

Responses of Soil Microbial Communities Associated with Phosphorus Transformation to Land-Use Alternations in a Meadow Grassland, Northeast China


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Location and Site Description
2.2. Experimental Design and Soil Sampling
2.3. Soil Property Determinations
2.4. Amplification, Quantification, and Sequencing of phoD and gcd Genes
2.5. Data Analysis
3. Results
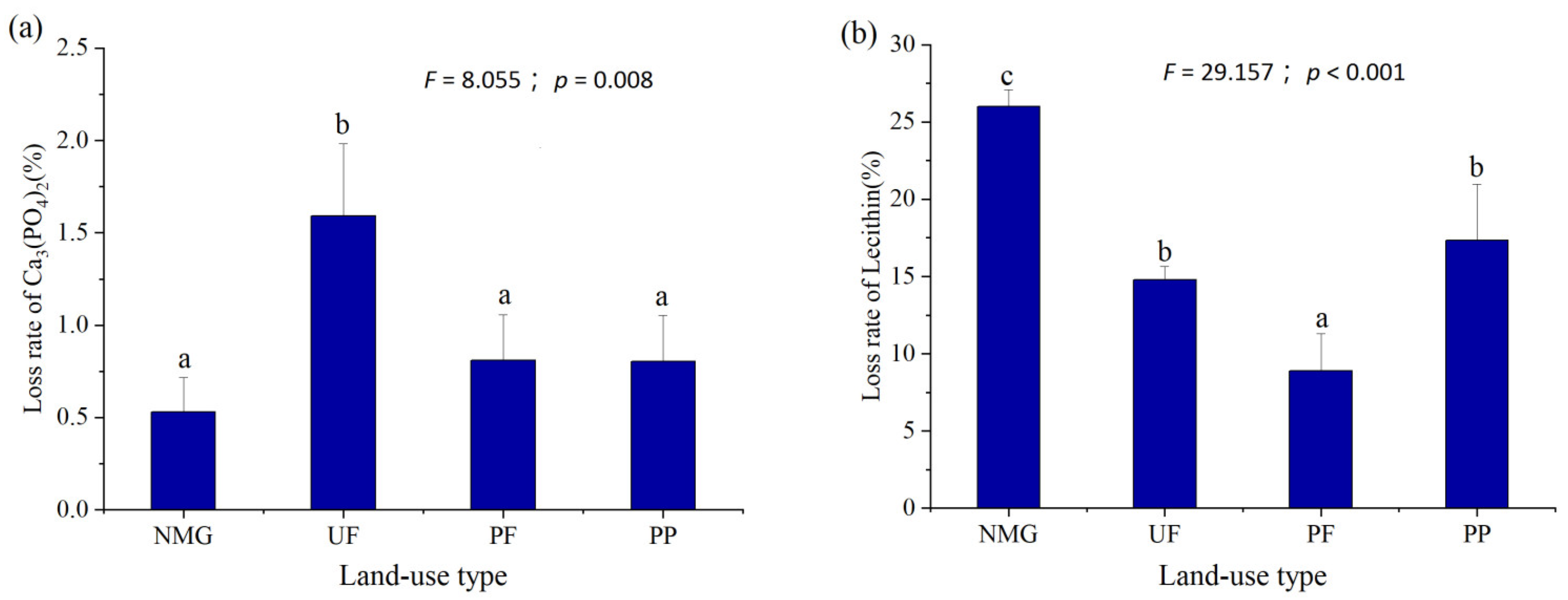
3.1. Soil Properties, P Fractions, and Potential of P Transformation
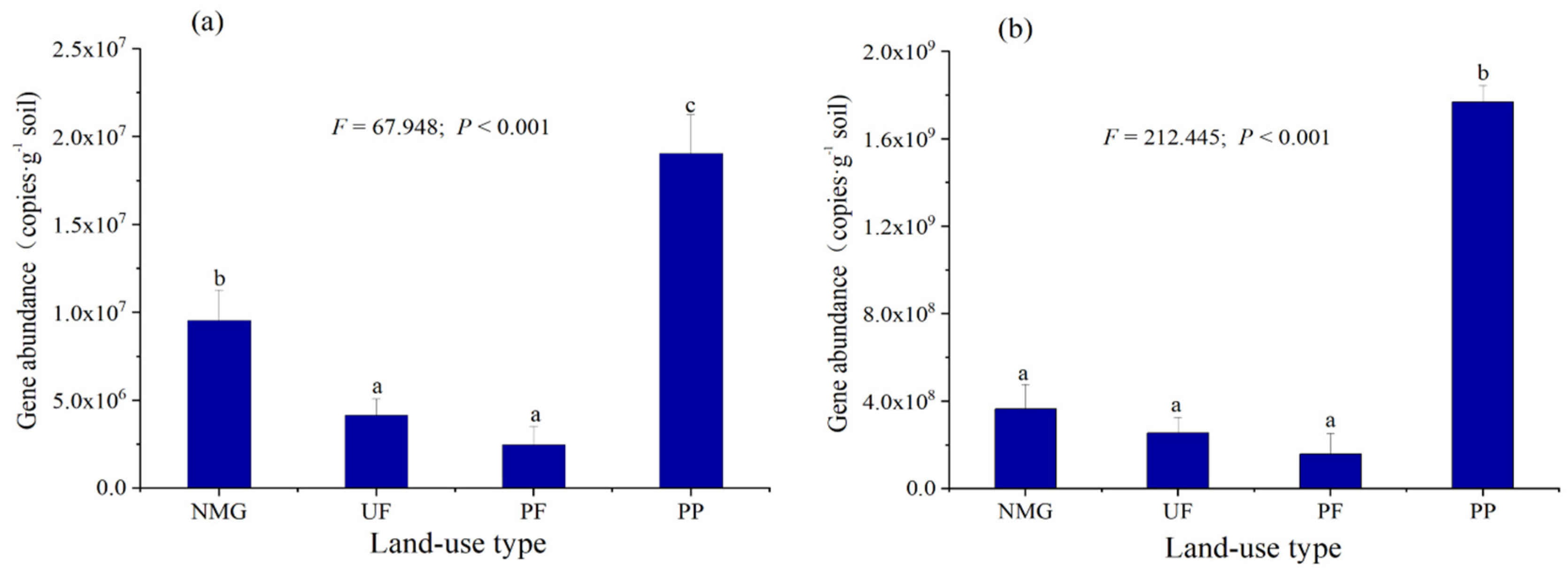
3.2. Abundances of gcd and phoD Genes
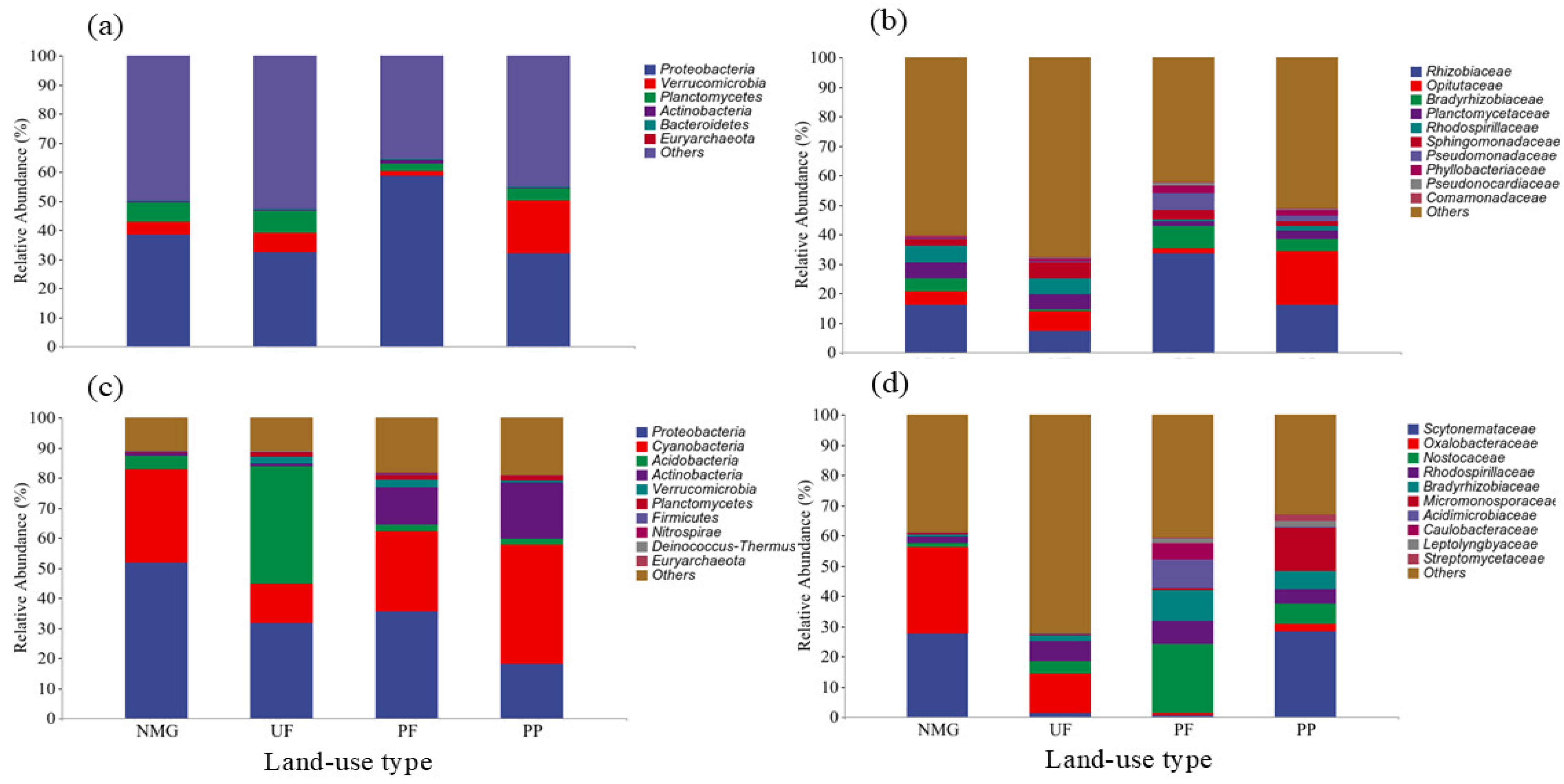
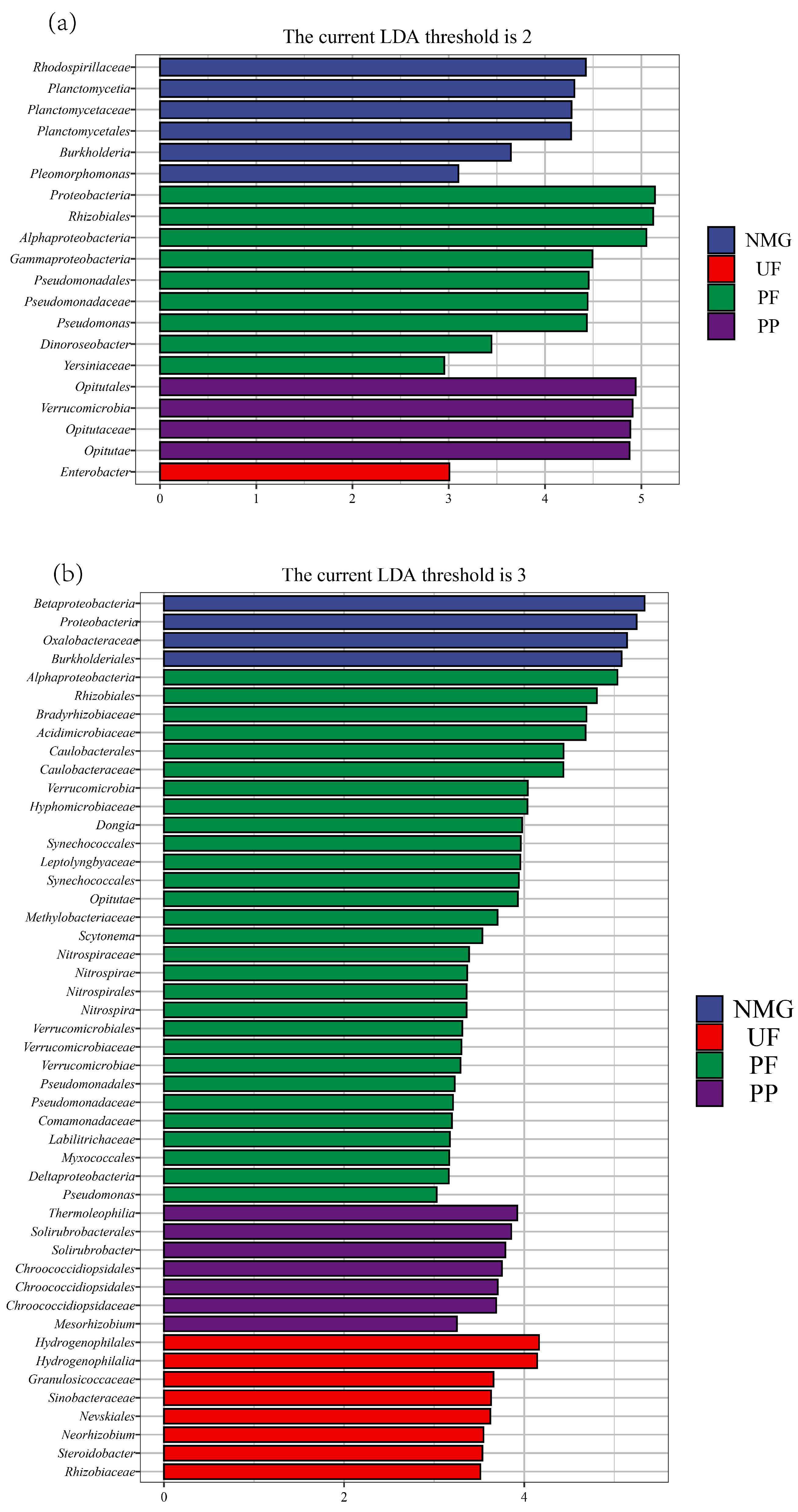
3.3. Diversities and Compositions of gcd- and phoD-Harboring Microbial Communities
3.4. Influence of Soil Properties on phoD and gcd Communities
4. Discussion
4.1. The Influence of Land-Use Type on Soil Properties
4.2. The Responses of Soil gcd and phoD Microbial Communities to Land-Use Type
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cui, F.Q.; Wang, B.J.; Zhang, Q.; Tang, H.P.; De Maeyer, P.; Hamdi, R.; Dai, L.W. Climate change versus land-use change—What affects the ecosystem services more in the forest-steppe ecotone? Sci. Total Environ. 2021, 759, 143525. [Google Scholar] [CrossRef] [PubMed]
- Li, F.R.; Liu, J.L.; Ren, W.; Liu, L.L. Land-use change alters patterns of soil biodiversity in arid lands of northwestern China. Plant Soil 2018, 428, 371–388. [Google Scholar] [CrossRef]
- Paltineanu, C.; Dumitru, S.; Vizitiu, O.; Mocanu, V.; Lăcătusu, A.R.; Ion, S.; Domnariu, H. Soil organic carbon and total nitrogen stocks related to land use and basic environmental properties−assessment of soil carbon sequestration potential in different ecosystems. Catena 2024, 246, 108435. [Google Scholar] [CrossRef]
- Zhu, R.H.; Azene, B.; Gruba, P.; Pan, K.; Nigussie, Y.; Guadie, A.; Sun, X.M.; Wu, X.G.; Zhang, L. Response of carbohydrate-degrading enzymes and microorganisms to land use change in the southeastern Qinghai-Tibetan Plateau, China. Appl. Soil Ecol. 2024, 200, 105442. [Google Scholar] [CrossRef]
- Liu, S.L.; Sun, Y.X.; Shi, F.N.; Liu, Y.X.; Wang, F.F.; Dong, S.K.; Li, M.Q. Composition and diversity of soil microbial community associated with land use types in the agro-pastoral area in the upper Yellow River Basin. Front. Plant Sci. 2022, 13, 819661. [Google Scholar] [CrossRef]
- Wang, J.C.; Zou, Y.K.; Di, G.D.; Singh, B.K.; Li, Q.F. Conversion to agroforestry and monoculture plantation is detrimental to the soil carbon and nitrogen cycles and microbial communities of a rainforest. Soil Biol. Biochem. 2020, 147, 107849. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef]
- Condron, L.M.; Turner, B.L.; Cademenun, B.J.; Sims, J.T.; Sharpley, A.N. Chemistry and dynamics of soil organic phosphorus. Phosphorus Agric. Environ. 2005, 46, 87–121. [Google Scholar]
- Mahdi, I.; Fahsi, N.; Hafidi, M.; Allaoui, A.; Biskri, L. Plant growth enhancement using rhizospheric halotolerant phosphate solubilizing bacterium Bacillus licheniformis QA1 and Enterobacter asburiae QF11 isolated from Chenopodium quinoa willd. Microorganisms 2020, 8, 948. [Google Scholar] [CrossRef]
- Mendoza-Arroyo, G.E.; Chan-Bacab, M.J.; Aguila-Ramírez, R.N.; Ortega-Morales, B.O.; Canché Solís, R.E.; Chab-Ruiz, A.O.; Cob-Rivera, K.I.; Dzib-Castillo, B.; Tun-Che, R.E.; Camacho-Chab, J.C. Inorganic phosphate solubilization by a novel isolated bacterial strain Enterobacter sp. ITCB-09 and its application potential as biofertilizer. Agriculture 2020, 10, 383. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Landi, L.; Renella, G. Role of Phosphatase Enzymes in Soil. In Phosphorus in Action; Else, B., Astrid, O., Emmanuel, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 26, pp. 215–243. [Google Scholar]
- Li, J.B.; Xie, T.; Zhu, H.; Zhou, J.; Li, C.N.; Xiong, W.J.; Xu, L.; Wu, Y.H.; He, Z.L.; Li, X.Z. Alkaline phosphatase activity mediates soil organic phosphorus mineralization in a subalpine forest ecosystem. Geoderma 2021, 404, 115376. [Google Scholar] [CrossRef]
- Fraser, T.; Lynch, D.H.; Entz, M.H.; Dunfield, K.E. Linking alkaline phosphatase activity with bacterial phoD gene abundance in soil from a long-term management trial. Geoderma 2015, 257, 115–122. [Google Scholar] [CrossRef]
- Kageyama, H.; Tripathi, K.; Rai, A.K.; Cha-Um, S.; Waditee-Sirisattha, R.; Takabe, T. An alkaline phosphatase/phosphodiesterase, phoD, induced by salt stress and secreted out of the cells of Aphanothece halophytica, a halotolerant cyanobacterium. Appl. Environ. Microbiol. 2011, 77, 5178–5183. [Google Scholar] [CrossRef]
- Wu, J.R.; Shien, J.H.; Shieh, H.K.; Hu, C.C.; Gong, S.R.; Chen, L.Y.; Chang, P.C. Cloning of the gene and characterization of the enzymatic properties of the monomeric alkaline phosphatase (PhoX) from Pasteurella multocida strain X-73. FEMS Microbiol. Lett. 2007, 267, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Barret, M.; Mooij, M.J.; Rice, O.; Morrissey, J.P.; Dobson, A.; Griffiths, B.; O’Gara, F. Long-term phosphorus fertilization increased the diversity of the total bacterial community and the phoD phosphorus mineralizer group in pasture soils. Biol. Fertil. Soils 2013, 49, 661–672. [Google Scholar] [CrossRef]
- Zeng, Q.W.; Wu, X.Q.; Wen, X.Y. Effects of soluble phosphate on phosphate-solubilizing characteristics and expression of gcd gene in Pseudomonas frederiksbergensis JW-SD2. Curr. Microbiol. 2016, 72, 198–206. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, J.Q.; Gong, Z.L.; Xu, W.L.; Mou, Z.S. Gcd gene diversity of quinoprotein glucose dehydrogenase in the sediment of Sancha Lake and its response to the environment. Int. J. Environ. Res. Public Health 2019, 16, 1. [Google Scholar] [CrossRef]
- Cao, C.Y.; Jiang, D.M.; Teng, X.H.; Jiang, Y.; Liang, W.J.; Cui, Z.B. Soil chemical and microbiological properties along a chronosequence of Caragana microphylla Lam. plantations in the Horqin Sandy Land of Northeast China. Appl. Soil Ecol. 2008, 40, 78–85. [Google Scholar] [CrossRef]
- ISSCAS (Institute of Soil Science, Chinese Academy of Sciences). Physical and Chemical Analysis Methods of Soils; Shanghai Science Technology Press: Shanghai, China, 1978; pp. 7–59. [Google Scholar]
- Ladd, J.N.; Butler, J.H.A. Short-term assays of soil proteolytic enzyme activities using proteins and dipeptide derivatives as substrates. Soil Biol. Biochem. 1972, 4, 19–30. [Google Scholar] [CrossRef]
- ISSCAS (Institute of Soil Science, Chinese Academy of Sciences). Methods on Soil Microorganisms Study; Science Press: Beijing, China, 1985; pp. 260–275. [Google Scholar]
- Perucci, P.; Casucci, C.; Dumontet, S. An improved method to evaluate the odiphenol oxidase activity of soil. Soil Biol. Biochem. 2000, 32, 1927–1933. [Google Scholar] [CrossRef]
- Schinner, F.; Öhlinger, R.; Kandeler, E.; Margesin, R. Methods in soil biology. J. Ecol. 1997, 85, 404–405. [Google Scholar]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Hedley, M.J.; Stewart, J.W.B.; Chauhan, B.S. Changes in inorganic and organic soil phosphorus fractions induced by cultivation practices and by laboratory incubations. Soil Sci. Soc. Am. J. 1982, 46, 970–976. [Google Scholar] [CrossRef]
- Da Silva, V.M.; Antoniolli, Z.I.; Jacques, R.J.S.; Ott, R.; Rodrigues, P.E.D.; Andrade, F.V.; Passos, R.R.; Mendonca, E.D. Influence of the tropical millipede, Glyphiulus granulatus (Gervais, 1847), on aggregation, enzymatic activity, and phosphorus fractions in the soil. Geoderma 2017, 289, 135–141. [Google Scholar] [CrossRef]
- Zheng, H.Y.; Zhang, D.S. Methods on Dynamics of Soil Biochemistry Study; Science Press: Beijing, China, 1982; pp. 167–169. [Google Scholar]
- Chen, X.D.; Jiang, N.; Condron, L.M.; Dunfield, K.E.; Chen, Z.H.; Wang, J.K.; Chen, L.J. Impact of long-term phosphorus fertilizer inputs on bacterial phoD gene community in a maize field, Northeast China. Sci. Total Environ. 2019, 669, 1011–1018. [Google Scholar] [CrossRef]
- Bergkemper, F.; Kublik, S.; Lang, F.; Kruger, J.; Vestergaard, G.; Schloter, M.; Schulz, S. Novel oligonucleotide primers reveal a high diversity of microbes which drive phosphorous turnover in soil. J. Microbiol. Methods 2016, 125, 91–97. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Opensource, platform independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J. Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef]
- Xie, Y.Y.; Wang, F.H.; Wang, K.; Yue, H.Z.; Lan, X.F. Responses of bacterial phoD gene abundance and diversity to crop rotation and feedbacks to phosphorus uptake in wheat. App. Soil Ecol. 2020, 154, 103604. [Google Scholar] [CrossRef]
- Wang, M.; Wu, Y.; Zhao, J.; Liu, Y.; Chen, Z.; Tang, Z. Long-term fertilization lowers the alkaline phosphatase activity by impacting the phoD-harboring bacterial community in rice-winter wheat rotation system. Sci. Total Environ. 2022, 821, 153406. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, H.; Xu, C.; Yuan, J.; Xu, X.J.; Wang, J.D.; Zhang, Y.C. Long-term nitrogen fertilization and sweet potato cultivation in the wheat-sweet potato rotation system decrease alkaline phosphomonoesterase activity by regulating soil phoD-harboring bacteria communities. Sci. Total Environ. 2023, 900, 165916. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Zou, J.L.; Meng, D.L.; Dang, S.N.; Zhou, J.H.; Osborne, B.; Ren, Y.Y.; Liang, T.; Yu, K.K. Effect of soil microorganisms and labile C availability on soil respiration in response to litter inputs in forest ecosystems: A meta-analysis. Ecol. Evol. 2020, 10, 13602–13612. [Google Scholar] [CrossRef] [PubMed]
- Clemmensen, K.E.; Bahr, A.; Ovaskainen, O.; Dahlberg, A.; Ekblad, A.; Wallander, H.; Stenlid, J.; Finlay, R.D.; Wardle, D.A.; Lindahl, B.D. Roots and associated fungi drive long-term carbon sequestration in boreal forest. Science 2023, 339, 1615–1618. [Google Scholar] [CrossRef]
- Lauber, C.L.; Ramirez, K.S.; Aanderud, Z.; Lennon, J.; Fierer, N. Temporal variability in soil microbial communities across land-use types. ISME J. 2013, 7, 1641–1650. [Google Scholar] [CrossRef]
- Creamer, R.E.; Hannula, S.E.; Van Leeuwen, J.P.; Stone, D.; Rutgers, M. Ecological network analysis reveals the inter-connection between soil biodiversity and ecosystem function as affected by land use across Europe. Appl. Soil Ecol. 2016, 97, 112–124. [Google Scholar] [CrossRef]
- Wang, Z.R.; Li, F.R.; Liu, L.L.; Yang, K. Changes in soil phosphorus fractions under different land uses in desert grasslands in northwestern China. Eur. J. Soil Sci. 2022, 73, e13255. [Google Scholar] [CrossRef]
- García-Velázquez, L.; Rodríguez, A.; Gallardo, A.; Maestre, F.; Santos, E.; Lafuente, A.; Fernández, M.; Singh, B.; Wang, J.; Durán, J. Climate and soil micro-organisms drive soil phosphorus fractions in coastal dune systems. Funct. Ecol. 2020, 34, 1690–1701. [Google Scholar] [CrossRef]
- Chen, L.F.; He, Z.B.; Zhao, W.Z.; Liu, J.L.; Zhou, H.; Li, J.; Meng, Y.Y.; Wang, L.S. Soil structure and nutrient supply drive changes in soil microbial communities during conversion of virgin desert soil to irrigated cropland. Eur. J. Soil Sci. 2019, 71, 768–781. [Google Scholar] [CrossRef]
- Feng, J.; He, K.; Zhang, Q.; Han, M.; Zhu, B. Changes in plant inputs alter soil carbon and microbial communities in forest ecosystems. Glob. Change Biol. 2022, 28, 3426–3440. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Bhattacharyya, R.; Dalal, R.C.; Wang, P.; Menzies, N.W.; Kopittke, P.M. Impact of land use change and soil type on total phosphorus and its fractions in soil aggregates. Land Degrad. Dev. 2020, 31, 828–841. [Google Scholar] [CrossRef]
- Wang, R.Z.; Liu, H.Y.; Sardans, J.; Feng, X.; Xu, Z.W.; Peñuelas, J. Interacting effects of urea and water addition on soil mineral bound phosphorus dynamics in semi-arid grasslands with different land-use history. Eur. J. Soil Sci. 2021, 72, 946–962. [Google Scholar] [CrossRef]
- Hu, M.J.; Penuelas, J.; Sardans, J.; Tong, C.; Chang, C.T.; Cao, W.Z. Dynamics of phosphorus speciation and the phoD phosphatase gene community in the rhizosphere and bulk soil along an estuarine freshwater-oligohaline gradient. Geoderma 2020, 365, 114236. [Google Scholar] [CrossRef]
- Turner, B.L.; Haygarth, P.M. Phosphatase activity in temperate pasture soils: Potential regulation of labile organic phosphorus turnover by phosphodiesterase activity: Linking landscape sources of phosphorus and sediment to ecological impacts in surface waters. Sci. Total Environ. 2005, 344, 27–36. [Google Scholar] [CrossRef]
- Ragot, S.A.; Kertesz, M.A.; Bünemann, E.K. phoD alkaline phosphatase gene diversity in soil. Appl. Environ. Microbiol. 2015, 81, 7281–7289. [Google Scholar] [CrossRef]
- Luo, G.W.; Ling, N.; Nannipieri, P.; Chen, H.; Raza, W.; Wang, M.; Guo, S.W.; Shen, Q.R. Long-term fertilisation regimes affect the composition of the alkaline phosphomonoesterase encoding microbial community of a vertisol and its derivative soil fractions. Biol. Fertil. Soils 2017, 53, 375–388. [Google Scholar] [CrossRef]
- Cheng, J.; Zhang, Y.; Wang, H.N.; Cui, Z.B.; Cao, C.Y. Sand-fixation plantation type affects soil phosphorus transformation microbial community in a revegetation area of Horqin Sandy Land, Northeast China. Ecol. Eng. 2022, 180, 106644. [Google Scholar] [CrossRef]
- Suleiman, A.K.A.; Manoeli, L.; Boldo, J.T. Shifts in soil bacterial community after eight years of land-use change. Syst. Appl. Microbiol. 2013, 36, 137–144. [Google Scholar] [CrossRef]
- Turley, N.E.; Bell-Dereske, L.; Evans, S.E.; Brudvig, L.A.; Yang, G.W. Agricultural land-use history and restoration impact soil microbial biodiversity. J. Appl. Ecol. 2020, 57, 852–863. [Google Scholar] [CrossRef]
- Kalayu, G. Phosphate solubilizing microorganisms: Promising approach as biofertilizers. Int. J. Agron. 2019, 7, 4917256. [Google Scholar] [CrossRef]
- Zhang, L.C.; Ren, G.; Chu, G.X. Land reclamation increased labile and moderately labile P fractions and strengthened co-occurrence network of gcd community in calcareous soils. Land Degrad. Dev. 2023, 34, 5542–5555. [Google Scholar] [CrossRef]
- Du, T.Y.; Hu, Q.F.; Mao, W.J.; Yang, Z.; Chen, H.; Sun, L.N.; Zhai, M.Z. Metagenomics insights into the functional profiles of soil carbon, nitrogen, and phosphorus cycles in a walnut orchard under various regimes of long-term fertilisation. Eur. J. Agron. 2023, 148, 126887. [Google Scholar] [CrossRef]
- Ragot, S.A.; Huguenin-Elie, O.; Kertesz, M.A.; Frossard, E.; Bunemann, E.K. Total and active microbial communities and phoD as affected by phosphate depletion and pH in soil. Plant Soil 2016, 408, 15–30. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | NMG | UF | PF | PP | F | p |
|---|---|---|---|---|---|---|
| SM | 2.89 ± 1.12 a | 2.01 ± 1.47 a | 2.82 ± 0.54 a | 4.72 ± 0.39 a | 4.048 | 0.051 |
| pH | 8.49 ± 0.28 b | 8.35 ± 0.10 b | 7.16 ± 0.70 a | 7.85 ± 0.05 ab | 7.52 | 0.01 |
| EC | 227.7 ± 18.2 b | 218.8 ± 69.4 b | 136.4 ± 8.4 a | 334.3 ± 17.5 c | 14.33 | 0.001 |
| AK | 86.1 ± 3.8 ab | 106.0 ± 24.1 b | 61.0 ± 10.1 a | 628.5 ± 4.6 c | 1239.8 | <0.001 |
| AP | 2.91 ± 0.51 a | 5.28 ± 1.48 a | 13.11 ± 1.21 c | 9.38 ± 1.28 b | 29.056 | 0.004 |
| NH4-N | 2.27 ± 0.35 a | 1.32 ± 0.03 a | 7.01 ± 3.02 b | 2.65 ± 0.45 a | 8.089 | 0.008 |
| TP | 0.38 ± 0.006 b | 0.19 ± 0.001 a | 0.19 ± 0.005 a | 0.46 ± 0.008 c | 2072.1 | <0.001 |
| TK | 1.82 ± 0.02 b | 1.73 ± 0.02 a | 1.72 ± 0.01 a | 1.75 ± 0.03 a | 8.562 | 0.007 |
| TN | 0.036 ± 0.004 a | 0.048 ± 0.008 a | 0.030 ± 0.009 a | 0.190 ± 0.034 b | 53.24 | <0.001 |
| SOM | 1.05 ± 0.07 a | 1.32 ± 0.19 a | 1.20 ± 0.19 a | 2.51 ± 0.31 b | 94.62 | <0.001 |
| PRA | 152.2 ± 48.0 a | 51.9 ± 6.82 a | 119.1 ± 73.6 a | 461.7 ± 112.2 b | 19.48 | <0.001 |
| DEA | 629.4 ± 29.0 c | 211.0 ± 43.5 ab | 77.9 ± 17.8 a | 351.7 ± 144.1 b | 28.08 | <0.001 |
| POA | 2.44 ± 0.06 b | 0.23 ± 0.142 a | 0.38 ± 0.140 a | 2.72 ± 0.52 b | 66.77 | <0.001 |
| ALP | 72.6 ± 3.98 b | 25.8 ± 7.39 a | 14.1 ± 4.92 a | 91.4 ± 7.80 c | 105.5 | <0.001 |
| URA | 133.0 ± 71.3 b | 374.9 ± 13.4 c | 37.5 ± 4.6 a | 569.3 ± 26.8 d | 115.5 | <0.001 |
| NRA | 0.110 ± 0.023 a | 0.224 ± 0.087 a | 0.023 ± 0.017 a | 0.735 ± 0.511 b | 4.693 | 0.036 |
| Fraction | NMG | UF | PF | PP | F | p |
|---|---|---|---|---|---|---|
| H2O-Pi | 2.00 ± 0.27 a | 9.17 ± 9.34 a | 5.09 ± 0.84 a | 7.44 ± 3.17 a | 1.18 | 0.377 |
| NaHCO3-Pi (0.5 mol·L−1) | 6.02 ± 0.08 a | 17.47 ± 9.58 b | 35.53 ± 4.34 c | 20.61 ± 5.46 b | 12.63 | 0.002 |
| NaHCO3-Po (0.5 mol·L−1) | 4.55 ± 0.57 a | 5.63 ± 1.53 a | 9.87 ± 3.97 a | 7.78 ± 0.45 a | 3.595 | 0.066 |
| NaOH-Pi (0.1 mol·L−1) | 2.70 ± 0.20 a | 5.06 ± 0.99 b | 26.22 ± 1.74 d | 19.25 ± 1.09 c | 293.3 | <0.001 |
| NaOH-Po (0.1 mol·L−1) | 20.8 ± 3.25 a | 19.2 ± 1.04 a | 24.7 ± 1.17 a | 79.1 ± 25.06 b | 15.56 | 0.001 |
| HCl-Pi (1.0 mol·L−1) | 20.0 ± 1.00 a | 64.3 ± 54.01 a | 28.3 ± 5.40 a | 20.5 ± 1.26 a | 1.802 | 0.225 |
| HCl-Po (1.0 mol·L−1) | 27.6 ± 7.32 a | 57.0 ± 36.05 a | 28.4 ± 11.69 a | 77.2 ± 9.57 a | 4.374 | 0.042 |
| NaOH-Pi (0.5 mol·L-1) | 3.02 ± 0.38 a | 7.71 ± 0.73 b | 8.22 ± 1.13 b | 12.29 ± 0.20 c | 86.96 | <0.001 |
| NaOH-Po (0.5 mol·L-1) | 27.9 ± 6.44 a | 37.9 ± 7.12 a | 22.0 ± 8.54 a | 70.0 ± 7.99 b | 24.03 | <0.001 |
| Residual-P | 484.3 ± 57.5 a | 1579.0 ± 111.0 c | 816.0 ± 20.9 b | 1523.7 ± 20.2 c | 210.9 | <0.001 |
| General-P | 596.9 ± 51.3 a | 1793.3 ± 129.1 c | 999.2 ± 30.1 b | 1830.4 ± 56.0 c | 190.1 | <0.001 |
| Index | Gene | NMG | UF | PF | PP | F | p |
|---|---|---|---|---|---|---|---|
| Chao1 | phoD | 1241.2 ± 100.9 a | 1421.9 ± 279.2 a | 1139.1 ± 191.0 a | 2425.1 ± 154.6 b | 28.183 | <0.001 |
| gcd | 2885.3 ± 244.3 c | 2235.6 ± 216.4 b | 1650.5 ± 114.6 a | 3541.6 ± 225.8 d | 46.901 | <0.001 | |
| Observed species | phoD | 652.2 ± 60.5 a | 828.0 ± 179.1 a | 815.3 ± 154.6 a | 1323.2 ± 94.9 b | 14.726 | 0.001 |
| gcd | 2029.1 ± 160.6 c | 1487.8 ± 213.6 b | 1099.1 ± 72.0 a | 2616.2 ± 21.0 d | 67.861 | <0.001 | |
| Pielou | phoD | 0.460 ± 0.036 a | 0.599 ± 0.040 b | 0.651 ± 0.045 b | 0.625 ± 0.006 b | 17.251 | 0.001 |
| gcd | 0.758 ± 0.014 b | 0.733 ± 0.011 b | 0.649 ± 0.044 a | 0.740 ± 0.054 b | 5.548 | 0.023 | |
| Shannon– Wiener | phoD | 4.30 ± 0.38 a | 5.81 ± 0.58 b | 6.29 ± 0.61 b | 6.48 ± 0.07 b | 13.602 | 0.002 |
| gcd | 8.32 ± 0.09 b | 7.72 ± 0.26 b | 6.56 ± 0.50 a | 8.40 ± 0.62 b | 12.356 | 0.002 | |
| Simpson | phoD | 0.845 ± 0.041 a | 0.935 ± 0.017 b | 0.948 ± 0.019 b | 0.923 ± 0.021 b | 9.443 | 0.005 |
| gcd | 0.986 ± 0.004 b | 0.981 ± 0.004 b | 0.924 ± 0.032 a | 0.977 ± 0.023 b | 6.197 | 0.018 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, L.; Zhang, Y.; Cui, Z.; Cao, C. Responses of Soil Microbial Communities Associated with Phosphorus Transformation to Land-Use Alternations in a Meadow Grassland, Northeast China. Microorganisms 2025, 13, 624. https://doi.org/10.3390/microorganisms13030624
Yu L, Zhang Y, Cui Z, Cao C. Responses of Soil Microbial Communities Associated with Phosphorus Transformation to Land-Use Alternations in a Meadow Grassland, Northeast China. Microorganisms. 2025; 13(3):624. https://doi.org/10.3390/microorganisms13030624
Chicago/Turabian StyleYu, Li, Ying Zhang, Zhenbo Cui, and Chengyou Cao. 2025. "Responses of Soil Microbial Communities Associated with Phosphorus Transformation to Land-Use Alternations in a Meadow Grassland, Northeast China" Microorganisms 13, no. 3: 624. https://doi.org/10.3390/microorganisms13030624
APA StyleYu, L., Zhang, Y., Cui, Z., & Cao, C. (2025). Responses of Soil Microbial Communities Associated with Phosphorus Transformation to Land-Use Alternations in a Meadow Grassland, Northeast China. Microorganisms, 13(3), 624. https://doi.org/10.3390/microorganisms13030624