Study
2.1.2. Key Variables
- (1)
- Independent Variables
- (a)
- Cas5 Depletion: presence or absence of the CAS5 gene (wild-type C. albicans SC5314 [WT] vs. Cas5 null mutant [cas5Δ/Δ]).
- (2)
- Dependent Variables
- (a)
- Cell Wall Structure: changes in C. albicans cell wall glycans exposures and CWPs expressions;
- (b)
- Adhesion to Host Cells: assessment of the ability of C. albicans to adhere to host endothelial and epithelial cells;
- (c)
- Morphology: assessment of the ability of hypha and biofilm formation;
- (d)
- Innate Immune Cells Recruitment: quantification of neutrophils and macrophages infiltration in the tongue tissues of BALB/C male mice with OPC.
2.1.3. Sources of Variation
- (1)
- Strain Variability
- (a)
- Genetic Background: both WT and cas5Δ/Δ strains were derived from the same parental strain (SC5314) to minimize genetic background differences;
- (b)
- Knockout Efficiency: the efficiency of the CAS5 gene knockout was confirmed using PCR and sequencing to ensure consistent genetic modification across all experimental samples;
- (c)
- Complementation Efficiency: the efficiency of the CAS5 gene complementation was confirmed using RT-qPCR to ensure consistent genetic modification across all experimental samples.
- (2)
- Culture Conditions
- (a)
- Medium: all cultures were grown in YPD medium at 30 °C or RPMI 1640 medium containing 10% FBS with constant shaking (220 rpm) to maintain consistent growth conditions;
- (b)
- Growth Phase: experiments were conducted using C. albicans cells harvested at late-log phase to ensure uniformity in metabolic activity and physiological state.
- (3)
- Mouse Model Variability
- (a)
- All the BALB/C mice used in this study were male, aged 6–8 weeks, and maintained in identical baseline health status.
- (4)
- Random Error
- (a)
- Replicate Measurements: each experiment was repeated at least three times, and the average value was taken as the final result to reduce the impact of random errors.
2.1.4. Study Design Overview
- (1)
- Strain Construction
- (a)
- Generation of C. albicans strains with and without Cas5 depletion using CRISPR/Cas9 homologous recombination technology.
- (2)
- In Vitro Experiments
- (a)
- Cell Wall Analysis: use transmission electron microscopy, flow cytometry, fluorescence microscopy, TMT-labeled proteomics, and CSH assay to evaluate changes in cell wall structure induced by Cas5 depletion;
- (b)
- Adhesion Assays: conduct adhesion assays using typical endothelial and epithelial host cells to measure the adhesion capacity of various C. albicans strains;
- (c)
- Morphological Transition Assessment: employ fluorescence microscopy, scanning electron microscopy, and biofilm formation assay to examine morphology transition of various C. albicans strains.
- (3)
- In Vitro Experiments
- (a)
- Mouse Model Establishment: infect BALB/C mice with various C. albicans strains via oral inoculation to establish OPC;
- (b)
- Innate Immune Cell Recruitment: collect tongue tissues at first day post-infection and quantify immune cell infiltration using immunofluorescence staining;
- (c)
- C. albicans pathogenicity: collect tongue tissues at the fifth day post-infection and evaluate the pathogenicity of various C. albicans strains using histopathologic examination, immunohistochemistry (IHC), and fungal burden.
- (4)
- Statistical Analysis
2.2. Plasmid and Strain Constructions
2.3. Medium and Growth Conditions
2.4. Transmission Electron Microscopy (TEM) of Cell Wall Structure
2.5. Cell Wall Glycans Exposures
2.6. Expression of Cell Wall Proteins (CWPs)
2.7. CSH Assay
2.8. Adhesion on Host Cells
2.9. Fluorescence Microscopy of C. albicans Hypha Growth
2.10. Scanning Electron Microscopy (SEM) of C. albicans Hypha Growth
2.11. Biofilm Formation Assay
2.12. Mouse Model of C. albicans Infection
2.13. Neutrophil and Macrophage Recruitment Assay
2.14. Histopathologic Examination, IHC, and Fungal Burden
3. Results
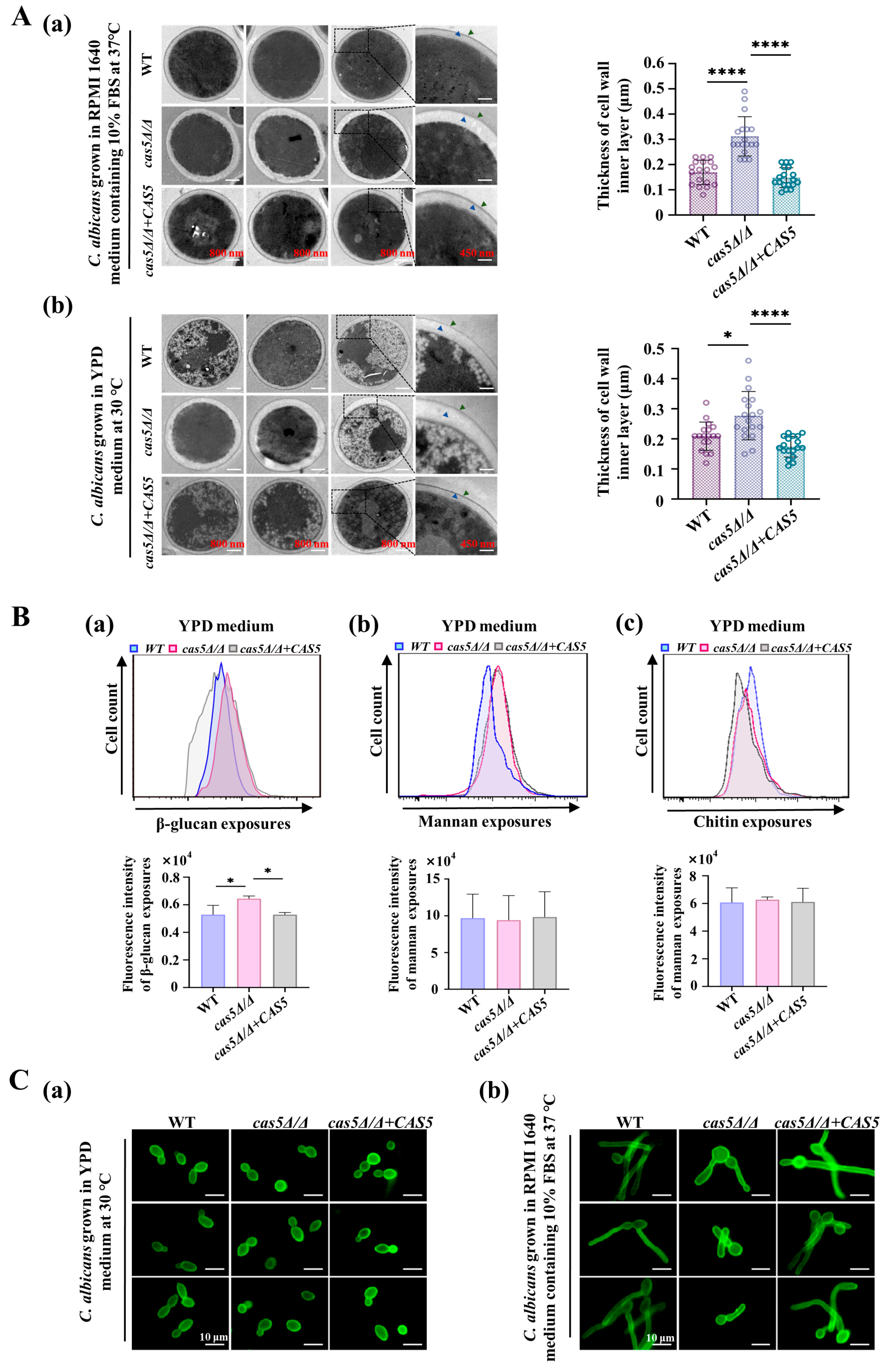
3.1. Cas5 Helps Cell Wall β-Glucans Mask
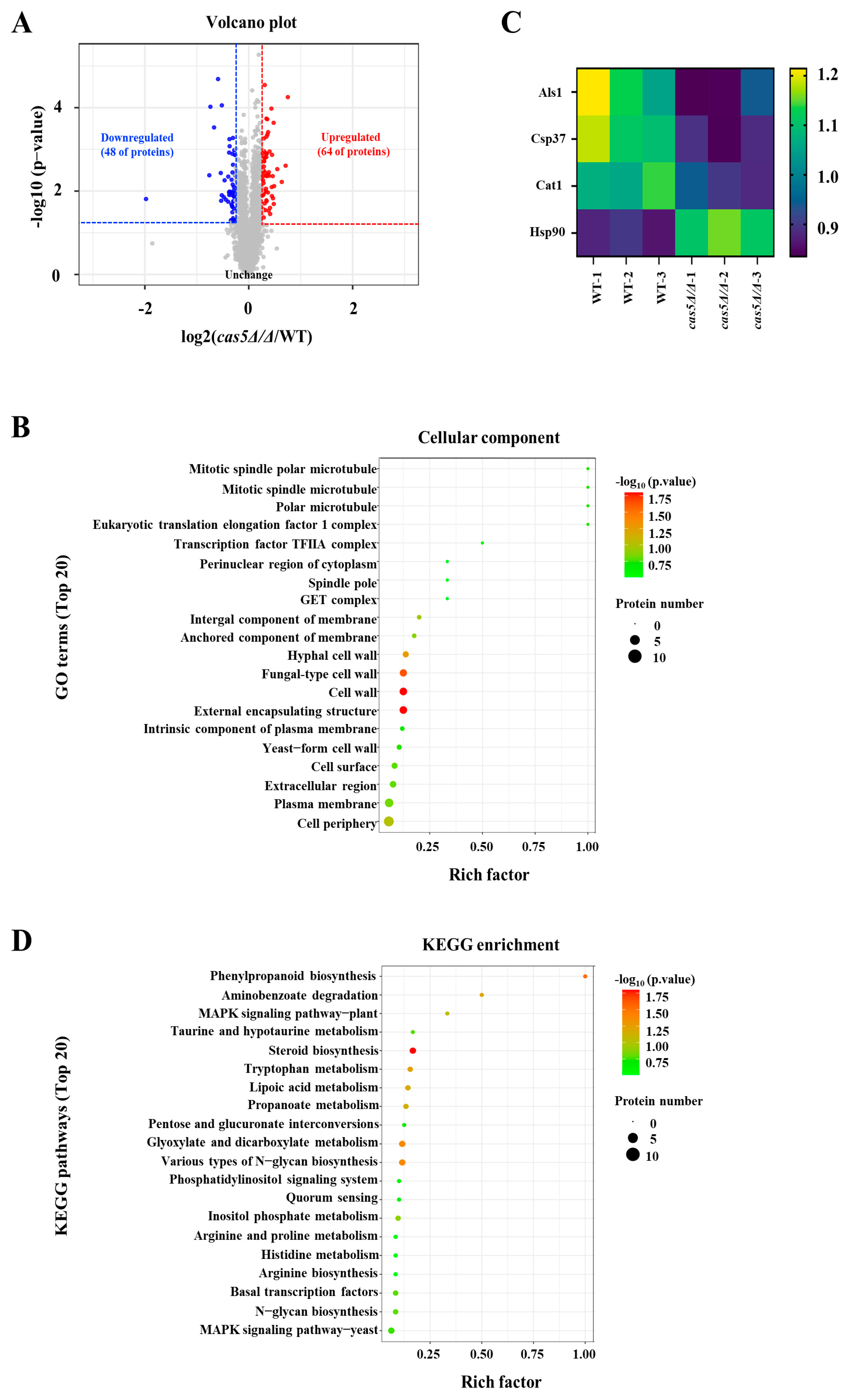
3.2. Cas5 Regulates CWP Expression
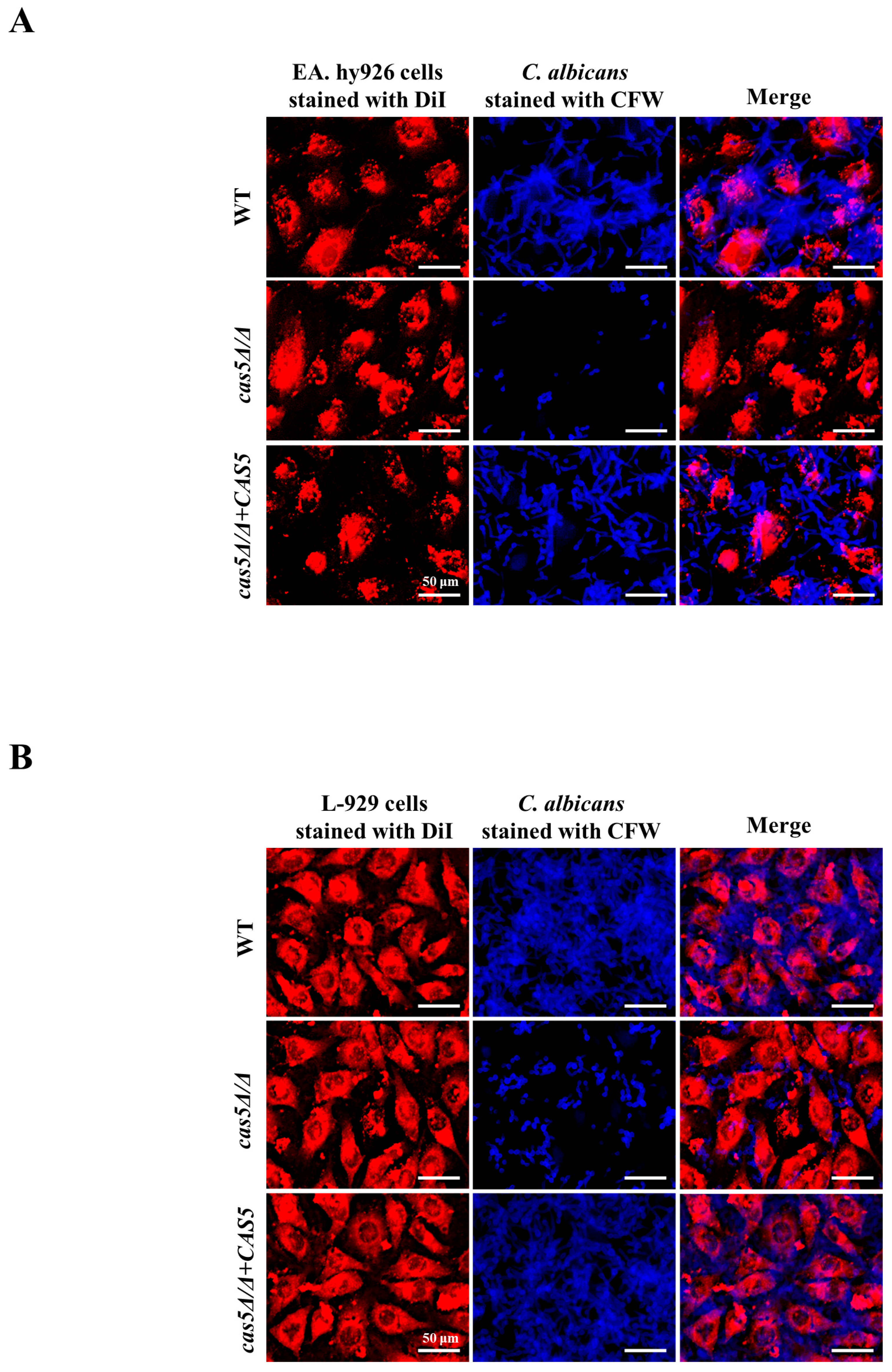
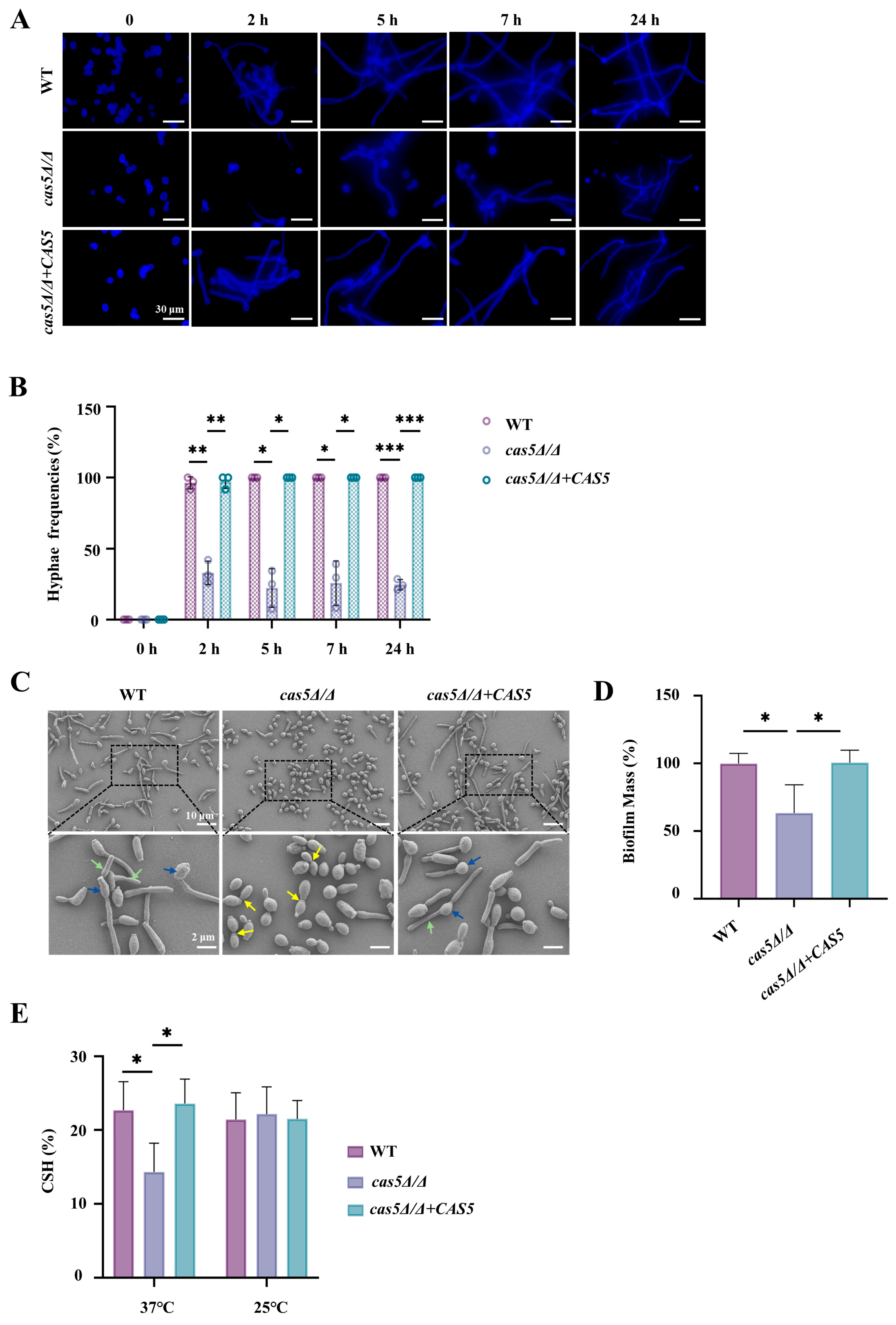
3.3. Cas5 Regulates C. albicans Adhesion to Host Cells, Morphology Transition, and Cell Surface Hydrophobicity (CSH)
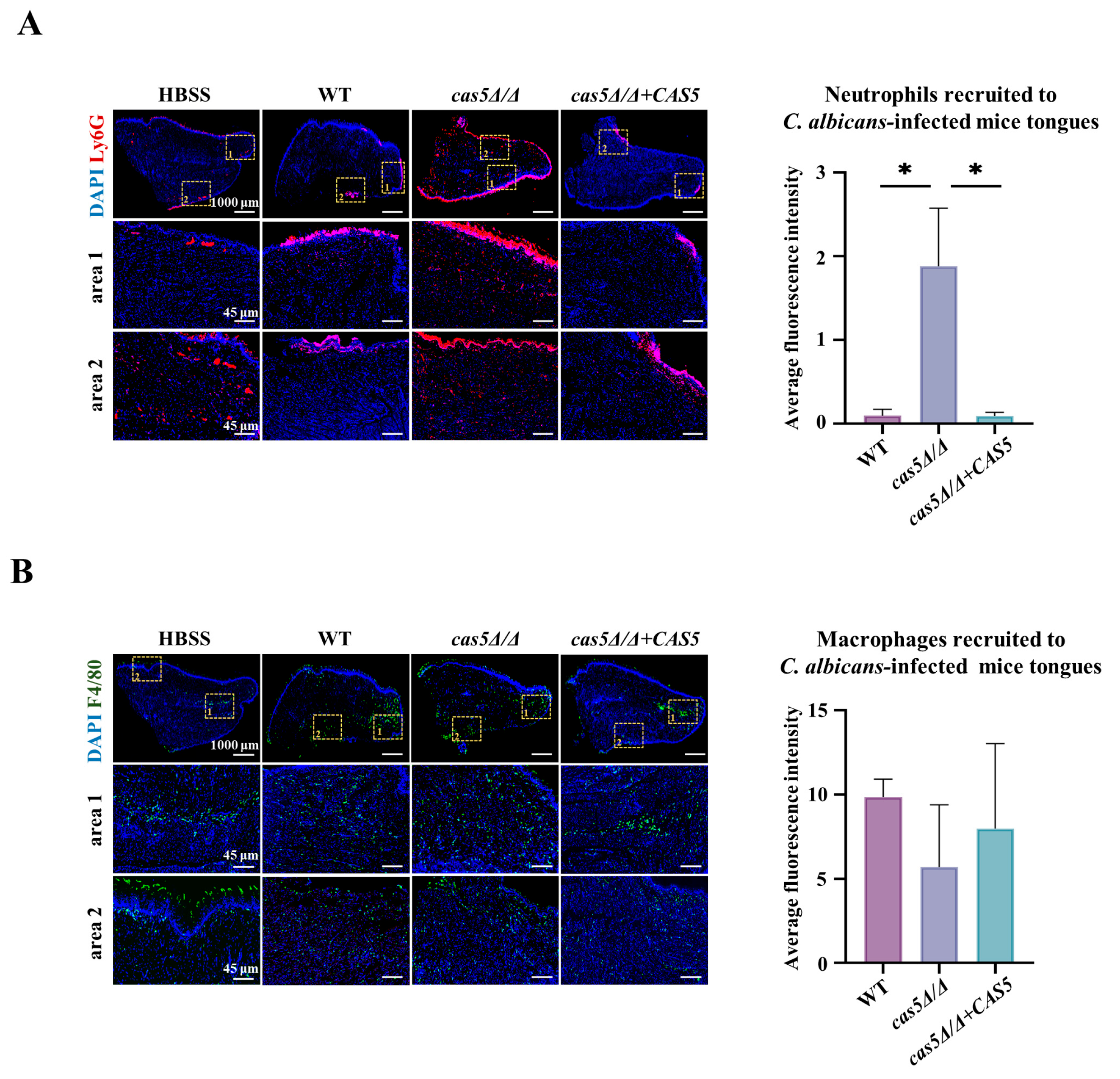
3.4. Cas5 Knockout Results in Increased Neutrophil Recruitment
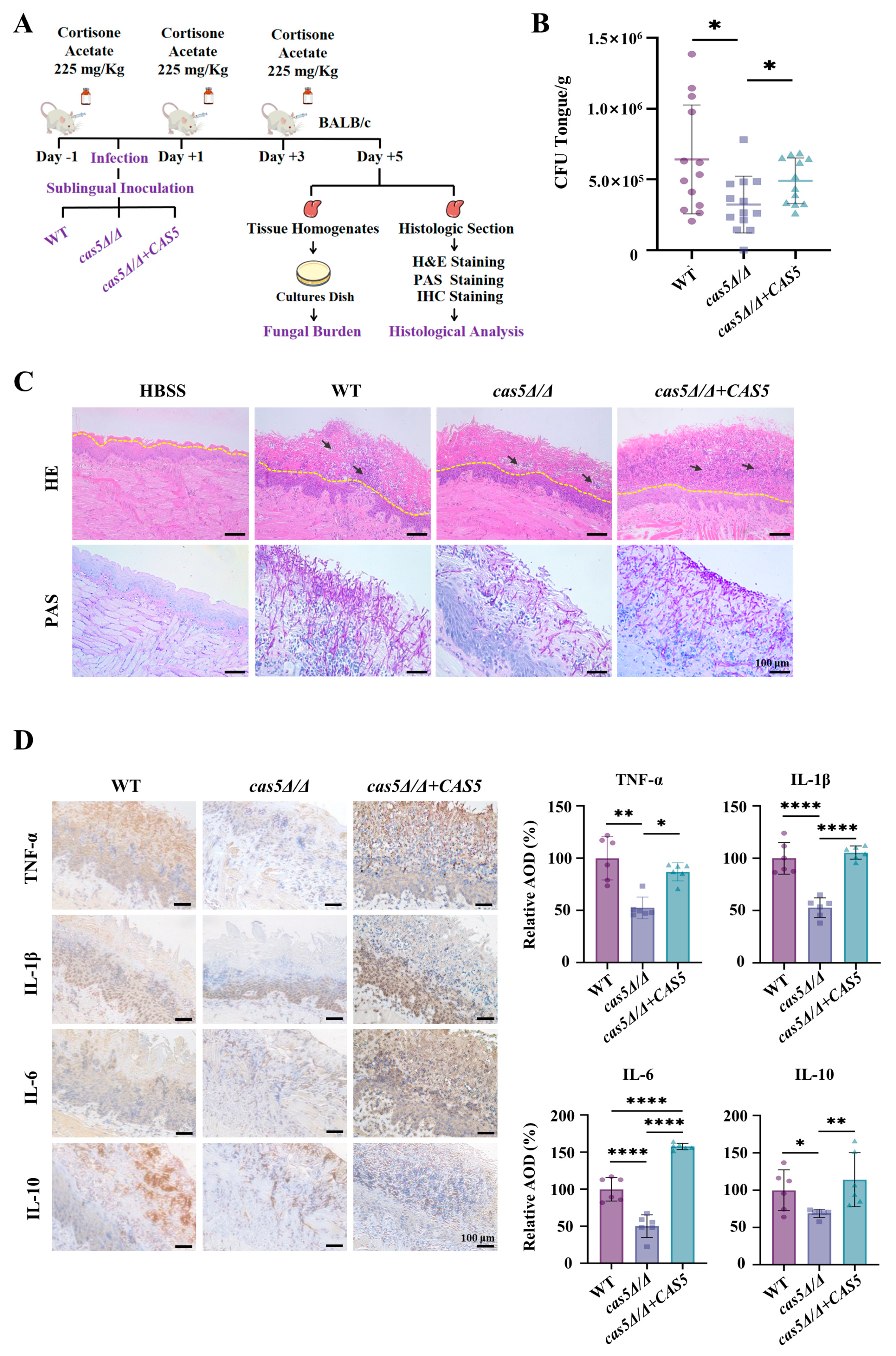
3.5. Cas5 Is Required for C. albicans Pathogenicity in Mouse Model of OPC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takagi, J.; Aoki, K.; Turner, B.S.; Lamont, S.; Lehoux, S.; Kavanaugh, N.; Gulati, M.; Valle Arevalo, A.; Lawrence, T.J.; Kim, C.Y.; et al. Mucin O-glycans are natural inhibitors of Candida albicans pathogenicity. Nat. Chem. Biol. 2022, 18, 762–773. [Google Scholar] [CrossRef]
- Lopes, J.P.; Lionakis, M.S. Pathogenesis and virulence of Candida albicans. Virulence 2022, 13, 89–121. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liu, J.; Chen, Y.; Yan, L.; Wu, J. Neutrophil Extracellular Traps in Candida albicans Infection. Front. Immunol. 2022, 13, 913028. [Google Scholar] [CrossRef] [PubMed]
- Kaur, J.; Nobile, C.J. Antifungal drug-resistance mechanisms in Candida biofilms. Curr. Opin. Microbiol. 2023, 71, 102237. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Huang, Y.; Tu, J.; Yang, W.; Liu, N.; Wang, W.; Sheng, C. Discovery of BRD4-HDAC Dual Inhibitors with Improved Fungal Selectivity and Potent Synergistic Antifungal Activity against Fluconazole-Resistant Candida albicans. J. Med. Chem. 2023, 66, 5950–5964. [Google Scholar] [CrossRef]
- Lv, Q.; Yan, L.; Wang, J.; Feng, J.; Gao, L.; Qiu, L.; Chao, W.; Qin, Y.L.; Jiang, Y. Combined Transcriptome and Metabolome Analysis Reveals That the Potent Antifungal Pyrylium Salt Inhibits Mitochondrial Complex I in Candida albicans. Microbiol. Spectr. 2023, 11, e0320922. [Google Scholar] [CrossRef]
- d’Enfert, C.; Kaune, A.K.; Alaban, L.R.; Chakraborty, S.; Cole, N.; Delavy, M.; Kosmala, D.; Marsaux, B.; Fróis-Martins, R.; Morelli, M.; et al. The impact of the Fungus-Host-Microbiota interplay upon Candida albicans infections: Current knowledge and new perspectives. FEMS Microbiol. Rev. 2021, 45, fuaa060. [Google Scholar] [CrossRef]
- Arita, G.S.; Faria, D.R.; Capoci, I.R.G.; Kioshima, E.S.; Bonfim-Mendonça, P.S.; Svidzinski, T.I.E. Cell wall associated proteins involved in filamentation with impact on the virulence of Candida albicans. Microbiol. Res. 2022, 258, 126996. [Google Scholar] [CrossRef]
- Chang, C.K.; Yang, M.C.; Chen, H.F.; Liao, Y.L.; Lan, C.Y. The Role of Sfp1 in Candida albicans Cell Wall Maintenance. J. Fungi 2022, 8, 1196. [Google Scholar] [CrossRef]
- Pérez-García, L.A.; Csonka, K.; Flores-Carreón, A.; Estrada-Mata, E.; Mellado-Mojica, E.; Németh, T.; López-Ramírez, L.A.; Toth, R.; López, M.G.; Vizler, C.; et al. Role of Protein Glycosylation in Candida parapsilosis Cell Wall Integrity and Host Interaction. Front. Microbiol. 2016, 7, 306. [Google Scholar] [CrossRef]
- Wang, W.H.; Lai, T.X.; Wu, Y.C.; Chen, Z.T.; Tseng, K.Y.; Lan, C.Y. Associations of Rap1 with Cell Wall Integrity, Biofilm Formation, and Virulence in Candida albicans. Microbiol. Spectr. 2022, 10, e0328522. [Google Scholar] [CrossRef] [PubMed]
- Hasim, S.; Allison, D.P.; Retterer, S.T.; Hopke, A.; Wheeler, R.T.; Doktycz, M.J.; Reynolds, T.B. β-(1,3)-Glucan Unmasking in Some Candida albicans Mutants Correlates with Increases in Cell Wall Surface Roughness and Decreases in Cell Wall Elasticity. Infect. Immun. 2017, 85, e00601-16. [Google Scholar] [CrossRef] [PubMed]
- Gantner, B.N.; Simmons, R.M.; Underhill, D.M. Dectin-1 mediates macrophage recognition of Candida albicans yeast but not filaments. Embo J. 2005, 24, 1277–1286. [Google Scholar] [CrossRef]
- Bain, J.M.; Louw, J.; Lewis, L.E.; Okai, B.; Walls, C.A.; Ballou, E.R.; Walker, L.A.; Reid, D.; Munro, C.A.; Brown, A.J.; et al. Candida albicans hypha formation and mannan masking of β-glucan inhibit macrophage phagosome maturation. mBio 2014, 5, e01874. [Google Scholar] [CrossRef]
- Munro, C.A.; Bates, S.; Buurman, E.T.; Hughes, H.B.; Maccallum, D.M.; Bertram, G.; Atrih, A.; Ferguson, M.A.; Bain, J.M.; Brand, A.; et al. Mnt1p and Mnt2p of Candida albicans are partially redundant alpha-1,2-mannosyltransferases that participate in O-linked mannosylation and are required for adhesion and virulence. J. Biol. Chem. 2005, 280, 1051–1060. [Google Scholar] [CrossRef]
- Ibe, C.; Munro, C.A. Fungal Cell Wall Proteins and Signaling Pathways Form a Cytoprotective Network to Combat Stresses. J. Fungi 2021, 7, 739. [Google Scholar] [CrossRef] [PubMed]
- Yadav, B.; Bhatnagar, S.; Ahmad, M.F.; Jain, P.; Pratyusha, V.A.; Kumar, P.; Komath, S.S. First step of glycosylphosphatidylinositol (GPI) biosynthesis cross-talks with ergosterol biosynthesis and Ras signaling in Candida albicans. J. Biol. Chem. 2014, 289, 3365–3382. [Google Scholar] [CrossRef] [PubMed]
- Cabral, V.; Znaidi, S.; Walker, L.A.; Martin-Yken, H.; Dague, E.; Legrand, M.; Lee, K.; Chauvel, M.; Firon, A.; Rossignol, T.; et al. Targeted changes of the cell wall proteome influence Candida albicans ability to form single- and multi-strain biofilms. PLoS Pathog. 2014, 10, e1004542. [Google Scholar] [CrossRef]
- Khodavandi, A.; Harmal, N.S.; Alizadeh, F.; Scully, O.J.; Sidik, S.M.; Othman, F.; Sekawi, Z.; Ng, K.P.; Chong, P.P. Comparison between allicin and fluconazole in Candida albicans biofilm inhibition and in suppression of HWP1 gene expression. Phytomedicine 2011, 19, 56–63. [Google Scholar] [CrossRef]
- Nobile, C.J.; Nett, J.E.; Andes, D.R.; Mitchell, A.P. Function of Candida albicans adhesin Hwp1 in biofilm formation. Eukaryot. Cell 2006, 5, 1604–1610. [Google Scholar] [CrossRef]
- Phan, Q.T.; Myers, C.L.; Fu, Y.; Sheppard, D.C.; Yeaman, M.R.; Welch, W.H.; Ibrahim, A.S.; Edwards, J.E., Jr.; Filler, S.G. Als3 is a Candida albicans invasin that binds to cadherins and induces endocytosis by host cells. PLoS Biol. 2007, 5, e64. [Google Scholar] [CrossRef]
- Blankenship, J.R.; Fanning, S.; Hamaker, J.J.; Mitchell, A.P. An extensive circuitry for cell wall regulation in Candida albicans. PLoS Pathog. 2010, 6, e1000752. [Google Scholar] [CrossRef]
- Xie, J.L.; Qin, L.; Miao, Z.; Grys, B.T.; Diaz, J.C.; Ting, K.; Krieger, J.R.; Tong, J.; Tan, K.; Leach, M.D.; et al. The Candida albicans transcription factor Cas5 couples stress responses, drug resistance and cell cycle regulation. Nat. Commun. 2017, 8, 499. [Google Scholar] [CrossRef]
- Xiong, K.; Su, C.; Sun, Q.; Lu, Y. Efg1 and Cas5 Orchestrate Cell Wall Damage Response to Caspofungin in Candida albicans. Antimicrob. Agents Chemother. 2021, 65, e01584-20. [Google Scholar] [CrossRef] [PubMed]
- Bruno, V.M.; Kalachikov, S.; Subaran, R.; Nobile, C.J.; Kyratsous, C.; Mitchell, A.P. Control of the C. albicans cell wall damage response by transcriptional regulator Cas5. PLoS Pathog. 2006, 2, e21. [Google Scholar] [CrossRef]
- Pataki, E.; Sipiczki, M.; Miklos, I. Schizosaccharomyces pombe rsv1 Transcription Factor and its Putative Homologues Preserved their Functional Homology and are Evolutionarily Conserved. Curr. Microbiol. 2017, 74, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Pukkila-Worley, R.; Peleg, A.Y.; Tampakakis, E.; Mylonakis, E. Candida albicans hyphal formation and virulence assessed using a Caenorhabditis elegans infection model. Eukaryot. Cell 2009, 8, 1750–1758. [Google Scholar] [CrossRef]
- Chamilos, G.; Nobile, C.J.; Bruno, V.M.; Lewis, R.E.; Mitchell, A.P.; Kontoyiannis, D.P. Candida albicans Cas5, a regulator of cell wall integrity, is required for virulence in murine and toll mutant fly models. J. Infect. Dis. 2009, 200, 152–157. [Google Scholar] [CrossRef]
- Finkel, J.S.; Xu, W.; Huang, D.; Hill, E.M.; Desai, J.V.; Woolford, C.A.; Nett, J.E.; Taff, H.; Norice, C.T.; Andes, D.R.; et al. Portrait of Candida albicans adherence regulators. PLoS Pathog. 2012, 8, e1002525. [Google Scholar] [CrossRef]
- Vyas, V.K.; Barrasa, M.I.; Fink, G.R. A Candida albicans CRISPR system permits genetic engineering of essential genes and gene families. Sci. Adv. 2015, 1, e1500248. [Google Scholar] [CrossRef]
- Care, R.S.; Trevethick, J.; Binley, K.M.; Sudbery, P.E. The MET3 promoter: A new tool for Candida albicans molecular genetics. Mol. Microbiol. 1999, 34, 792–798. [Google Scholar] [CrossRef] [PubMed]
- Dorward, D.W. Ultrastructural Analysis of Bacteria–Host Cell Interactions. In Bacterial Pathogenesis: Methods and Protocols; DeLeo, F.R., Otto, M., Eds.; Humana Press: Totowa, NJ, USA, 2008; pp. 173–187. [Google Scholar] [CrossRef]
- Kippert, F.; Lloyd, D. The aniline blue fluorochrome specifically stains the septum of both live and fixed Schizosaccharomyces pombe cells. FEMS Microbiol. Lett. 1995, 132, 215–219. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, R.; Li, M.; Ou, W.; Zhang, X.; Yang, W.; Huang, X.; Chai, H.; Wang, Q. TMT proteomics analysis of cerebrospinal fluid from patients with cerebral venous sinus thrombosis. J. Proteom. 2023, 275, 104820. [Google Scholar] [CrossRef]
- Ciok-Pater, E.; Gospodarek, E.; Prazyńska, M. Hydrophobic properties of Candida sp. comparing the two methods. Med. Dosw. Mikrobiol. 2008, 60, 243–251. [Google Scholar] [PubMed]
- Olivier, F.A.B.; Hilsenstein, V.; Weerasinghe, H.; Weir, A.; Hughes, S.; Crawford, S.; Vince, J.E.; Hickey, M.J.; Traven, A. The escape of Candida albicans from macrophages is enabled by the fungal toxin candidalysin and two host cell death pathways. Cell Rep. 2022, 40, 111374. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Song, Y.; Zhao, J.; Xu, B. Antifungal efficiency and cytocompatibility of polymethyl methacrylate modified with zinc dimethacrylate. Front. Cell Infect. Microbiol. 2023, 13, 1138588. [Google Scholar] [CrossRef]
- Solis, N.V.; Filler, S.G. Mouse model of oropharyngeal candidiasis. Nat. Protoc. 2012, 7, 637–642. [Google Scholar] [CrossRef]
- Fanning, S.; Xu, W.; Beaurepaire, C.; Suhan, J.P.; Nantel, A.; Mitchell, A.P. Functional control of the Candida albicans cell wall by catalytic protein kinase A subunit Tpk1. Mol. Microbiol. 2012, 86, 284–302. [Google Scholar] [CrossRef]
- Kamai, Y.; Kubota, M.; Kamai, Y.; Hosokawa, T.; Fukuoka, T.; Filler, S.G. Contribution of Candida albicans ALS1 to the pathogenesis of experimental oropharyngeal candidiasis. Infect. Immun. 2002, 70, 5256–5258. [Google Scholar] [CrossRef]
- Nakagawa, Y. Catalase gene disruptant of the human pathogenic yeast Candida albicans is defective in hyphal growth, and a catalase-specific inhibitor can suppress hyphal growth of wild-type cells. Microbiol. Immunol. 2008, 52, 16–24. [Google Scholar] [CrossRef]
- Pitarch, A.; Sánchez, M.; Nombela, C.; Gil, C. Sequential fractionation and two-dimensional gel analysis unravels the complexity of the dimorphic fungus Candida albicans cell wall proteome. Mol. Cell Proteom. 2002, 1, 967–982. [Google Scholar] [CrossRef]
- Shapiro, R.S.; Uppuluri, P.; Zaas, A.K.; Collins, C.; Senn, H.; Perfect, J.R.; Heitman, J.; Cowen, L.E. Hsp90 orchestrates temperature-dependent Candida albicans morphogenesis via Ras1-PKA signaling. Curr. Biol. 2009, 19, 621–629. [Google Scholar] [CrossRef]
- Mayer, F.L.; Wilson, D.; Hube, B. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Negri, M.; Henriques, M.; Oliveira, R.; Williams, D.W.; Azeredo, J. Adherence and biofilm formation of non-Candida albicans Candida species. Trends Microbiol. 2011, 19, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Masuoka, J.; Hazen, K.C. Cell wall mannan and cell surface hydrophobicity in Candida albicans serotype A and B strains. Infect. Immun. 2004, 72, 6230–6236. [Google Scholar] [CrossRef]
- Yang, M.; Solis, N.V.; Marshall, M.; Garleb, R.; Zhou, T.; Wang, D.; Swidergall, M.; Pearlman, E.; Filler, S.G.; Liu, H. Control of β-glucan exposure by the endo-1,3-glucanase Eng1 in Candida albicans modulates virulence. PLoS Pathog. 2022, 18, e1010192. [Google Scholar] [CrossRef]
- Bojang, E.; Ghuman, H.; Kumwenda, P.; Hall, R.A. Immune Sensing of Candida albicans. J. Fungi 2021, 7, 119. [Google Scholar] [CrossRef]
- Kawakita, M.; Oyama, T.; Shirai, I.; Tanaka, S.; Akaki, K.; Abe, S.; Asahi, T.; Cui, G.; Itoh, F.; Sasaki, M.; et al. Cell wall N-glycan of Candida albicans ameliorates early hyper- and late hypo-immunoreactivity in sepsis. Commun. Biol. 2021, 4, 342. [Google Scholar] [CrossRef]
- Spellberg, B.; Edwards, J.E., Jr. Type 1/Type 2 immunity in infectious diseases. Clin. Infect. Dis. 2001, 32, 76–102. [Google Scholar] [CrossRef]
- Duvenage, L.; Walker, L.A.; Bojarczuk, A.; Johnston, S.A.; MacCallum, D.M.; Munro, C.A.; Gourlay, C.W. Inhibition of Classical and Alternative Modes of Respiration in Candida albicans Leads to Cell Wall Remodeling and Increased Macrophage Recognition. mBio 2019, 10, e02535-18. [Google Scholar] [CrossRef]
- Cottier, F.; Sherrington, S.; Cockerill, S.; Del Olmo Toledo, V.; Kissane, S.; Tournu, H.; Orsini, L.; Palmer, G.E.; Pérez, J.C.; Hall, R.A. Remasking of Candida albicans β-Glucan in Response to Environmental pH Is Regulated by Quorum Sensing. mBio 2019, 10, e02347-19. [Google Scholar] [CrossRef]
- Heredia, M.Y.; Gunasekaran, D.; Ikeh, M.A.C.; Nobile, C.J.; Rauceo, J.M. Transcriptional regulation of the caspofungin-induced cell wall damage response in Candida albicans. Curr. Genet. 2020, 66, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Moyes, D.L.; Richardson, J.P.; Naglik, J.R. Candida albicans-epithelial interactions and pathogenicity mechanisms: Scratching the surface. Virulence 2015, 6, 338–346. [Google Scholar] [CrossRef]
- Yoshijima, Y.; Murakami, K.; Kayama, S.; Liu, D.; Hirota, K.; Ichikawa, T.; Miyake, Y. Effect of substrate surface hydrophobicity on the adherence of yeast and hyphal Candida. Mycoses 2010, 53, 221–226. [Google Scholar] [CrossRef]
- Kim, J.M.; Moon, H.Y.; Lee, D.W.; Kang, H.A.; Kim, J.Y. The transcription factor Cas5 suppresses hyphal morphogenesis during yeast-form growth in Candida albicans. J. Microbiol. 2021, 59, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Bockmühl, D.P.; Ernst, J.F. A potential phosphorylation site for an A-type kinase in the Efg1 regulator protein contributes to hyphal morphogenesis of Candida albicans. Genetics 2001, 157, 1523–1530. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.; Johnston, D.; Phan, Q.T.; Edwards, J.E., Jr.; French, S.W.; Ibrahim, A.S.; Filler, S.G. Parenchymal organ, and not splenic, immunity correlates with host survival during disseminated candidiasis. Infect. Immun. 2003, 71, 5756–5764. [Google Scholar] [CrossRef]
- Naglik, J.R.; König, A.; Hube, B.; Gaffen, S.L. Candida albicans-epithelial interactions and induction of mucosal innate immunity. Curr. Opin. Microbiol. 2017, 40, 104–112. [Google Scholar] [CrossRef]
- Nikou, S.A.; Zhou, C.; Griffiths, J.S.; Kotowicz, N.K.; Coleman, B.M.; Green, M.J.; Moyes, D.L.; Gaffen, S.L.; Naglik, J.R.; Parker, P.J. The Candida albicans toxin candidalysin mediates distinct epithelial inflammatory responses through p38 and EGFR-ERK pathways. Sci. Signal 2022, 15, eabj6915. [Google Scholar] [CrossRef]
- Arocho, A.; Chen, B.Y.; Ladanyi, M.; Pan, Q.L. Validation of the 2(-Delta Delta Ct) calculation as an alternate method of data analysis for quantitative PCR of BCR-ABL P210 transcripts. Diagn. Mol. Pathol. 2006, 15, 56–61. [Google Scholar] [CrossRef]
- Ma, J.; Chen, T.; Wu, S.; Yang, C.; Bai, M.; Shu, K.; Li, K.; Zhang, G.; Jin, Z.; He, F.; et al. iProX: An integrated proteome resource. Nucleic Acids Res. 2019, 47, D1211–D1217. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Ma, J.; Liu, Y.; Chen, Z.; Xiao, N.; Lu, Y.; Fu, Y.; Yang, C.; Li, M.; Wu, S.; et al. iProX in 2021: Connecting proteomics data sharing with big data. Nucleic Acids Res. 2022, 50, D1522–D1527. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P. The Effects of Cas5 on the Protein Biosynthesis of Candida albicans [Database]. ProteomeXchange. Available online: https://proteomecentral.proteomexchange.org/cgi/GetDataset?ID=PXD052043 (accessed on 6 May 2024).
- Zhang, P. Cas5 promotes Candida albicans virulence via regulating cell wall remodeling [database]. Figshare 2024. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Li, G.; Wang, Y.; Yang, C.; Bai, W.; Li, Q.; Zhang, J.; Zhang, P. Cas5 Regulates the Exposure of β-Glucan, the Cell Surface Hydrophobicity, and the Expression of Cell Wall Proteins to Remodel the Candida albicans Cell Wall and Participates in the Recruitment of Neutrophils. Microorganisms 2025, 13, 683. https://doi.org/10.3390/microorganisms13030683
Zhang Q, Li G, Wang Y, Yang C, Bai W, Li Q, Zhang J, Zhang P. Cas5 Regulates the Exposure of β-Glucan, the Cell Surface Hydrophobicity, and the Expression of Cell Wall Proteins to Remodel the Candida albicans Cell Wall and Participates in the Recruitment of Neutrophils. Microorganisms. 2025; 13(3):683. https://doi.org/10.3390/microorganisms13030683
Chicago/Turabian StyleZhang, Qiyue, Guanglin Li, Yanmei Wang, Chen Yang, Wenhui Bai, Qingqing Li, Jiye Zhang, and Peipei Zhang. 2025. "Cas5 Regulates the Exposure of β-Glucan, the Cell Surface Hydrophobicity, and the Expression of Cell Wall Proteins to Remodel the Candida albicans Cell Wall and Participates in the Recruitment of Neutrophils" Microorganisms 13, no. 3: 683. https://doi.org/10.3390/microorganisms13030683
APA StyleZhang, Q., Li, G., Wang, Y., Yang, C., Bai, W., Li, Q., Zhang, J., & Zhang, P. (2025). Cas5 Regulates the Exposure of β-Glucan, the Cell Surface Hydrophobicity, and the Expression of Cell Wall Proteins to Remodel the Candida albicans Cell Wall and Participates in the Recruitment of Neutrophils. Microorganisms, 13(3), 683. https://doi.org/10.3390/microorganisms13030683