
The Detection of Extensively Drug-Resistant Proteus mirabilis Strains Harboring Both VIM-4 and VIM-75 Metallo-β-Lactamases from Patients in Germany

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Bacterial Isolates
2.2. Antibiotic Susceptibility Testing
2.3. Whole-Genome Sequencing and Genome Analyses
2.4. Ethical Approval
3. Results
3.1. Antibiotic Susceptibility
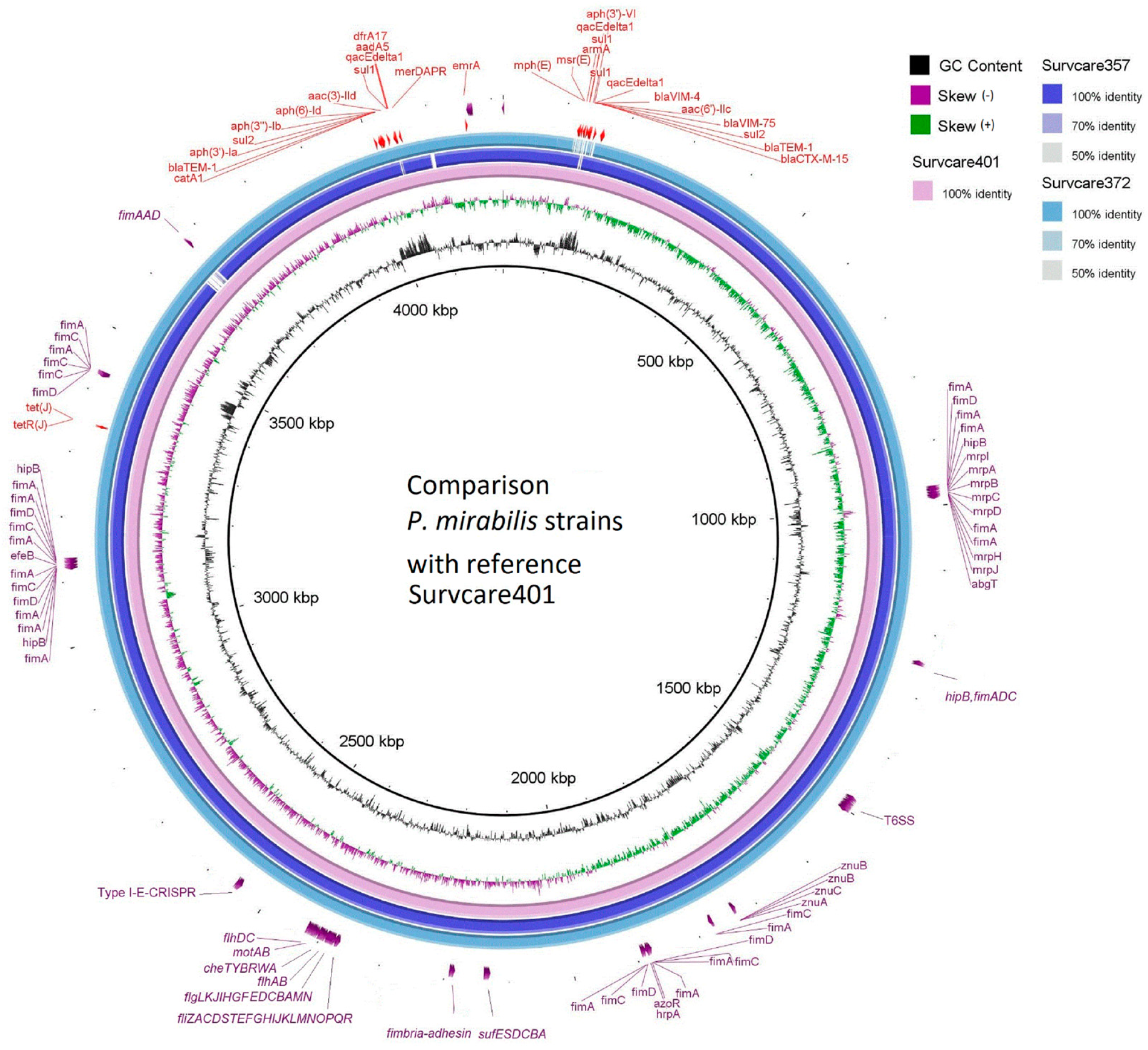
3.2. Whole-Genome Sequencing and Genomic Features
3.3. Antimicrobial Resistance Genes and the Co-Occurrences of Both blaVIM-75 and blaVIM-4
3.4. Plasmids
3.5. Virulence Factor Genes
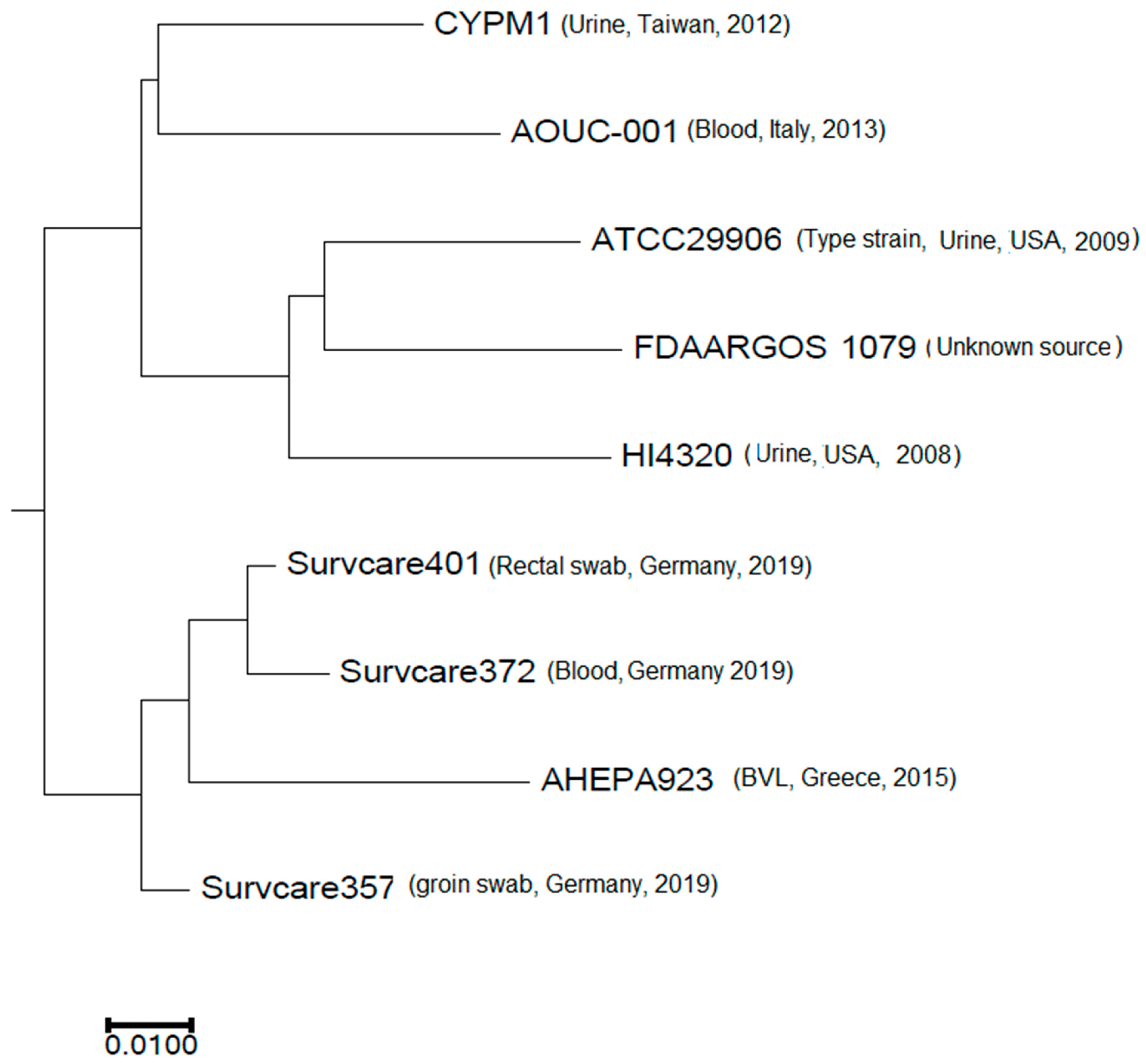
3.6. Phylogeny
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Armbruster, C.E.; Mobley, H.L.T.; Pearson, M.M. Pathogenesis of Proteus mirabilis Infection. EcoSal Plus 2018, 8, 1. [Google Scholar] [CrossRef]
- Giammanco, G.M.; Grimont, P.A.D.; Grimont, F.; Lefevre, M.; Giammanco, G.; Pignato, S. Phylogenetic analysis of the genera Proteus, Morganella and Providencia by comparison of rpoB gene sequences of type and clinical strains suggests the reclassification of Proteus myxofaciens in a new genus, Cosenzaea gen. nov., as Cosenzaea myxofaciens comb. nov. Int. J. Syst. Evol. Microbiol. 2011, 61, 1638–1644. [Google Scholar]
- Pearson, M.M.; Sebaihia, M.; Churcher, C.; Quail, M.A.; Seshasayee, A.S.; Luscombe, N.M.; Abdellah, Z.; Arrosmith, C.; Atkin, B.; Chillingworth, T.; et al. Complete genome sequence of uropathogenic Proteus mirabilis, a master of both adherence and motility. J. Bacteriol. 2008, 190, 4027–4037. [Google Scholar] [CrossRef]
- Schaffer, J.N.; Pearson, M.M. Proteus mirabilis and Urinary Tract Infections. In Urinary Tract Infections: Molecular Pathogenesis and Clinical Management; Wiley: Hoboken, NJ, USA, 2015; Volume 3. [Google Scholar]
- Stock, I. Natural antibiotic susceptibility of Proteus spp., with special reference to P. mirabilis and P. penneri strains. J. Chemother. 2003, 15, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Girlich, D.; Bonnin, R.A.; Dortet, L.; Naas, T. Genetics of Acquired Antibiotic Resistance Genes in Proteus spp. Front. Microbiol. 2020, 11, 256. [Google Scholar] [CrossRef] [PubMed]
- Vourli, S.; Tsorlini, H.; Katsifa, H.; Polemis, M.; Tzouvelekis, L.S.; Kontodimou, A.; Vatopoulos, A.C. Emergence of Proteus mirabilis carrying the blaVIM-1 metallo-β-lactamase gene. Clin. Microbiol. Infect. 2006, 12, 691–694. [Google Scholar] [CrossRef] [PubMed]
- Markovska, R.; Schneider, I.; Keuleyan, E.; Ivanova, D.; Lesseva, M.; Stoeva, T.; Sredkova, M.; Bauernfeind, A.; Mitov, I. Dissemination of a multidrug-resistant VIM-1- and CMY-99-producing Proteus mirabilis clone in Bulgaria. Microb. Drug Resist. 2017, 23, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Robert Koch-Institut. Bericht des Nationalen Referenzzentrums für Gramnegative Krankenhauserreger; Robert Koch-Institut: Berlin, Germany, 2023. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Yao, Y.; Imirzalioglu, C.; Falgenhauer, L.; Falgenhauer, J.; Heinmüller, P.; Domann, E.; Chakraborty, T. Plasmid-Mediated Spread of Carbapenem Resistance in Enterobacterales: A Three-Year Genome-Based Survey. Antibiotics 2024, 13, 682. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Doijad, S.; Falgenhauer, J.; Schmiedel, J.; Imirzalioglu, C.; Chakraborty, T. Co-occurrence of dual carbapenemases KPC-2 and OXA-48 with the mobile colistin resistance gene mcr-9.1 in Enterobacter xiangfangensis. Front. Cell. Infect. Microbiol. 2022, 12, 960892. [Google Scholar] [CrossRef] [PubMed]
- Schwengers, O.; Hoek, A.; Fritzenwanker, M.; Falgenhauer, L.; Hain, T.; Chakraborty, T.; Goesmann, A. ASA3P: An automatic and scalable pipeline for the assembly, annotation and higher level analysis of closely related bacterial isolates. PLoS Comput. Biol. 2020, 16, e1007134. [Google Scholar] [CrossRef] [PubMed]
- Carattoli, A.; Zankari, E.; García-Fernández, A.; Voldby Larsen, M.; Lund, O.; Villa, L.; Møller Aarestrup, F.; Hasman, H. In silico detection and typing of plasmids using PlasmidFinder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef]
- Naas, T.; Oueslati, S.; Bonnin, R.A.; Dabos, M.L.; Zavala, A.; Dortet, L.; Retailleau, P.; Iorga, B.I. Beta-lactamase database (BLDB)–structure and function. J. Enzyme Inhib. Med. Chem. 2017, 32, 917–919. [Google Scholar] [CrossRef]
- Li, X.; Xie, Y.; Liu, M.; Tai, C.; Sun, J.; Deng, Z.; Ou, H.Y. OriTfinder: A web-based tool for the identification of origin of transfers in DNA sequences of bacterial mobile genetic elements. Nucleic Acids Res. 2018, 46, W229–W234. [Google Scholar] [CrossRef]
- Johansson, M.H.K.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Tsafnat, G. Automated annotation of mobile antibiotic resistance in Gram-negative bacteria: The Multiple Antibiotic Resistance Annotator (MARA) and database. J. Antimicrob. Chemother. 2018, 73, 883–890. [Google Scholar] [CrossRef]
- Couvin, D.; Bernheim, A.; Toffano-Nioche, C.; Touchon, M.; Michalik, J.; Néron, B.; Rocha, E.P.C.; Vergnaud, G.; Gautheret, D.; Pourcel, C. CRISPRCasFinder, an update of CRISRFinder, includes a portable version, enhanced performance and integrates search for Cas proteins. Nucleic Acids Res. 2018, 46, W246–W251. [Google Scholar] [CrossRef]
- Mojica, M.F.; Bonomo, R.A.; Fast, W. B1-Metallo-β-Lactamases: Where Do We Stand? Curr. Drug Targets 2015, 17, 1029–1050. [Google Scholar] [CrossRef]
- Fuchs, F.; Ahmadzada, A.; Plambeck, L.; Wille, T.; Hamprecht, A. Susceptibility of Clinical Enterobacterales Isolates With Common and Rare Carbapenemases to Mecillinam. Front. Microbiol. 2021, 11, 627267. [Google Scholar] [CrossRef] [PubMed]
- Heiden, S.E.; Sydow, K.; Schaefer, S.; Klempien, I.; Balau, V.; Bauer, P.; Hübner, N.O.; Schaufler, K. Nearly identical plasmids encoding vim-1 and mercury resistance in enterobacteriaceae from north-eastern germany. Microorganisms 2021, 9, 1345. [Google Scholar] [CrossRef] [PubMed]
- Campos-Madueno, E.I.; Sigrist, T.; Flückiger, U.M.; Risch, L.; Bodmer, T.; Endimiani, A. First report of a blaVIM-1 metallo-β-lactamase-possessing Klebsiella michiganensis. J. Glob. Antimicrob. Resist. 2021, 25, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Papagiannitsis, C.C.; Miriagou, V.; Kotsakis, S.D.; Tzelepi, E.; Vatopoulos, A.C.; Petinaki, E.; Tzouvelekis, L.S. Characterization of a transmissible plasmid encoding VEB-1 and VIM-1 in Proteus mirabilis. Antimicrob. Agents Chemother. 2012, 56, 4024–4025. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, K.; Voets, G.M.; Scharringa, J.; Voskuil, S.; Fluit, A.C.; Rottier, W.C.; Leverstein-Van Hall, M.A.; Cohen Stuart, J.W.T. A disc diffusion assay for detection of class A, B and OXA-48 carbapenemases in Enterobacteriaceae using phenyl boronic acid, dipicolinic acid and temocillin. Clin. Microbiol. Infect. 2014, 20, 345–349. [Google Scholar] [CrossRef]
- Protonotariou, E.; Poulou, A.; Politi, L.; Meletis, G.; Chatzopoulou, F.; Malousi, A.; Metallidis, S.; Tsakris, A.; Skoura, L. Clonal outbreak caused by VIM-4-producing Proteus mirabilis in a Greek tertiary-care hospital. Int. J. Antimicrob. Agents 2020, 56, 106060. [Google Scholar] [CrossRef] [PubMed]
- Aberkane, S.; Compain, F.; Barraud, O.; Ouédraogo, A.S.; Bouzinbi, N.; Vittecoq, M.; Jean-Pierre, H.; Decré, D.; Godreuil, S. Non-O1/non-O139 Vibrio cholerae avian isolate from France cocarrying the blaVIM-1 and blaVIM-4 genes. Antimicrob. Agents Chemother. 2015, 59, 6594–6596. [Google Scholar] [CrossRef] [PubMed]
- Bontron, S.; Poirel, L.; Kieffer, N.; Savov, E.; Trifonova, A.; Todorova, I.; Kueffer, G.; Nordmann, P. Increased resistance to carbapenems in Proteus mirabilis mediated by amplification of the blaVIM-1-carrying and IS26-associated class 1 integron. Microb. Drug Resist. 2019, 25, 663–667. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Lee, Y.S.; Park, Y.K.; Kim, B.S. Alterations in the GyrA and GyrB subunits of topoisomerase II and the ParC and ParE subunits of topoisomerase IV in ciprofloxacin-resistant clinical isolates of Pseudomonas aeruginosa. Int. J. Antimicrob. Agents 2005, 25, 290–295. [Google Scholar] [CrossRef]
- Varughese, L.R.; Rajpoot, M.; Goyal, S.; Mehra, R.; Chhokar, V.; Beniwal, V. Analytical profiling of mutations in quinolone resistance determining region of gyrA gene among UPEC. PLoS ONE 2018, 13, e0190729. [Google Scholar] [CrossRef] [PubMed]
- Abdelkreem, R.H.; Yousuf, A.M.; Elmekki, M.A.; Elhassan, M.M. Dna gyrase and topoisomerase iv mutations and their effect on quinolones resistant Proteus mirabilis among utis patients. Pak. J. Med. Sci. 2020, 36, 1234–1240. [Google Scholar] [CrossRef] [PubMed]
- Saito, R.; Sato, K.; Kumita, W.; Inami, N.; Nishiyama, H.; Okamura, N.; Moriya, K.; Koike, K. Mutations of DNA gyrase and topoisomerase IV in clinical isolates of fluoroquinolone-resistant Proteus mirabilis. Jpn. J. Antibiot. 2006, 59, 41–43. [Google Scholar] [PubMed]
- Bitar, I.; Marchetti, V.M.; Mercato, A.; Nucleo, E.; Anesi, A.; Bracco, S.; Rognoni, V.; Hrabak, J.; Migliavacca, R. Complete genome and plasmids sequences of a clinical Proteus mirabilis isolate producing plasmid mediated ndm-1 from Italy. Microorganisms 2020, 8, 339. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Isolates | Survcare357 | Survcare372 | Survcare401 |
|---|---|---|---|
| Isolate source | Groin swab | Blood | Rectal swab |
| Patient ages (years) | 82 | 80 | 91 |
| Patient sex | Female | Male | Male |
| Imipenem | ≥16 [R] (>32) | ≥16 [R] (12) | 8 [R] (6) |
| Meropenem | ≥16 [R] (8) | 4 [I] (0.1) | 1 [I] (0.125) |
| Ertapenem | ≥8 [R] | 4 [I] | ≤0.12 [S] |
| Tigecycline | R | R | ND |
| Trimethoprim + Sulfamethoxazol | ≥320 [R] | ≥320 [R] | ND |
| Ampicillin | ≥32 [R] | ≥32 [R] | ≥32 [R] |
| Ampicillin + Sulbactam | ≥32 [R] | ≥32 [R] | ≥32 [R] |
| Piperacillin | ≥128 [R] | ≥128 [R] | ≥128 [R] |
| Piperacillin + Tazobactam | ≥128 [R] | 8 [R] | 8 [R] |
| Cefepim | R | R | R |
| Cefpodoxim | R | R | R |
| Cefotaxime | ≥64 [R] | ≥64 [R] | 16 [R] |
| Ceftazidime | ≥64 [R] | ≥64 [R] | ≥64 [R] |
| Cefuroxim | ≥64 [R] | ≥64 [R] | ≥64 [R] |
| Aztreonam | R | R | R |
| Ciprofloxaxin | ≥4 [R] | ≥4 [R] | 0.5 [I] |
| Moxifloxacin | R | R | R |
| Ofloxacin | R | R | R |
| Gentamicin | R | ≥16 [R] | ≥16 [R] |
| Isolate | Genomic Features | Assembly Status | Size (bp) | ARGs * |
|---|---|---|---|---|
| Survcare401 | Chromosome | circular | 4,218,249 | β–lactams: blaVIM-75, blaVIM-4, blaCTX-M-15, blaTEM-1A, blaTEM-1B Aminoglycosides: aadA1, aadA5, armA, aac(6′)-IIc, aac(3)-IId, aph(3′)-Ia, aph(3′)-VI, strA, strB Amphenicol: cat, catA1 Sulphonamide: sul1 (3x), sul2 (2x) Tetracycline: tet(J) Macrolide/Streptogramin B: msr(E), mph(E) Trimethoprim: dfrA1, dfrA17 Quaternary ammonium compound: qacΔE (3x) |
| Plasmid p401-1 | circular | 40,917 | β–lactams: blaTEM-2 | |
| Plasmid p401-2 | circular | 101,867 | Aminoglycosides: aadA1, aph(3′)-Ic Lincosamide: lnu(F) | |
| Survcare357 | Chromosome | linear | ~4,191,000 | β–lactams: blaVIM-75, blaVIM-4, blaCTX-M-15, blaTEM-1A, blaTEM-1B Aminoglycosides: aadA1, armA, aac(6′)-IIc, aac(3)-IId, aph(3′)-Ia, aph(3′)-VI, strA, strB Amphenicol: cat, catA1, Sulphonamide: sul1, sul2 (2x) Tetracycline: tet(J) Macrolide/Streptogramin B: msr(E), mph(E) Trimethoprim: dfrA1 Quaternary ammonium compound: qacΔE (3x). |
| Plasmid p357-1 | circular | 40,920 | β–lactams: blaTEM-2 | |
| Survcare372 | Chromosome | linear | ~4,165,000 | β—lactams: blaCTX-M-15 (2x), blaTEM-1A, blaTEM-1B Aminoglycosides: aadA1, aadA5, aac(3)-IId, aph(3′)-Ia, strA, strB Amphenicol: cat, catA1 Sulphonamide: sul1 Tetracycline: tet(J) Trimethoprim: dfrA1, dfrA17 Quaternary ammonium compound: qacΔE. |
| Plasmid p372-1 | circular | 40,918 | β–lactams: blaTEM-2 | |
| Plasmid p372-2 | circular | 101,866 | Aminoglycosides: aadA1, aph(3′)-Ic Lincosamide: lnu(F) | |
| Plasmid p372-3 | circular | 46,639 | β—lactams: blaVIM-75, blaVIM-4 Aminoglycosides: armA, aac(6′)-IIc, aph(3′)-VIa Sulphonamide: sul1 (2x), sul2 Macrolide/Streptogramin B: msr(E), mph(E) Quaternary ammonium compound: qacΔE (2x). |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fritzenwanker, M.; Falgenhauer, J.; Hain, T.; Imirzalioglu, C.; Chakraborty, T.; Yao, Y. The Detection of Extensively Drug-Resistant Proteus mirabilis Strains Harboring Both VIM-4 and VIM-75 Metallo-β-Lactamases from Patients in Germany. Microorganisms 2025, 13, 266. https://doi.org/10.3390/microorganisms13020266
Fritzenwanker M, Falgenhauer J, Hain T, Imirzalioglu C, Chakraborty T, Yao Y. The Detection of Extensively Drug-Resistant Proteus mirabilis Strains Harboring Both VIM-4 and VIM-75 Metallo-β-Lactamases from Patients in Germany. Microorganisms. 2025; 13(2):266. https://doi.org/10.3390/microorganisms13020266
Chicago/Turabian StyleFritzenwanker, Moritz, Jane Falgenhauer, Torsten Hain, Can Imirzalioglu, Trinad Chakraborty, and Yancheng Yao. 2025. "The Detection of Extensively Drug-Resistant Proteus mirabilis Strains Harboring Both VIM-4 and VIM-75 Metallo-β-Lactamases from Patients in Germany" Microorganisms 13, no. 2: 266. https://doi.org/10.3390/microorganisms13020266
APA StyleFritzenwanker, M., Falgenhauer, J., Hain, T., Imirzalioglu, C., Chakraborty, T., & Yao, Y. (2025). The Detection of Extensively Drug-Resistant Proteus mirabilis Strains Harboring Both VIM-4 and VIM-75 Metallo-β-Lactamases from Patients in Germany. Microorganisms, 13(2), 266. https://doi.org/10.3390/microorganisms13020266